To eat? To sleep? To run? Coordination of innate behaviors by lateral hypothalamus
-
Tatiana Korotkova
Photo: Philipp Jester
Tatiana Korotkova studied biology with a focus on human and animal physiology in Lomonosov Moscow State University. During her Ph.D. in the lab of Prof.H.L. Haas in Düsseldorf she investigated actions of various hypothalamic neuropeptides on aminergic nuclei, particularly on dopaminergic system. As a postdoc in the lab of Prof H.Monyer in Heidelberg, and later in a collaboration with Prof. T. J. Jentsch in Berlin she studied mechanisms of hippocampal network oscillations in behaving transgenic mice. Since 2012 T.Korotkova is a junior group leader at the Leibniz Institute of Molecular Pharmacology (FMP)/Neurocure Cluster of Excellence in Berlin, since April 2017 – a Research group leader at the Max Planck Institute for Metabolism Research, Cologne, Germany. Her work focuses on functions of lateral hypothalamus in hunger and sleep. To unravel the roles of hypothalamic circuits in health and disease, she combines optogenetics with neuronal recordings in behaving mice. She was a holder of the DFG Graduate College 320 and Schering foundation research stipends, and was awarded the Junior Brain Prize by Lundbeck Foundation.
 and
Alexey Ponomarenko
and
Alexey Ponomarenko
Alexey Ponomarenko studied biology with a focus on physiology of humans and animals in Lomonosov Moscow State University. During his Ph.D. with Prof. H.L.Haas in Duesseldorf he studied fast network oscillations in hippocampus and amygdala using electrophysiological recordings in behaving rats. He then learned
in vivo stereotrode recordings in the laboratory of Prof. G. Buzsaki (Rutgers University, USA). During postdoctoral training with Prof. H. Monyer in Heidelberg and following joint studies with Prof. T.J. Jentsch in Berlin, he implemented neuronal recordings in genetic mouse models and studied network mechanisms of hippocampal oscillations and spatial representations. Since 2009 A. Ponomarenko leads a junior group at the FMP Institute/Neurocure Cluster of Excellence in Berlin. He uses optogenetics and high-density electrophysiology to manipulate synchronization between cortical and subcortical regions and to explore its function in innate behaviors. He is a holder of Arthur A. Hancock Young Investigator Award in the European Histamine Research Society.

Abstract
Lateral hypothalamus (LH) is crucial for the regulation of innate behaviors, including food intake, sleep, wakefulness and locomotion. Pathologies of LH are associated with widespread sleep and eating disorders. Recent progress in several technologies, including optogenetics, multisite high-density neuronal recordings and Ca2+ imaging in behaving mice have enabled cell-type specific readout of activity in hypothalamus, as well as identification and manipulation of defined neuronal populations and their projections with high temporal resolution. Here we summarize recent studies characterizing the functions of orexin/hypocretin-, GABA- and MCH (melanin-concentrating hormone)-expressing LH neurons in innate behaviors. Further, we describe how top-down inputs convey information from other forebrain regions to LH and regulate innate behaviors. We then elucidate how spontaneous or optogenetically induced changes of neuronal activity in hypothalamic circuits are translated into behavior.
Innate behaviors, their disorders and neural substrates
Coordination of innate behaviors is instrumental for survival: a sufficient level of arousal is necessary to perform food-seeking and consumption, while hyperactivity might prevent goal-directed behavior, necessary for successful food-seeking. On the other hand, the ability to stop feeding and start locomotion is crucial to escape threats, e.g. if a predator appears. Pathologies of innate behaviors lead to sleep and eating disorders, which are widespread in all age groups. Eating disorders are difficult to treat and extremely dangerous, displaying the highest mortality rate of all psychiatric disorders (Kaye et al., 2009). Remarkably, changes of innate behaviors are usually not isolated: patients suffering from narcolepsy or chronic sleep restriction have an increased risk of developing obesity; obesity patients have disrupted sleep patterns (Broussard and Van Cauter, 2016); the majority of patients suffering from anorexia nervosa exhibit excessive physical activity (Davis et al., 1997). Thus, understanding of neuronal regulation of innate behaviors also gains insights into mechanisms of eating and sleep disorders.
One of the key brain regions, instrumental for feeding, sleep and locomotion, is hypothalamus (Saper et al., 1997; Morton et al., 2014; Waterson and Horvath, 2015). An indication of functional specializations in hypothalamus stems from classical studies, showing that lesions or electrical stimulation of distinct parts of hypothalamus, designated as discrete anatomical “centers”, results in dramatic changes of drinking, feeding, sleep and other innate behaviors (Stellar, 1954; Sinnamon, 1993). The concept of hypothalamic “centers” has been, however, challenged by findings of their considerable redundancy, overlap and ambiguous anatomical localization, partially due to a relatively low anatomical specificity of manipulation techniques (reviewed by (Berridge, 2004)). Further, recent genetic studies revealed the presence of neurochemically distinct cell populations in hypothalamic areas, implicated in the control of feeding, sleep (Steculorum et al., 2015; Steiger et al., 2011; Bonnavion et al., 2016) and defensive (Lin et al., 2011) behaviors. Also in the lateral hypothalamus (LH), in addition to “fast” neurotransmitters (glutamate or GABA), cells co-express neuropeptides, including orexins/hypocretins or MCH (melanin-concentrating hormone) (reviewed in (Bonnavion et al., 2016; Schone and Burdakov, 2012)). Administration of these neuropeptides as well as studies in mutant mice showed that each of these neuronal groups plays a unique, complementary role in regulation of sleep, feeding and locomotion. For instance, orexins have been implicated in vigilance state control, feeding, exploratory activity and reward processing (reviewed in (Li and Lecea, 2014)). A causal involvement of these neuronal populations in behavior has been further revealed by ablation, opto- and chemogenetic manipulation and readouts of neuronal activity. Recent physiological and genetic manipulations of hypothalamic neuronal populations, including pro-opiomelanocortin (POMC), MCH, Agouti-related protein (AGRP) and orexins, vesicular GABA transporter-expressing (VGAT) cells, and their connections within and outside hypothalamus have led to further elaboration of the models of neural circuits, which enable feeding, foraging and vigilance state control (reviewed in (Bonnavion et al., 2016)).
Innate behaviors are conserved across species, their monitoring allows straightforward experimental designs in mice, where most molecular biological tools are available. Their application for brain-behavior analysis is further facilitated by links of specific behaviors to genetically distinct populations of hypothalamic neurons. At the same time, studies of innate behaviors pose a unique set of challenges, leading to new research horizons. Firstly, unlike other basal ganglia and cortical areas, hypothalamic connectivity has been only recently revealed and is still being elaborated using genetic anatomical tools. Secondly, data about intrinsic dynamics of hypothalamic networks in vivo are largely unavailable. Thirdly, since innate behaviors and their coordination are crucial for survival, their specific phases, i.e. initiation, execution and termination, eventually not only satisfy homeostatic needs, but are also tightly adjusted to a variety of environmental stimuli. For instance, initiation of feeding includes food approach, which can be more or less exploratory, depending on the familiarity of food locations in the environment. This involves prior associative learning and retrieval of information encoded by multiple forebrain regions involved in cognition. Similarly, termination of feeding by ventral striatal inputs to the lateral hypothalamus is a classic example of a top-down sentinel control, which terminates ingestion depending on environmental threats (O’Connor et al., 2015; Kelley et al., 2005). Embedding of innate patterns into complex adaptive behaviors involves function-specific readout of network dynamics, operating in other forebrain regions, by the lateral hypothalamus (Carus-Cadavieco et al., 2017).
Manipulation and monitoring of neural circuits
Recent progress in several technologies, including optogenetics, chemogenetics, Ca2+ imaging and multisite high-density neuronal recordings in behaving mice have enabled readout of activity of multiple cells in hypothalamus in behaving animals, as well as identification and manipulation of defined neuronal populations and their projections with high temporal resolution. The development and implementation of genetically encoded Ca2+ indicators, such as GCaMP, which emit fluorescent signal upon Ca2+ binding, allows optical monitoring of changes in intracellular Ca2+ concentrations, associated with fluctuations of neuronal activity. Implanting a microendoscopic lens (gradient-refractive-index (GRIN) lenses attached on either end of a relay lens) further enables simultaneous visualization of multiple single cells (Ziv et al., 2013) even in deep brain regions (Resendez et al., 2016). Electrophysiological recordings in behaving mice allow directly measuring of neuronal activity at high temporal resolution and collecting high yields of recorded cells using movable multisite electrodes which dimensions compare to a size of a neuron. Multitrodes (e.g. tetrodes and silicon probes) enable to better distinguish neurons in close proximity to each other, using a principle of triangulation (McNaughton et al., 1983; Buzsaki et al., 2015). The electrodes may be mounted on a microdrive, implanted and gradually advanced during several weeks of recordings to maximize number of neurons recorded.
The combination of electrophysiological recordings in behaving mice with optogenetics enables identification of cells of interest, since these cells respond to a laser pulse with a short latency, and may have their spontaneous activity monitored. Control of neuronal activity using excitatory or inhibitory opsins (light-sensitive channels or pumps) (Boyden et al., 2005; Wietek et al., 2014; Gunaydin et al., 2010; Zhang et al., 2007) further permits study of the causal role of a particular cell group. To target cell groups selectively, Cre-dependent opsins are introduced in a brain region of interest in Cre-recombinase driver mouse lines (Tye and Deisseroth, 2012). By combining monitoring of neuronal activity and rhythmic optogenetic stimulation of brain regions and cells, directly involved in the generation of network oscillations in particular brain circuits, mechanisms and causal role of synchronization across behaviors can be investigated (Buzsaki et al., 2015; Bender etz al., 2015; Yamamoto et al., 2014; Dupont et al., 2006; Draguhn et al., 1998; Wulff et al., 2009; Korotkova et al., 2010; Behrens et al., 2005; Korotkova and Ponomarenko, 2017). Readout of neuronal activity in a circuit in behaving animals prior to optogenetic manipulations allows establish a correlative link between specific behaviors and changes in neuronal activity. Subsequent optogenetic manipulation of the same circuit enables to study whether this relationship is causal. Finally, simultaneous readout of optogenetically induced changes of neuronal activity ensures that protocols chosen for optogenetic manipulations recapitulate physiological patterns of neuronal activity.
Broad capabilities of neural network manipulation and monitoring can be optimally exploited when experimental designs, preparations and data analysis are well tailored to questions in focus. The choice of an optical actuator depends on a time scale of neuronal dynamics and on an investigated functional output, which can be matched to photocurrent parameters of opsins, ranging from ultrafast variants (Wietek et al., 2014; Gunaydin et al., 2010; Berndt et al., 2014) to step-function opsins featuring steady activation (Yizhar et al., 2011). Opto- and chemogenetic inhibition are further examples of tools optimal for brief and temporally discrete vs. more prolonged behavioral read-outs, respectively, like food seeking vs. food intake (Stachniak et al., 2014). Anatomical and cell-type specificity of optogenetic stimulation is defined to a large extent by features of Cre driver mouse lines and/or viral promoters used to drive opsin expression, by viral serotypes and titers as well as by injection techniques which together influence volume of the transduced brain tissue. Reliable protocols have been established for major neuronal types and their efficacy has been demonstrated by many reports (reviewed in (Lerner et al., 2016; Grundemann and Luthi, 2015)). Mice which express the same fluorophor (e.g. YFP) in same brain regions, and receive the same light stimulation are used to control for effects of light exposure and local heating on behavior and neuronal activity. In the following parts we will summarize recent studies showing functions of LH neurons in innate behaviors.
Causal role of orexin-, MCH- and VGAT-expressing LH neurons in regulation of sleep and wakefulness
Each of these three non-overlapping (Jennings et al., 2015) LH populations has a unique role in regulation of sleep (Fig. 1). Administration of orexins/hypocretins activates aminergic arousal-related nuclei (Brown et al., 2002; Eriksson et al., 2001; Korotkova et al., 2003; Bourgin et al., 2000) and induces arousal (Saper et al., 2010), whereas disturbances of orexin system due to mutations causes narcolepsy both in humans and animals (Kilduff and Peyron, 2000). Electrophysiological recordings of various LH cells (GABA-, orexins-, and MCH-expressing) showed that each of these subgroups exhibit a characteristic state-dependent activity across sleep-wake cycle. Firing rate of orexin-expressing neurons is highest during active waking, and lowest during non-REM and REM sleep (Lee et al., 2005; Mileykovskiy et al., 2005). Optogenetic activation of orexin neurons increases the probability of wakening from non-REM sleep as well as from REM sleep (Adamantidis et al., 2007).
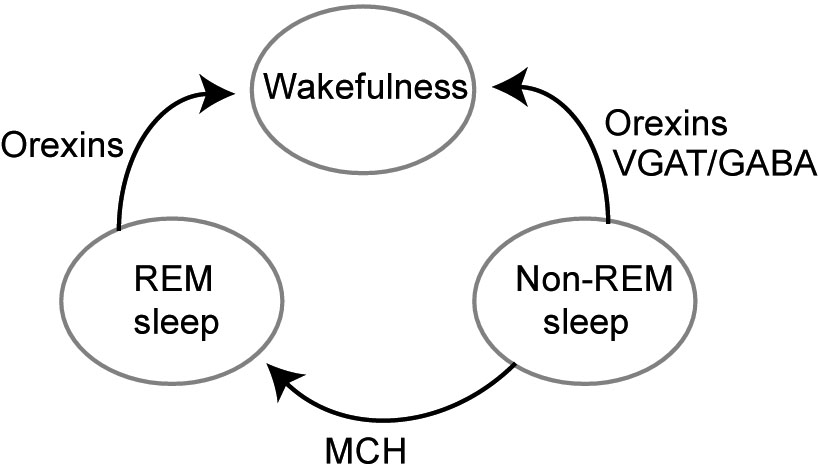
Scheme illustrating effects of wake-active orexin, wake transitions-active VGAT/GABA and sleep-active MCH cell groups on transitions between non-REM sleep, REM sleep, and wakefulness.
VGAT (vesicular GABA transporter)-expressing LH GABA (VGAT/GABA) cells also change their activity across sleep-wake cycle. We optogenetically identified these cells using opsin expression in VGAT-Cre mice and found that they show a transient increase of firing rate during transitions from non-REM sleep to wakefulness (Herrera et al., 2016). In accordance with this finding, optical stimulation during non-REM sleep induced a rapid transition to wakefulness, stronger stimulation led to faster transitions. The wakening upon VGAT/GABA LH cells activation occurred faster than upon activation of orexin-expressing LH cells (Adamantidis et al., 2007). The behavioral response was selective for non-REM sleep state, optogenetic activation of LH VGAT/GABA neurons during REM sleep did not lead to wakening, suggesting that LH VGAT/GABA cells are part of neuronal circuit selectively modulating transitions from non-REM. Non-REM sleep differs from REM sleep in a number of crucial features (see (Brown et al., 2012) for extensive review), and these stages of sleep are controlled by different neuronal circuits (Saper et al., 2010; Lee and Dan, 2012). In accordance with effects of selective optogenetic activation, a more prolonged, chemogenetic, activation of LH VGAT/GABA cells, using the excitatory DREADDs (designer receptors exclusively activated by a designer drug (DREADD), (Armbruster et al., 2007)), promoted wakefulness, whereas chemogenetic inhibition increased sleep (Venner et al., 2016).
LH VGAT/GABA cells project to multiple sleep-related regions, including locus coeruleus, dorsal thalamus (Herrera et al., 2016) and ventral lateral preoptic area (VLPO) (Venner et al., 2016). Further, LH VGAT/GABA neurons massively project to the reticular thalamic nucleus (RTN) – an inhibitory nucleus innervating multiple thalamic nuclei. Optogenetic stimulation of LH GABA terminals in the RTN during non-REM sleep induced a rapid transition to wakefulness, thus reproducing sleep wake transitions upon activation of LH VGAT/GABA cells bodies. To characterize changes of RTN activity upon sleep-wake transitions, we recorded their firing across sleep-wake cycle. We found that the majority of TRN cells displayed state-dependent activity: the variability of their firing was lower (i.e. cells fired more regularly) during spontaneous waking than during non-REM sleep. Optical stimulation of LH VGAT/GABA-TRN GABA projections replicated these state-dependent changes of firing variability in RTN cells (Herrera et al., 2016). Further, optogenetic inhibition of this pathway prolonged the duration of non-REM sleep, showing the necessity of LH GABA-RTN pathway for normal transitions to wakefulness.
Classical models suggested LH to be an “arousal” center while preoptic area since studies of von Economo and Nauta (1946) considered to be a “sleep center”, as its lesions produced insomnia. Accordingly, sleep-active neurons were identified both in inhibitory neurons in VLPO and the median preoptic area, reviewed in (Saper et al., 2010). However, recent studies showed that LH also contains sleep-active neurons. MCH-expressing cells exhibited increased activity during REM sleep compared to non-REM sleep and were least active during wakefulness (Jones and Hassani, 2013). MCH administration in rats caused hypersomnia (Verret et al., 2003), while the MCH-R1 antagonist promoted wakefulness (Ahnaou et al., 2008). In agreement, ablation of MCH neurons by cell-specific expression of diphtheria toxin A increased wakefulness and decreased non-REM sleep duration (Tsunematsu et al., 2014). Optogenetic activation of MCH neurons extended the duration of REM sleep episodes, induced transitions from non-REM to REM sleep (Jego et al., 2013) and increased REM sleep time while decreased non-REM sleep time (Tsunematsu et al., 2014). A 24-hours optogenetic stimulation of MCH cells hastened sleep onset, reduced length of wake episodes and increased total time in non-REM and REM sleep (Konadhode et al., 2013). Another possible sleep-promoting population in LH are LH GABA cells, expressing the long form of the leptin receptor b (LepRb). Selective optogenetic stimulation of these cells led to a significant increase of latency to wake (Herrera et al., 2016). Thus, orexins-, MCH- and VGAT/GABA-expressing LH cells selectively regulate particular phases of sleep: orexins and VGAT being wake promoting, and MCH and LH LepRB- sleep-promoting.
Multifaceted mechanisms of feeding behavior
Changes of VGAT/GABA-, orexin- and MCH-expressing LH neurons’ activity also influence feeding behavior. Optogenetic stimulation of LH VGAT/GABA neurons promotes voracious feeding while optogenetic inhibition of these cells reduces feeding even in food-deprived mice (Jennings et al., 2015). Similarly, selective activation of LH VGAT/GABA cells, projecting to the ventral tegmental area (VTA), a reward-related brain region, which is a major source of dopamine, leads to increase of sucrose-licking (Nieh et al., 2015), suggesting that reward system is involved in LH VGAT/GABA cells-mediated effects on feeding. Another pathway through which effects of LH VGAT/GABA cells may be mediated, are LH VGAT/GABA projections to hypothalamic paraventricular nucleus, since selective optogenetic stimulation of this pathway promotes feeding (Wu et al., 2015). Further, stimulation of LH VGAT/GABA at cells at different frequencies elicits differential effects on feeding and reward (Barbano et al., 2016), which might be explained by selective, frequency-dependent release of neuropeptides (Stuber and Wise, 2016), described also for other neurons (Schone and Burdakov, 2012). Interestingly, activation of LepRB-expressing LH cells, which also express GABA, shows an opposite effect to the activation of whole VGAT/GABA population, namely decreased feeding (Leinninger et al., 2009). This suggests that LH VGAT/GABA cell group actually comprises several populations with distinct functions depending on co-expressing peptides or differential connections.
The role of orexins in feeding seems to be more multidirectional. Their administration stimulates eating (Inutsuka et al, 2014), and orexin cells are activated during hunger (Diano et al., 2003). Activity of orexin cell population, measured by fiber photometry, decreased within milliseconds after eating onset, and remained decreased during eating, in hungry as well as in satiated mice (Gonzalez et al., 2016), suggesting that orexin neurons might signal hunger till food becomes available. At the same time, chronic inactivation of orexin cells by cell-specific expression of diphtheria toxin A induced increased feeding (Gonzalez et al., 2016). Since disturbances of orexin system lead to narcolepsy, which is often accompanied by overweight, it could be suggested that normal patterns and timing of sleep are important for feeding behavior, while typical for narcolepsy disturbed sleep timing, e.g. increased periods of arousal at times when mice eat most, lead to disturbed feeding behavior and obesity.
MCH system is also feeding-promoting, although its effects on feeding are more mild than actions of LH VGAT/GABA cells. MCH administration increases feeding, and levels of MCH mRNA increase during hunger (Qu et al., 1996). MCH knockout mice eat less and are more sensitive to starvation, showing greater weight loss and mortality, in contrast to littermates, even after a short fasting period (Shimada et al., 1998). Deletion of MCH receptor 1 disrupts overeating in the presence of food cues (Sherwood et al., 2015). Mice lacking MCH neurons show a reduced preference for sucrose over the artificial sweetener sucralose, compared to normal mice, and they release less dopamine than control mice during sucrose consumption. Finally, optogenetic stimulation of MCH neurons reverses natural preference for sucrose over sucralose (Domingos et al., 2013) suggesting that MCH neurons participate in sensing the nutritional value of sugar.
Feeding is also regulated by multiple extra-LH inputs (Fig. 2). First, feeding-promoting hypothalamic nuclei send projections onto LH. Optogenetic stimulation of projections of AGRP-expressing cells, located in arcuate nucleus (Betley et al., 2013), elicits feeding. Second, several extrahypothalamic inputs selectively regulate feeding (for review see e.g. (Bonnavion et al., 2016; Stuber and Wise, 2016). Bed nucleus of the stria terminalis, a region regulating various components of anxiety (Kim et al., 2013), sends inhibitory projections onto glutamatergic (VGlu2-expressing) LH cells, and its stimulation increases feeding (as does inhibition of VGlut cells) (Jennings et al., 2013). In contrast, a top-down input from NAcc shell neurons onto LH VGAT/GABA cells abruptly stop ongoing feeding (O’Connor et al., 2015; Kelley et al., 2005). Synchronization of LH inputs interacts with intrinsic LH activity as indicated by the role of the inhibitory input from the lateral septum (LS). Gamma oscillatory activity of LS projections, coordinated with gamma oscillations in LH, makes LH neurons with different feeding-related activity fire at distinct oscillation phases. The separate signaling by these LH populations is associated with changes of their firing rates, opposite to changes observed during feeding-inducing optostimulation of LH VGAT/GABA cells. The gamma-rhythmic input to LH from somatostatin-positive LS cells elicits food approach without affecting food intake (Carus-Cadavieco et al., 2017). Thus, multiple groups of LH neurons as well as inputs onto those cells selectively and complementary regulate various aspects of feeding behavior.
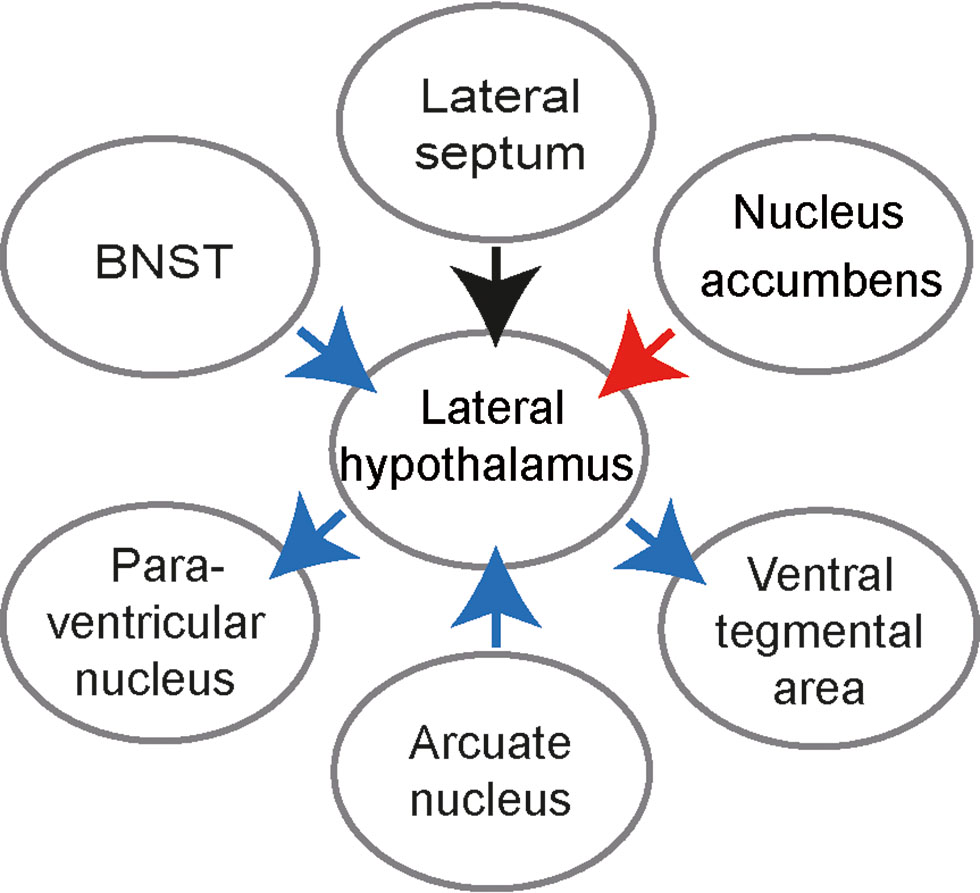
Scheme showing effects of LH inputs and outputs on feeding. Blue arrows: activation of a given pathway increases feeding, red arrow: activation of a pathway decreases feeding; black arrow: activation elicits food approach. BNST – Bed nucleus of the stria terminalis.
Oscillatory top-down signals and role of LH in locomotion
Eating disorders are often accompanied by changes in locomotor activity: about 80% of anorexia nervosa patients engaging in compulsive, excessive exercise (Davis et al., 1997). Rodents also display hyperactivity after food restriction, so-called activity-based anorexia (ABA): rats on a restricted diet profoundly increase activity in a running wheel (Routtenberg and Kuznesof, 1967; Adan et al., 2011). From classical electrical stimulation studies it is known that stimulation of LH induces locomotion, and can induce walking-like behavior even in anaesthetized rats (Sinnamon, 1993). We found that the majority of recorded LH neurons, including GABA cells, displayed locomotion-dependent firing: they increased discharge during initiation of locomotion and maintained increased firing during spontaneous running (Bender et al., 2015). As a part of the diencephalic locomotor region, LH is connected with midbrain and brain stem locomotor circuits and upstream forebrain regions (Sinnamon, 1993), which can provide LH with experience-dependent interpretation of sensory inputs.
A conceptual framework, helping to study regulation of innate behaviors, has been proposed by Swanson based on investigations of hypothalamic connections (Risold and Swanson, 1996). Behavioral control columns comprise dedicated behavioral systems of hypothalamus together with their cortical and basal ganglia input regions. Rostrally to premammillary nuclei, this system is involved in ingestive, reproductive, and defensive behaviors, while more caudal parts drive exploratory behavior. Upstream, an important interface between hypothalamus and cortical regions is the lateral septal nucleus (LS). LS is anatomically and functionally distinct from the medial septum, which projects to hippocampus and rhinal cortices in the medial temporal lobe and plays a key role in controlling timing of network activity in these areas. Medial septum has little if any connectivity with LS. The LS is the main source of forebrain inputs to hypothalamus, and the main subcortical output area of hippocampus (Fig. 3). In the rat, each pyramidal cell in the CA3 area and most of CA1 pyramidal cells project to LS, additionally to projections to the entorhinal cortex. There is a strict topographical mapping between hippocampus, LS and hypothalamus, i.e. hippocampal areas project to distinct portions of the LS and along the medio-lateral axis of hypothalamus (Risold and Swanson, 1996). These connections provide basis for behavioral control columns, routing hippocampal signals to hypothalamic regions, involved in feeding, locomotion, arousal and other innate behaviors. Our group has recently investigated information processing and some of the physiological functions mediated by cortico-hypothalamic circuits.
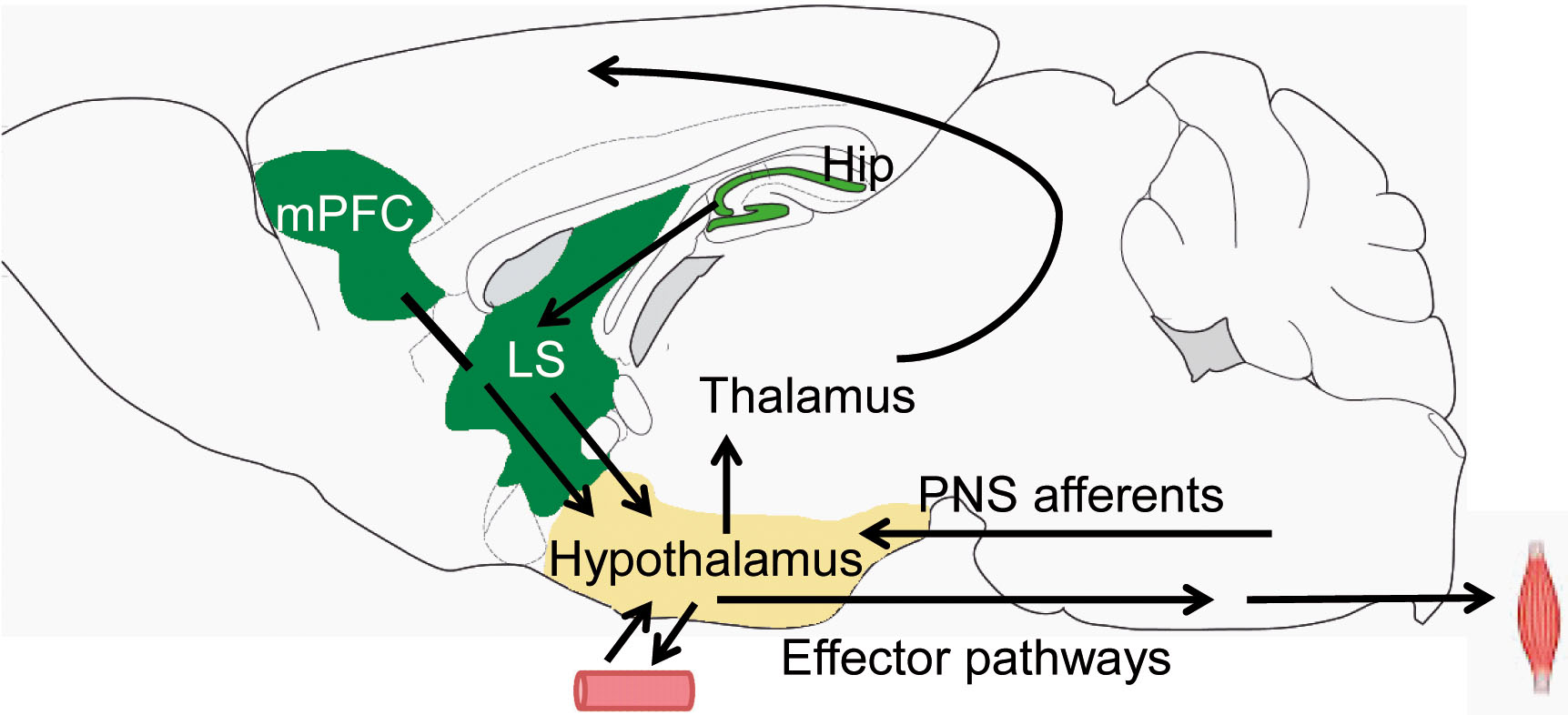
Circuit diagram showing inputs to hypothalamus from medial prefrontal cortex (mPFC) and hippocampus (Hip), routed via lateral septum (LS) as well as hypothalamic outputs to forebrain, brain stem and secretory effector pathways via hypophysis. PNS – peripheral nervous system. The mouse brain image was modified from (Paxinos und Franklin, 2001).
When animals or humans find themselves in a specific location in the environment, a place cell in the hippocampus becomes active. This brain navigation system signals each spatial position by activating relatively few place cells and, together with a grid cell network of the medial entorhinal cortex, is a fine-tuned dynamic machinery which follows its own rhythm. Rhythmic ups and downs in the activity of a large number of place cells during running occur 5 to 10 times per second, making activity of cells and electrical signals recorded from hippocampus oscillate with an impressive regularity of the so called theta oscillation. Since their discovery almost 80 years ago (Jung, 1938) theta oscillations have been implicated in communication of hippocampus with cortical and subcortical regions during spatial navigation and memory processing (reviewed by (Buzsaki and Moser, 2013; Seidenbecher et al., 2003; Stempel et al., 2016; Bikbaev and Manahan-Vaughan, 2008; Sirota et al., 2008)), motivational aspects of running, and in behavioral control of complex voluntary movements, including locomotion (Grastyan et al., 1965; Vanderwolf, 1969). Studies using electrophysiological monitoring, lesions or electrical stimulation indicated, however, a complex relationship between theta oscillations and locomotion.
Our laboratory used optogenetics to gain dynamic control over brain networks that pace theta oscillations in hippocampus. Using viral gene transfer and a light-sensitive protein, Channelrhodopsin-2, we targeted projections from the main pacemaker of the theta rhythm, medial septum, to the hippocampus. This way, brain synchronization could be selectively manipulated by an implanted optic fiber and microelectrode assembly in behaving mice with high temporal precision (Bender et al., 2015). The optogenetic entrainment regularized theta oscillations, i.e. rendered their frequency constant and the magnitude more regular while other aspects of information processing, including rhythmic discharge of pyramidal cells and place fields features, remained unchanged. Optogenetic control of theta oscillations released them from a multitude of influences normally generated by the interplay of brain state and sensory inputs and therefore opened a way to answering long-standing questions about the impact of hippocampal theta rhythmic activity on behavior (Grastyan et al., 1965; Vanderwolf, 1969). As the oscillation in hippocampus was optogenetically regularized, mice changed the way they moved through environment: they moved slower and with a more constant speed depending on the experimentally adjusted regularity of theta oscillations (Fig. 4 A,B).
How can hippocampal theta oscillations influence locomotion, initiation and pattern of which are controlled by hierarchically organized hypothalamic, midbrain and brain stem regions? One can view brain oscillations as traffic lights, which signal when neurons can fire and when they must stay silent; more regular oscillations resemble temporally coordinated traffic lights repeated with regular intervals. Hence, during regular theta rhythm activity of many traffic participants – populations of place cells – is more consistent over time. Does this activity affect outbound connections of hippocampus to the lateral septum and hypothalamus? We found that theta-periodic signals operate also downstream from hippocampus, in the LS: many cells in this brain region see and respond to “green” and “red” lights, switching on and off in synchrony with theta oscillations in the hippocampus. Thus swift transmission of neural signals along this route relies on theta oscillations, coordinated between hippocampus and LS.
Would mice move differently if the neural signal traffic in the hippocampus to LS pathway or downstream, from LS to lateral hypothalamus, is inhibited or stimulated? Inhibition of hippocampus to LS communication became possible applying “red light” protein switches (halorhodopsin, an inhibitory optogenetic tool) in the axons of hippocampal pyramidal cells in LS. The mouse’s speed did not react to manipulations with theta oscillations in the hippocampus when the outbound signalling from hippocampus to lateral septum was optogenetically blocked (Fig. 4 C,D). Further, many cells in the lateral hypothalamus become more active when an animal starts running, and theta rhythmic stimulation of inhibitory LS-LH projections decreased running speed (Fig. 4 E,F). Thus, theta oscillations provide a natural physiological signal, which operates in hippocampus – LS – lateral hypothalamus pathway and supports rapid adjustments of exploratory behavior in response to changes of hippocampal, cognition-related dynamics.
Interestingly, earlier studies demonstrated that theta rhythmic activation of hippocampus – LS – ventral tegmental area pathway is sufficient for context-dependent reinstatement of reward seeking (Luo et al., 2011). These findings highlight a prospective on theta oscillations, complementary to their role as a global clock signal, organizing encoding of spatial and, more generally, behavioral sequences. Indeed, theta synchronization operates at time scales potentially relevant for multiple brain functions. While an animal‘s path is signalled by place cells within single theta cycles, variations of theta synchronization at a slower scale (up to seconds) provide feedback from hippocampus to subcortical networks during locomotion and adjust its pattern. Since activity of place cells reflects animal experience, participation of pyramidal cells in synchronized ensembles can change depending on ongoing sensory input and thus determine spatiotemporal pattern of the oscillatory output (Bahner et al., 2011; Denker et al., 2011).
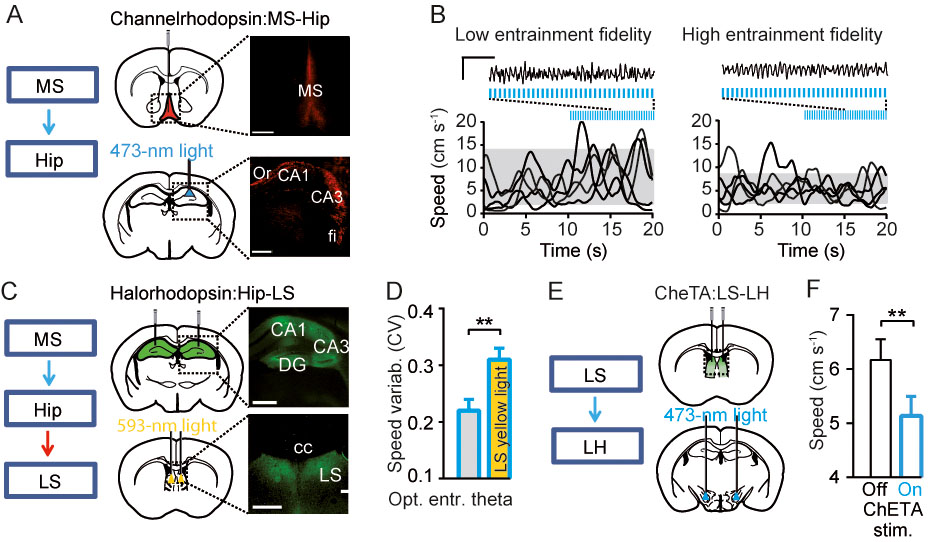
Regulation of locomotion via hippocampus-lateral septum-lateral hypothalamus pathway by theta oscillations. A. Optogenetic control of theta oscillations in behaving mice. Injections of Cre-dependent ChR2 in medial septum (MS) of parvalbumin-Cre mice and light-induced stimulation of MS-hippocampus (Hip) projections. Expression of ChR2.tdTomato: neuronal somata in MS and axons in Hip. Scale bars: 500 µm (left), 50 µm (right). B. Representative local field potential signals, recorded from hippocampus, before and after the onset of stimulation (7Hz) with a low (<0.3; left) or high (>0.8; right) theta oscillations entrainment fidelity, grey shadows mark 10-90% ranges of speed distributions during stimulation, blue lines mark optostimulation. Scale bars: 1 s, 2 mV. Note that efficiently controlled theta oscillations are accompanied by reduced speed variability. C. Left: Scheme of experimental design for simultaneous optogenetic entrainment of theta oscillations in Hip and inhibition of hippocampal outputs to LS. Right: Injections and expression of halorhodopsin (eNpHR3.0) in hippocampal pyramidal cells (top), axonal immunofluorescence in LS; bilateral optic fibers implantation for light-induced inhibition of Hip-LS projections (right). Scale bars: 500 µm (left), 50 µm (right). D. Reduction of speed variability during optogenetic theta entrainment was prevented by simultaneous LS yellow light (593 nm) delivery (yellow bar, p=0.007, N=8 mice). E. Targeting of ChETA, a faster version of channelrhodopsin (Gunaydin et al., 2010) to GABAergic cells in lateral septum (LS) mice and light-induced stimulation of LSGABA-LH projections. F. Optogenetic theta-frequency activation of LS-LH pathway decreases running speed (P=0.003, N=7 mice). Modified from (Bender et al., 2015).
Outlook
The results reviewed here suggest that innate behaviors provide a good model for functional investigation of neural circuits using a broad repertoire of genetic manipulation and physiological monitoring techniques in mice. Thanks to the integration of the core circuit for innate behaviors – hypothalamus – with sensory and associative cortices as well as basal ganglia, brain dynamics adaptively governing foraging, ingestion, vigilance state and other innate patterns can now be studied with high temporal resolution and anatomical specificity. These studies rely on quickly accumulating knowledge about connections between chemically and anatomically distinct classes of hypothalamic neurons as well as on anatomical mapping of their extrahypothalamic afferents and efferents. Further, investigation of intrinsic responses of hypothalamic cells to metabolic and hormonal signals together with recording and manipulation of their activity in vivo will provide detailed mechanistic information about hypothalamic integration. Future research will also address how hypothalamic networks read out information from upstream forebrain regions, providing new clues about a common language used by evolutionarily conserved subcortical and computationally advanced cortical networks (Gray and Singer, 1989; Yang et al., 2009; Herz et al., 2006). This will help understand organization of motivation behind exploratory stages of feeding, often executed in advance of nutritional deficits. Alterations of these processes highlight core symptoms of eating disorders and investigation of these circuits will lead to novel pathogenesis – oriented treatment strategies.
About the authors

Photo: Philipp Jester
Tatiana Korotkova studied biology with a focus on human and animal physiology in Lomonosov Moscow State University. During her Ph.D. in the lab of Prof.H.L. Haas in Düsseldorf she investigated actions of various hypothalamic neuropeptides on aminergic nuclei, particularly on dopaminergic system. As a postdoc in the lab of Prof H.Monyer in Heidelberg, and later in a collaboration with Prof. T. J. Jentsch in Berlin she studied mechanisms of hippocampal network oscillations in behaving transgenic mice. Since 2012 T.Korotkova is a junior group leader at the Leibniz Institute of Molecular Pharmacology (FMP)/Neurocure Cluster of Excellence in Berlin, since April 2017 – a Research group leader at the Max Planck Institute for Metabolism Research, Cologne, Germany. Her work focuses on functions of lateral hypothalamus in hunger and sleep. To unravel the roles of hypothalamic circuits in health and disease, she combines optogenetics with neuronal recordings in behaving mice. She was a holder of the DFG Graduate College 320 and Schering foundation research stipends, and was awarded the Junior Brain Prize by Lundbeck Foundation.

Alexey Ponomarenko studied biology with a focus on physiology of humans and animals in Lomonosov Moscow State University. During his Ph.D. with Prof. H.L.Haas in Duesseldorf he studied fast network oscillations in hippocampus and amygdala using electrophysiological recordings in behaving rats. He then learned in vivo stereotrode recordings in the laboratory of Prof. G. Buzsaki (Rutgers University, USA). During postdoctoral training with Prof. H. Monyer in Heidelberg and following joint studies with Prof. T.J. Jentsch in Berlin, he implemented neuronal recordings in genetic mouse models and studied network mechanisms of hippocampal oscillations and spatial representations. Since 2009 A. Ponomarenko leads a junior group at the FMP Institute/Neurocure Cluster of Excellence in Berlin. He uses optogenetics and high-density electrophysiology to manipulate synchronization between cortical and subcortical regions and to explore its function in innate behaviors. He is a holder of Arthur A. Hancock Young Investigator Award in the European Histamine Research Society.
Acknowledgements:
We thank Franziska Bender for her comments on the manuscript and for its translating into German language, and Constance Holman for her comments on the manuscript. This work was supported by the Deutsche Forschungsgemeinschaft (DFG; SPP1665, AP; Exc 257 NeuroCure, TK and AP), The German-Israeli Foundation for Scientific Research and Development (GIF; I-1326-421.13/2015, TK) and The Human Frontier Science Program (HFSP; RGY0076/2012, TK).
References
Adamantidis, A.R., et al. (2007). Neural substrates of awakening probed with optogenetic control of hypocretin neurons. Nature. 450(7168): p. 420–4.10.1038/nature06310Search in Google Scholar PubMed PubMed Central
Adan, R.A., et al. (2011). Neurobiology driving hyperactivity in activity-based anorexia. Curr Top Behav Neurosci. 6: p. 229–50.10.1007/7854_2010_77Search in Google Scholar PubMed
Ahnaou, A., et al. (2008). Blocking melanin-concentrating hormone MCH1 receptor affects rat sleep-wake architecture. Eur J Pharmacol. 579(1–3): p. 177–88.10.1016/j.ejphar.2007.10.017Search in Google Scholar PubMed
Armbruster, B.N., et al. (2007). Evolving the lock to fit the key to create a family of G protein-coupled receptors potently activated by an inert ligand. Proc. Natl. Acad. Sci. U.S.A. 104(12): p. 5163–8.10.1073/pnas.0700293104Search in Google Scholar PubMed PubMed Central
Bahner, F., et al. (2011). Cellular correlate of assembly formation in oscillating hippocampal networks in vitro. Proc. Natl. Acad. Sci. U.S.A. 108(35): p. E607–16.10.1073/pnas.1103546108Search in Google Scholar PubMed PubMed Central
Barbano, M.F., et al. (2016). Feeding and Reward Are Differentially Induced by Activating GABAergic Lateral Hypothalamic Projections to VTA. J Neurosci. 36(10): p. 2975–85.10.1523/JNEUROSCI.3799-15.2016Search in Google Scholar PubMed PubMed Central
Behrens, C.J., et al. (2005). Induction of sharp wave-ripple complexes in vitro and reorganization of hippocampal networks. Nat Neurosci. 8(11): p. 1560–7.10.1038/nn1571Search in Google Scholar PubMed
Bender, F., et al. (2015). Theta oscillations regulate the speed of locomotion via a hippocampus to lateral septum pathway. Nat Commun. 6: p. 8521.10.1038/ncomms9521Search in Google Scholar PubMed PubMed Central
Berndt, A., et al. (2014). Structure-guided transformation of channelrhodopsin into a light-activated chloride channel. Science. 344(6182): p. 420–4.10.1126/science.1252367Search in Google Scholar PubMed PubMed Central
Berridge, K.C. (2004). Motivation concepts in behavioral neuroscience. Physiol Behav. 81(2): p. 179–209.10.1016/j.physbeh.2004.02.004Search in Google Scholar PubMed
Betley, J.N., et al. (2013). Parallel, redundant circuit organization for homeostatic control of feeding behavior. Cell. 155(6): p. 1337–50.10.1016/j.cell.2013.11.002Search in Google Scholar PubMed PubMed Central
Bikbaev, A. and D. Manahan-Vaughan (2008). Relationship of hippocampal theta and gamma oscillations to potentiation of synaptic transmission. Front Neurosci. 2(1): p. 56–63.10.3389/neuro.01.010.2008Search in Google Scholar PubMed PubMed Central
Bonnavion, P., et al. (2016). Hubs and spokes of the lateral hypothalamus: cell types, circuits and behaviour. J Physiol. 594(22): p. 6443–6462.10.1113/JP271946Search in Google Scholar PubMed PubMed Central
Bourgin, P., et al. (2000). Hypocretin-1 modulates rapid eye movement sleep through activation of locus coeruleus neurons. J Neurosci. 20(20): p. 7760–5.10.1523/JNEUROSCI.20-20-07760.2000Search in Google Scholar
Boyden, E.S., et al. (2005). Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci. 8(9): p. 1263–8.10.1038/nn1525Search in Google Scholar PubMed
Broussard, J.L. and E. Van Cauter (2016). Disturbances of sleep and circadian rhythms: novel risk factors for obesity. Curr Opin Endocrinol Diabetes Obes. 23(5): p. 353–9.10.1097/MED.0000000000000276Search in Google Scholar PubMed PubMed Central
Brown, R.E., et al. (2002). Convergent excitation of dorsal raphe serotonin neurons by multiple arousal systems (orexin/hypocretin, histamine and noradrenaline). J Neurosci. 22(20): p. 8850–9.10.1523/JNEUROSCI.22-20-08850.2002Search in Google Scholar
Brown, R.E., et al. (2012). Control of sleep and wakefulness. Physiol Rev. 92(3): p. 1087–187.10.1152/physrev.00032.2011Search in Google Scholar PubMed PubMed Central
Buzsaki, G. and E.I. Moser (2013). Memory, navigation and theta rhythm in the hippocampal-entorhinal system. Nat Neurosci. 16(2): p. 130–8.10.1038/nn.3304Search in Google Scholar PubMed PubMed Central
Buzsaki, G., et al. (2015). Tools for probing local circuits: high-density silicon probes combined with optogenetics. Neuron. 86(1): p. 92–105.10.1016/j.neuron.2015.01.028Search in Google Scholar PubMed PubMed Central
Carus-Cadavieco, M., Gorbati, M., Ye, L., Bender, F., van der Veldt, S., Kosse, C., Börgers, C., Lee, S.Y., Ramakrishnan, C., Hu, Y., Denisova, N., Ramm, F., Volitaki, E., Burdakov, D., Deisseroth, K., Ponomarenko, A., Korotkova, T (2017). Gamma oscillations organize top-down signalling to hypothalamus and enable food seeking. Nature 542 (7640): p. 232–236. 10.1038/nature21066Search in Google Scholar
Davis, C., et al. (1997). The prevalence of high-level exercise in the eating disorders: etiological implications. Compr Psychiatry. 38(6): p. 321–6.10.1016/S0010-440X(97)90927-5Search in Google Scholar
Denker, M., et al. (2011). The local field potential reflects surplus spike synchrony. Cereb Cortex. 21(12): p. 2681–95.10.1093/cercor/bhr040Search in Google Scholar PubMed PubMed Central
Diano, S., et al. (2003). Fasting activates the nonhuman primate hypocretin (orexin) system and its postsynaptic targets. Endocrinology. 144(9): p. 3774–8.10.1210/en.2003-0274Search in Google Scholar PubMed
Domingos, A.I., et al. (2013). Hypothalamic melanin concentrating hormone neurons communicate the nutrient value of sugar. Elife. 2: p. e01462. 10.7554/eLife.01462Search in Google Scholar PubMed PubMed Central
Draguhn, A., et al. (1998). Electrical coupling underlies high-frequency oscillations in the hippocampus in vitro. Nature. 394(6689): p. 189–92.10.1038/28184Search in Google Scholar PubMed
Dupont, E., et al. (2006). Rapid developmental switch in the mechanisms driving early cortical columnar networks. Nature. 439(7072): p. 79–83.10.1038/nature04264Search in Google Scholar PubMed
Eriksson, K.S., et al. (2001). Orexin/hypocretin excites the histaminergic neurons of the tuberomammillary nucleus. J Neurosci. 21(23): p. 9273–9.10.1523/JNEUROSCI.21-23-09273.2001Search in Google Scholar
Gonzalez, J.A., et al. (2016). Inhibitory Interplay between Orexin Neurons and Eating. Curr Biol. 26(18): p. 2486–9110.1016/j.cub.2016.07.013Search in Google Scholar PubMed PubMed Central
Grastyan, E., et al. (1965). Hypothalamic Motivational Processes as Reflected by their Hippocampal Electrical Correlates. Science. 149(3679): p. 91–3.10.1126/science.149.3679.91Search in Google Scholar PubMed
Gray, C.M. and W. Singer (1989). Stimulus-specific neuronal oscillations in orientation columns of cat visual cortex. Proc. Natl. Acad. Sci. U.S.A. 86(5): p. 1698–702.10.1073/pnas.86.5.1698Search in Google Scholar PubMed PubMed Central
Grundemann, J. and A. Luthi (2015). Ensemble coding in amygdala circuits for associative learning. Curr Opin Neurobiol. 35: p. 200–6.10.1016/j.conb.2015.10.005Search in Google Scholar PubMed
Gunaydin, L.A., et al., Ultrafast optogenetic control. Nat Neurosci, (2010). 13(3): p. 387–92.10.1038/nn.2495Search in Google Scholar PubMed
Herrera, C.G., et al. (2016). Hypothalamic feedforward inhibition of thalamocortical network controls arousal and consciousness. Nat Neurosci. 19(2): p. 290–8.10.1038/nn.4209Search in Google Scholar PubMed PubMed Central
Herz, A.V., et al. (2006). Modeling single-neuron dynamics and computations: a balance of detail and abstraction. Science. 314(5796): p. 80–5.10.1126/science.1127240Search in Google Scholar PubMed
Inutsuka, A., et al. (2014). Concurrent and robust regulation of feeding behaviors and metabolism by orexin neurons. Neuropharmacology. 85: p. 451–60.10.1016/j.neuropharm.2014.06.015Search in Google Scholar PubMed
Jego, S., et al. (2013). Optogenetic identification of a rapid eye movement sleep modulatory circuit in the hypothalamus. Nat Neurosci. 16(11): p. 1637–43.10.1038/nn.3522Search in Google Scholar PubMed PubMed Central
Jennings, J.H., et al. (2013). The inhibitory circuit architecture of the lateral hypothalamus orchestrates feeding. Science. 341(6153): p. 1517–21.10.1126/science.1241812Search in Google Scholar PubMed PubMed Central
Jennings, J.H., et al. (2015). Visualizing hypothalamic network dynamics for appetitive and consummatory behaviors. Cell. 160(3): p. 516–27.10.1016/j.cell.2014.12.026Search in Google Scholar PubMed PubMed Central
Jones, B.E. and O.K. Hassani (2013). The role of Hcrt/Orx and MCH neurons in sleep-wake state regulation. Sleep. 36(12): p. 1769–72.10.5665/sleep.3188Search in Google Scholar PubMed PubMed Central
Jung R., K.A. E. (1938). Eine Methodik der Ableitung Iokalisierter Potentialschwankungen aus subcorticalen Hirngebieten. European Archives of Psychiatry and Clinical Neuroscience. 109(1): p. 1–30.10.1007/BF02157817Search in Google Scholar
Kaye, W.H., J.L. Fudge, and M. Paulus (2009). New insights into symptoms and neurocircuit function of anorexia nervosa. Nat Rev Neurosci. 10(8): p. 573–84.10.1038/nrn2682Search in Google Scholar
Kelley, A. E., et al. (2005). Corticostriatal-hypothalamic circuitry and food motivation: integration of energy, action and reward. Physiol Behav. 86(5): p. 773–95.10.1016/j.physbeh.2005.08.066Search in Google Scholar
Kilduff, T.S. and C. Peyron (2000). The hypocretin/orexin ligand-receptor system: implications for sleep and sleep disorders. Trends Neurosci. 23(8): p. 359–65.10.1016/S0166-2236(00)01594-0Search in Google Scholar
Kim, S.Y., et al. (2013). Diverging neural pathways assemble a behavioural state from separable features in anxiety. Nature. 496(7444): p. 219–23.10.1038/nature12018Search in Google Scholar PubMed PubMed Central
Konadhode, R.R., et al. (2013). Optogenetic stimulation of MCH neurons increases sleep. J Neurosci. 33(25): p. 10257–63.10.1523/JNEUROSCI.1225-13.2013Search in Google Scholar PubMed PubMed Central
Korotkova, T., et al. (2010). NMDA receptor ablation on parvalbumin-positive interneurons impairs hippocampal synchrony, spatial representations, and working memory. Neuron. 68(3): p. 557–69.10.1016/j.neuron.2010.09.017Search in Google Scholar PubMed
Korotkova, T.M., et al. (2003). Excitation of ventral tegmental area dopaminergic and nondopaminergic neurons by orexins/hypocretins. J Neurosci. 23(1): p. 7–11.10.1523/JNEUROSCI.23-01-00007.2003Search in Google Scholar
Korotkova, T., Ponomarenko A. (2017). In In Vivo Neuropharmacology and Neurophysiology, P. A., Editor. Springer: New York. p. 67–88.10.1007/978-1-4939-6490-1_4Search in Google Scholar
Lee, M.G., O.K. Hassani, and B.E. Jones (2005). Discharge of identified orexin/hypocretin neurons across the sleep-waking cycle. J Neurosci. 25(28): p. 6716–20.10.1523/JNEUROSCI.1887-05.2005Search in Google Scholar PubMed PubMed Central
Lee, S.H. and Y. Dan (2012). Neuromodulation of brain states. Neuron. 76(1): p. 209–22.10.1016/j.neuron.2012.09.012Search in Google Scholar PubMed PubMed Central
Leinninger, G.M., et al. (2009). Leptin acts via leptin receptor-expressing lateral hypothalamic neurons to modulate the mesolimbic dopamine system and suppress feeding. Cell Metab. 10(2): p. 89–98.10.1016/j.cmet.2009.06.011Search in Google Scholar
Lerner, T.N., L. Ye, and K. Deisseroth (2016). Communication in Neural Circuits: Tools, Opportunities, and Challenges. Cell. 164(6): p. 1136–50.10.1016/j.cell.2016.02.027Search in Google Scholar
Li, J., Z. Hu, and L. de Lecea (2014). The hypocretins/orexins: integrators of multiple physiological functions. Br J Pharmacol. 171(2): p. 332–50.10.1111/bph.12415Search in Google Scholar
Lin, D., et al. (2011). Functional identification of an aggression locus in the mouse hypothalamus. Nature. 470(7333): p. 221–6.10.1038/nature09736Search in Google Scholar
Luo, A.H., et al. (2011). Linking context with reward: a functional circuit from hippocampal CA3 to ventral tegmental area. Science. 333(6040): p. 353–7.10.1126/science.1204622Search in Google Scholar
McNaughton, B.L., J. O’Keefe, and C.A. Barnes (1983). The stereotrode: a new technique for simultaneous isolation of several single units in the central nervous system from multiple unit records. J Neurosci Methods. 8(4): p. 391–7.10.1016/0165-0270(83)90097-3Search in Google Scholar
Mileykovskiy, B.Y., L.I. Kiyashchenko, and J.M. Siegel (2005). Behavioral correlates of activity in identified hypocretin/orexin neurons. Neuron. 46(5): p. 787–98.10.1016/j.neuron.2005.04.035Search in Google Scholar PubMed PubMed Central
Morton, G.J., T.H. Meek, and M.W. Schwartz (2014). Neurobiology of food intake in health and disease. Nat Rev Neurosci. 15(6): p. 367–78.10.1038/nrn3745Search in Google Scholar PubMed PubMed Central
Nieh, E.H., et al. (2015). Decoding neural circuits that control compulsive sucrose seeking. Cell. 160(3): p. 528–41.10.1016/j.cell.2015.01.003Search in Google Scholar PubMed PubMed Central
O’Connor, E.C., et al. (2015). Accumbal D1R Neurons Projecting to Lateral Hypothalamus Authorize Feeding. Neuron. 88(3): p. 553–64.10.1016/j.neuron.2015.09.038Search in Google Scholar PubMed
Paxinos, G. und Franklin, K. B. J. The Mouse Brain in Stereotaxic Coordinates (Elsevier Academic Press, 2001).Search in Google Scholar
Qu, D., et al. (1996). A role for melanin-concentrating hormone in the central regulation of feeding behaviour. Nature. 380(6571): p. 243–7.10.1038/380243a0Search in Google Scholar
Resendez, S.L., et al. (2016). Visualization of cortical, subcortical and deep brain neural circuit dynamics during naturalistic mammalian behavior with head-mounted microscopes and chronically implanted lenses. Nat Protoc. 11(3): p. 566–97.10.1038/nprot.2016.021Search in Google Scholar
Risold, P.Y. and L.W. Swanson (1996). Structural evidence for functional domains in the rat hippocampus. Science. 272(5267): p. 1484–6.10.1126/science.272.5267.1484Search in Google Scholar
Routtenberg, A. and A.W. Kuznesof (1967). Self-starvation of rats living in activity wheels on a restricted feeding schedule. J Comp Physiol Psychol. 64(3): p. 414–21.10.1037/h0025205Search in Google Scholar
Saper, C.B., et al. (2010). Sleep state switching. Neuron. 68(6): p. 1023–42.10.1016/j.neuron.2010.11.032Search in Google Scholar
Schone, C. and D. Burdakov (2012). Glutamate and GABA as rapid effectors of hypothalamic “peptidergic” neurons. Front Behav Neurosci. 6: p. 81.10.3389/fnbeh.2012.00081Search in Google Scholar
Seidenbecher, T., et al. (2003). Amygdalar and hippocampal theta rhythm synchronization during fear memory retrieval. Science. 301(5634): p. 846–50.10.1126/science.1085818Search in Google Scholar
Sherwood, A., et al. (2015). Deletion of Melanin Concentrating Hormone Receptor-1 disrupts overeating in the presence of food cues. Physiol Behav. 152(Pt B): p. 402–7.10.1016/j.physbeh.2015.05.037Search in Google Scholar
Shimada, M., et al. (1998). Mice lacking melanin-concentrating hormone are hypophagic and lean. Nature. 396(6712): p. 670–4.10.1038/25341Search in Google Scholar
Sinnamon, H.M. (1993). Preoptic and hypothalamic neurons and the initiation of locomotion in the anesthetized rat. Prog Neurobiol. 41(3): p. 323–44.10.1016/0301-0082(93)90003-BSearch in Google Scholar
Sirota, A., et al. (2008). Entrainment of neocortical neurons and gamma oscillations by the hippocampal theta rhythm. Neuron. 60(4): p. 683–97.10.1016/j.neuron.2008.09.014Search in Google Scholar
Stachniak, T.J., A. Ghosh, and S.M. Sternson (2014). Chemogenetic synaptic silencing of neural circuits localizes a hypothalamus ➙ midbrain pathway for feeding behavior. Neuron. 82(4): p. 797–808.10.1016/j.neuron.2014.04.008Search in Google Scholar
Steculorum, S.M., et al. (2015). Hypothalamic UDP Increases in Obesity and Promotes Feeding via P2Y6-Dependent Activation of AgRP Neurons. Cell. 162(6): p. 1404–17.10.1016/j.cell.2015.08.032Search in Google Scholar
Steiger, A., et al. (2011). Ghrelin in mental health, sleep, memory. Mol Cell Endocrinol. 340(1): p. 88–96.10.1016/j.mce.2011.02.013Search in Google Scholar
Stellar, E. (1954). The physiology of motivation. Psychol Rev. 61(1): p. 5–22.10.1037/h0060347Search in Google Scholar
Stempel, A.V., et al. (2016). Cannabinoid Type 2 Receptors Mediate a Cell Type-Specific Plasticity in the Hippocampus. Neuron. 90(4): p. 795–809.10.1016/j.neuron.2016.03.034Search in Google Scholar
Stuber, G.D. and R.A. Wise (2016). Lateral hypothalamic circuits for feeding and reward. Nat Neurosci. 19(2): p. 198–205.10.1038/nn.4220Search in Google Scholar
Tsunematsu, T., et al. (2014). Optogenetic manipulation of activity and temporally controlled cell-specific ablation reveal a role for MCH neurons in sleep/wake regulation. J Neurosci. 34(20): p. 6896–909.10.1523/JNEUROSCI.5344-13.2014Search in Google Scholar
Tye, K.M. and K. Deisseroth (2012). Optogenetic investigation of neural circuits underlying brain disease in animal models. Nat Rev Neurosci. 13(4): p. 251–66.10.1038/nrn3171Search in Google Scholar
Vanderwolf, C.H. (1969). Hippocampal electrical activity and voluntary movement in the rat. Electroencephalogr Clin Neurophysiol. 26(4): p. 407–18.10.1016/0013-4694(69)90092-3Search in Google Scholar
Venner, A., et al. (2016). A Novel Population of Wake-Promoting GABAergic Neurons in the Ventral Lateral Hypothalamus. Curr Biol. 26(16): p. 2137–43.10.1016/j.cub.2016.05.078Search in Google Scholar PubMed PubMed Central
Verret, L., et al. (2003). A role of melanin-concentrating hormone producing neurons in the central regulation of paradoxical sleep. BMC Neurosci. 4: p. 19.10.1186/1471-2202-4-19Search in Google Scholar PubMed PubMed Central
Waterson, M.J. and T.L. Horvath (2015). Neuronal Regulation of Energy Homeostasis: Beyond the Hypothalamus and Feeding. Cell Metab. 22(6): p. 962–70.10.1016/j.cmet.2015.09.026Search in Google Scholar PubMed
Wietek, J., et al. (2014). Conversion of channelrhodopsin into a light-gated chloride channel. Science. 344(6182): p. 409–12.10.1126/science.1249375Search in Google Scholar PubMed
Wulff, P., et al. (2009). Hippocampal theta rhythm and its coupling with gamma oscillations require fast inhibition onto parvalbumin-positive interneurons. Proc. Natl. Acad. Sci. U.S.A. 106(9): p. 3561–6.10.1073/pnas.0813176106Search in Google Scholar PubMed PubMed Central
Wu, Z., et al. (2015). GABAergic projections from lateral hypothalamus to paraventricular hypothalamic nucleus promote feeding. J Neurosci. 35(8): p. 3312–8.10.1523/JNEUROSCI.3720-14.2015Search in Google Scholar PubMed PubMed Central
Yamamoto, J., et al. (2014), Successful execution of working memory linked to synchronized high-frequency gamma oscillations. Cell. 157(4): p. 845–57.10.1016/j.cell.2014.04.009Search in Google Scholar PubMed
Yang, J.W., et al. (2009). Three patterns of oscillatory activity differentially synchronize developing neocortical networks in vivo. J Neurosci. 29(28): p. 9011–25.10.1523/JNEUROSCI.5646-08.2009Search in Google Scholar PubMed PubMed Central
Yizhar, O., et al. (2011). Neocortical excitation/inhibition balance in information processing and social dysfunction. Nature. 477(7363): p. 171–8.10.1038/nature10360Search in Google Scholar PubMed PubMed Central
Zhang, F., et al. (2007). Circuit-breakers: optical technologies for probing neural signals and systems. Nat Rev Neurosci. 8(8): p. 577–81.10.1038/nrn2192Search in Google Scholar PubMed
Ziv, Y., et al. (2013). Long-term dynamics of CA1 hippocampal place codes. Nat Neurosci. 16(3): p. 264–6.10.1038/nn.3329Search in Google Scholar PubMed PubMed Central
© 2017 by De Gruyter
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Schlaf, Nahrungsaufnahme und Fortbewegung – Koordination von angeborenem Verhalten durch den lateralen Hypothalamus
- To eat? To sleep? To run? Coordination of innate behaviors by lateral hypothalamus
- Apollos Fluch und Segen: Musizieren als Neuroplastizitätsmotor
- Apollos Gift and Curse: Making Music as a model for Adaptive and Maladaptive Plasticity
- Lust an Gewalt: appetitive Aggression als Teil der menschlichen Natur
- Lust for violence: Appetitive aggression as a fundamental part of human nature
- Bedeutung von Activin für Kognition, Affekt und neuronales Überleben
- Role of activin in cognitive functions, affective behavior and neuronal survival
- Neuronale Vielfalt in der Netzhaut
- Neuronal Diversity In The Retina
- Forschungsförderung
- DFG-Sonderforschungsbereich SFB1193 „Neurobiologie der Resilienz gegenüber stressinduzierter psychischer Dysfunktion: Mechanismen verstehen und Prävention fördern“
- Rezension
- Norman Sieroka: Leibniz, Husserl, and the Brain
- Nachrichten
- Nachrichten
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Schlaf, Nahrungsaufnahme und Fortbewegung – Koordination von angeborenem Verhalten durch den lateralen Hypothalamus
- To eat? To sleep? To run? Coordination of innate behaviors by lateral hypothalamus
- Apollos Fluch und Segen: Musizieren als Neuroplastizitätsmotor
- Apollos Gift and Curse: Making Music as a model for Adaptive and Maladaptive Plasticity
- Lust an Gewalt: appetitive Aggression als Teil der menschlichen Natur
- Lust for violence: Appetitive aggression as a fundamental part of human nature
- Bedeutung von Activin für Kognition, Affekt und neuronales Überleben
- Role of activin in cognitive functions, affective behavior and neuronal survival
- Neuronale Vielfalt in der Netzhaut
- Neuronal Diversity In The Retina
- Forschungsförderung
- DFG-Sonderforschungsbereich SFB1193 „Neurobiologie der Resilienz gegenüber stressinduzierter psychischer Dysfunktion: Mechanismen verstehen und Prävention fördern“
- Rezension
- Norman Sieroka: Leibniz, Husserl, and the Brain
- Nachrichten
- Nachrichten