Anti-stress effects of a GSK-3β inhibitor, AR-A014418, in immobilization stress of variable duration in mice
-
Anjana Bali


Abstract
Background:
The present study was designed to explore the anti-stress role of AR-A014418, a selective glycogen synthase kinase-3β inhibitor (GSK-3β), on changes provoked by immobilization stress of varying duration.
Methods:
Acute stress of varying degree was induced by subjecting mice to immobilization stress of short duration (30 min) or long duration (120 min). Thereafter, these animals were exposed to the same stressor for 5 days to induce stress adaptation. The behavioral alterations were assessed using an actophotometer, a hole-board, and the open field and social interaction tests. The serum corticosterone levels were assessed as markers of the hypothalamic-pituitary-adrenal (HPA) axis activity. The levels of total GSK-3β and p-GSK-3β-S9 were determined in the prefrontal cortex.
Results:
A single exposure to short or long immobilization stress produced behavioral and biochemical changes and the levels of p-GSK-3β-S9 decreased without affecting the total GSK-3β levels in the brain. However, repeated exposure to both short and long stress reversed the behavioral and biochemical changes along with the normalization of p-GSK-3β-S9 levels. The administration of AR-A014418, a selective GSK-3β inhibitor, diminished acute stress-induced behavioral and biochemical changes. Furthermore, AR-A014418 normalized acute stress-induced alterations in p-GSK-3β-S9 levels without changing total GSK-3β levels.
Conclusions:
Our study suggests that acute stress-induced decrease in p-GSK-3β-S9 levels in the brain contributes to the development of behavioral and biochemical alterations and the normalization of GSK-3β signaling may contribute to stress adaptive behavior in mice which have been subjected to repeated immobilization stress.
Introduction
Stress produces widespread changes at the psychological (emotional and cognitive), behavioral (fight or flight) and biological level (autonomic and neuro-endocrine function). Furthermore, it has been assumed to be involved in the pathophysiology of a variety of diseases, including anxiety, depression, post-traumatic stress disorder and memory loss [1]. However, individuals have an inherent capacity to cope with stress in the form of stress adaptation [2, 3] and there is a reduction in the sensitivity of stress responsive elements, including the hypothalamic-pituitary-adrenal (HPA) axis during repeated stress episodes [4]. Indeed, the blunted response of a body to the stress stimulus in the form of stress adaptation is a key protective mechanism against repeated stress exposure [5]. In experimental settings, stress adaptation is characterized in terms of the restoration of behavioral alterations and the normalization of neuroendocrinological changes in comparison with the initial stress response [2, 6]. Despite the knowledge of the existence of stress adaptation in animals and humans, the precise molecular signaling pathways involved in stress adaptive processes are not delineated.
Glycogen synthase kinase-3 (GSK-3), a serine/threonine kinase, exists in two isoforms, α and β, and is widely expressed in the stress-sensitive brain regions, including the amygdala, prefrontal cortex and hippocampus [7]. It has also been implicated in the development of neuronal disorders such as neuroinflammation, Alzheimer’s disease, schizophrenia and bipolar disorder [8]. Research evidence indicates that GSK-3β activity is inhibited by phosphorylation at serine 9, and this inhibition exerts protective effects and increases neuroplasticity [7]. It has also been reported that GSK-3β phosphorylation is reduced in patients with schizophrenia, leading to increased GSK-3β activity. Moreover, antipsychotic drugs increase the amount of phosphorylated GSK-3β [9]. Exposure to stressors including immobilization, swim and restraint stress has been shown to increase the GSK-3β activity in the hippocampus and frontal cortex of animals [10, 11]. In addition, studies have indicated that the hyperactivity of GSK-3β may contribute to depression [12, 13]. The administration of flupirtine (a centrally acting, non-opioid analgesic) attenuates the stress-induced deleterious effects by decreasing GSK-3β activity, suggesting the role of activation of GSK-3β signaling pathways during stress induction [14]. The administration of lithium, a GSK-3β inhibitor, potentiates the antidepressant-like effects of ketamine in a mouse model of stress [15]. However, there has also been a study showing that the administration of a GSK-3β inhibitor does not affect the immobility time in chronic unpredictable stress-subjected animals [16]. Therefore, the present study was designed to explore the stress attenuating potential of the selective GSK-3β inhibitor, AR-A014418, on immobilization stress-subjected animals of variable duration.
Materials and methods
Animals and drugs
Swiss albino mice weighing 25 g±5 g were used in the present study and fed on the standard laboratory feed and water. The animals were housed in the departmental animal house and were exposed to natural cycles of 12 h light and dark. The experimental protocol was approved by the Institutional Animal Ethics Committee (IAEC) and care of the animals was carried out as per the guidelines of the Committee for the Purpose of Control and Supervision of Experiments on Animals (CPCSEA), Ministry of Environment and Forests, Government of India (Reg. No. CPCSEA/107/1999). All experiments were done between 8:30 a.m. and 12:00 a.m., when the levels of HPA axis hormones are stable [5].
AR-A014418 was purchased from Cayman Chemical Co., USA and dissolved in normal saline containing 4% DMSO, while diethyldithiocarbamic acid was procured from Sigma Aldrich, USA. The Elisa kit for estimation of serum corticosterone was purchase from Arbor assays, USA and GSK-3β Elisa kit was purchase from Ray biotech Pvt. Ltd, USA. The statistical analysis was performed by using Graph Pad prism software 6.0. The behavioral apparatus were purchased from Imcorp, Ambala, India.
Immobilization stress and stress adaptation
The animals were subjected to immobilization stress by immobilizing the mice in a prone position with their limbs stretched on a board using adhesive tape. The animals were immobilized either for 30 min (short duration stress) or 120 min (long duration stress) [17]. The movement of the head was also restricted by keeping the head in a metal loop coiled around the neck [18, 19]. The mice were exposed to homotypic immobilization stress for 5 days to induce adaptive behavior.
Acclimatization of animals
Before initiating the actual experimentation, the mice were acclimatized for 5 min per day for 3 days on the behavioral test apparatus. Such acclimatization of animals to the test apparatus potentially avoids the confounding effects due to the novelty of the test apparatus and thus reduces variations in the experimental data [3, 20]. The animals were also habituated to a blood withdrawal procedure by subjecting them to a tail vein nick under resting conditions.
Assessment of stress and stress adaptation
Behavioral parameters:
Ten min after the immobilization stress protocol, a series of behavioral tests were performed on the animals in the following sequence: actophotometer, hole-board, open field and social interaction tests, with a time gap of 5 min between each test. The behavioral test equipment was cleaned after each test with alcohol and water. The behavior in the hole-board, open field and social interaction tests was video-recorded and then analyzed.
Actophotometer test
In an actophotometer, the movement of an animal interrupts a beam of light falling on a photocell and a count is recorded digitally. Therefore, the number of counts is directly related to the movement of animals inside the actophotometer chamber. The animals were placed in the actophotometer for 10 min and their activity was assessed in terms of count per 10 min [2, 21].
Open field test
The open field test was employed to assess the stress-related behavior in rodents on the basis of changes in the exploration, general locomotor activity and spontaneous activity. Each mouse was exposed to the open field test for 10 min in a dimly light room. The mice were placed in the center of the open field and the number of line crossings and rearings were noted [2].
Hole-board test
A mouse was placed in the center of the hole-board and was allowed to explore the apparatus for 10 min. The animals were assessed for head dipping and rearing [2, 22]. The low number of head dipping reflects the high anxiety state level and the number of rearings represents the exploration in the novel surrounding.
Social interaction test
The social interaction test was used to assess the anxiety-related behavior. The social interaction test was carried out in the same box in which the open field test was performed. During a 10 min test, each experimental mouse was allowed to interact with a partner mouse, which was socially housed and not subjected to any stressor. The behavior of a test mouse with a test partner such as close proximity, facing and sniffing the partner, contact interaction (physical contact) and climbing over was considered to be social behavior. The remaining time interval was considered to be non-social behavior, which included actively turning away, keeping the partner at a distance with forepaws in an upright posture, freezing, self-grooming and remaining alone [2, 22].
Serum corticosterone levels
A blood sample was taken using a tail vein nick procedure which consisted of gently placing the animal in a restrainer and making a 2 mm incision at the end of the tail vein to collect the blood. This method is extensively used in various laboratories [5, 23] and the hormone levels are maintained very close to the resting levels, provided that the animal has had previous experience with the procedure. Thereafter, the serum was isolated by centrifugation and samples were assayed for corticosterone using a Corticosterone Enzyme Immunoassay kit (K014-H1, Arbor Assay).
Enzyme-linked immunosorbent assay (ELISA)
Following sacrifice, the prefrontal cortex region of the brain was isolated and separately homogenized in 1 mL of lysis buffer containing protease and phosphatase inhibitors. The homogenates were used immediately for ELISA-based estimation of total GSK-3β and p-GSK-3β-S9 levels using a San microplate reader and the procedure was conducted in accordance with the instructions of the manufacturer.
Experimental protocol
Ten groups, each comprising eight Swiss albino mice, were employed in the study. Due to ethical concerns, all animals from these ten groups were not sacrificed. Only animals belonging to seven representative groups (from group I to group VII) were sacrificed (five animals from each group) to isolate the prefrontal cortex.
Group I: Non-stress:
Non-stressed mice were subjected to different behavioral tests on day 1 of the corresponding immobilization stress group. The blood was withdrawn on the same day and afterwards, the animals were sacrificed to isolate the prefrontal cortex.
Group II: Acute short immobilization stress:
In this paradigm, mice were immobilized for 30 min to produce acute stress. The blood was withdrawn on the same day and afterwards the animals were sacrificed to isolate the prefrontal cortex.
Group III: Repeated short immobilization stress:
In this paradigm, mice were repeatedly exposed to 30 min immobilization stress for 5 consecutive days to induce stress adaptation. The behavioral and biochemical parameters were assessed on the 5th day of stress paradigm.
Group IV: Acute long immobilization stress:
In this paradigm, mice were immobilized for 120 min to produce acute stress. The rest of the procedure was as described in group II.
Group V: Repeated long immobilization stress:
In this paradigm, mice were subjected to repeated immobilization stress of 120 min to induce stress adaptation. The behavioral and biochemical parameters were assessed on the 5th day of stress exposure.
Groups VI: AR-A014418 (3 mg/kg) in acute short immobilization stress:
AR-A014418 (3 mg/kg) was administered 30 min prior to the exposure of acute immobilization stress and thereafter behavioral tests were conducted followed by isolation of blood.
Groups VII: AR-A014418 (3 mg/kg i.p.) in acute long immobilization stress:
AR-A014418 (3 mg/kg) was administered 30 min prior to the exposure to acute immobilization stress and thereafter behavioral tests were conducted followed by isolation of blood.
Group VIII: DMSO (vehicle) in acute short immobilization stress:
Four percent DMSO dissolved in normal saline (vehicle for AR-A014418) was administered (0.1 mL/10 g) 30 min prior to exposure to acute short immobilization stress. Thereafter, behavioral tests were conducted followed by isolation of blood.
Group IX: DMSO (vehicle) in acute long immobilization stress:
Four percent DMSO dissolved in normal saline (vehicle for AR-A014418) was administered (0.1 mL/10 g) 30 min prior to exposure to acute long immobilization stress. Thereafter, behavioral tests were conducted followed by isolation of blood.
Group X: AR-A014418 (3 mg/kg i.p.) per se:
AR-A014418 was administered to non-stressed mice. Thereafter, behavioral tests were conducted followed by isolation of blood.
Statistical analysis
The results were expressed as mean±standard deviation (SD). The results were analyzed using one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test for comparison between the different groups. The p value<0.05 was considered to be statistically significant.
Results
The administration of the vehicle did not modulate short and long immobilization stress-induced changes in behavior and serum corticosterone levels in a significant manner. Per se administration of AR-A014418 (3 mg/kg) in non-stressed mice did not modulate the behavior and serum corticosterone levels in a significant manner.
The effect of AR-A014418 on short and long immobilization stress-induced alterations in the locomotor activity
The acute exposure to short and long immobilization stress decreased the locomotor activity on day 1 as compared to non-stress control mice in a significant manner. However, repeated exposure to short or long immobilization stress for 5 consecutive days led to significant restoration in the locomotor activity as compared to day 1 of acute short and long stress exposure. A single administration of AR-A014418 (3 mg/kg) significantly restored acute short and long immobilization stress-induced decrease in locomotor activity on the first day of acute stress exposure (Figure 1).

The assessment of locomotor activity in terms of counts in 10 min time intervals in the actophotometer test for evaluating the stress adaptive effect of short duration and long duration immobilization stress and its pharmacological modulation with AR-A014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with stress control.
The effect of AR-A014418 on short and long immobilization stress-induced alterations in other behavioral parameters
The acute exposure to immobilization stress led to a significant decrease in other behavioral activities including the number of rearings and head dips in the hole-board (Figures 2 and 3 ), total motor activity, i.e. line crossings and rearing in the open field (Figures 4 and 5 ) and social behavior (Table 1) on day 1 as compared to non-stress control. Similarly, on repeated stress exposure to short and long immobilization stress for 5 consecutive days, the behavioral activities were significantly restored on the 5th day of stress exposure. A single administration of AR-A014418 significantly restored acute short and long stress-induced behavioral deficits on the first day of stress exposure.
The assessment of social and non-social behavior in the 10 min social interaction test by noting time of following and avoiding the partner for investigating the stress adaptation in short and long duration immobilization stress and its pharmacological modulation with AR014418.
| Sample number | Experimental groups | Social interaction tests | |
|---|---|---|---|
| Following time(s) | Avoidance time(s) | ||
| 1 | Non-stress | 528.0±11.8 | 72.0±11.8 |
| 2 | Acute short immobilization stress | 110.0±17.3a | 490.0±17.3a |
| 3 | Repeated short immobilization stress | 495.0±9.5b | 105.0±9.5b |
| 4 | AR-A014418 (3 mg/kg) in acute short immobilization stress | 490.5±15.6b | 109.5±15.6b |
| 5 | Acute long immobilization stress | 89.5±16.3a′ | 510.5±16.3a′ |
| 6 | Repeated long immobilization stress | 520.4±9.9b′ | 79.6±9.9b′ |
| 7 | AR-A014418 (3 mg/kg) in acute long immobilization stress | 496.5±13.5b′ | 103.5±13.5b′ |
| 8 | DMSO (vehicle) in acute short immobilization stress | 107.5±8.2 | 492.5±8.2 |
| 9 | DMSO (vehicle) in acute long immobilization stress | 98.5±9.8 | 501.5±9.8 |
Values are expressed as mean±SD. ap<0.05 as compared with normal control of day 1 for acute short duration immobilization stress; bp<0.05 as compared with acute short duration stress control; a′p<0.05 as compared with normal control of day 1 for acute long duration immobilization stress; b′p<0.05 as compared with acute long duration stress control.

The assessment of the exploratory behavior of animals in terms of head dips in 10 min time intervals in the hole-board test for investigating the stress adaptive effect of short duration and long duration immobilization stress and its pharmacological modulation with AR-A014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with stress control.

The assessment of the exploratory behavior of animals in terms of the number of rearings in 10 min time intervals in the hole-board test for investigating the stress adaptive effect of short duration and long duration immobilization stress and its pharmacological modulation with AR-A014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with stress control.

The assessment of spontaneous, exploratory and ambulatory activity in terms of the frequency of line crossing in 10 min time intervals in the open field test for investigating the stress adaptation in short duration and long duration immobilization stress and its pharmacological modulation with AR-A014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared to normal control; bp<0.05 as compared to stress control.
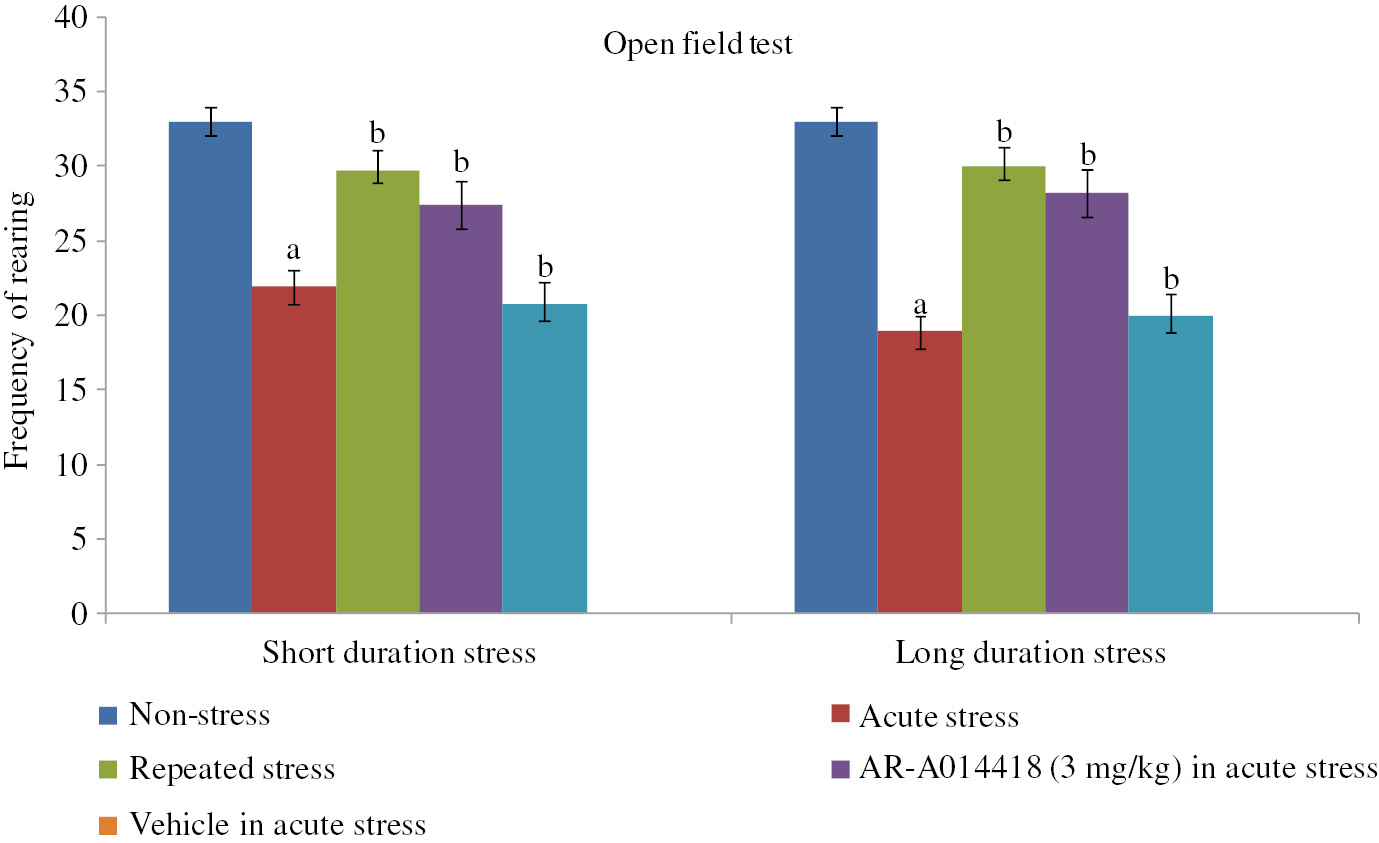
The assessment of spontaneous, exploratory and ambulatory activity in terms of the frequency of rearings in 10 min time intervals in the open field test for investigating the stress adaptation in short duration and long duration immobilization stress and its pharmacological modulation with AR-A014418. Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with stress control.
The effect of AR-A014418 and repeated stress exposure on immobilization stress-induced serum corticosterone levels
The acute exposure to short and long immobilization stress produced a significant increase in the serum corticosterone levels as compared to non-stress control mice. However, following repeated exposures of short and long immobilization stress, the serum corticosterone levels were restored on the 5th day and the levels were comparable to the levels of corticosterone on the 5th day of non-stress control. A single administration of AR-A014418 also restored the acute short and long stress-induced increases in corticosterone levels (Figure 6).

The estimation of serum corticosterone levels for investigating the stress adaptation in short and long duration immobilization stress and its pharmacological modulation with AR-A014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with stress control.
The effect of AR-A014418 and repeated stress exposure on immobilization stress-induced pGSK-3β expression levels
The acute short and long immobilization stress exposure significantly decreased the p-GSK-3β-S9 levels in the prefrontal cortex in comparison to non-stressed mice (Figure 7). However, on repeated exposure to the same stress for 5 days, the p-GSK-3β-S9 levels were significantly restored in the prefrontal cortex as compared to acute short and long stress subjected mice (Figure 8). A single administration of AR-A014418 significantly restored acute short and long stress-induced decreases in the levels of p-GSK-3β-S9 in the prefrontal cortex (Figures 7 and 8).

The assessment of the p-GSK-3β-S9 levels for investigating the stress adaptive effect of short immobilization stress and its pharmacological modulation with AR-AR014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with acute immobilization stress.
Discussion
In the present study, a single exposure of immobilization stress of either short or long duration produced the significant behavioral alterations assessed in terms of a decrease in locomotor, spontaneous and social activities. Furthermore, a single exposure to short as well as long duration immobilization stress also led to an increase in the serum corticosterone levels, suggesting the over-activation of the HPA axis [24]. The observed behavioral as well as biochemical changes in the present study are in agreement with other studies showing an alteration in behavior and an increase in serum corticosterone levels in response to acute immobilization stress exposure [5, 25, 26]. However, on repeated exposure to immobilization stress for 5 consecutive days, there was significant restoration of both behavioral and biochemical alterations indicating the induction of stress adaptation. The development of stress adaptation in the present study is consistent with our previous study showing the restoration of behavioral and biochemical changes following repeated applications of immobilization stress for 5 days [25].
Furthermore, a single dose administration of AR-A014418 significantly attenuated acute immobilization in short as well as long duration stress-induced alterations in behavior and corticosterone levels. AR-A014418 is a selective GSK-3β inhibitor with IC50=104 nM. It exhibits high specificity for GSK-3 over cdk2 and cdk5 (IC50 values are >100 μM) and over 26 other kinases. The administration of AR-A014418 has been shown to attenuate the development of learning deficits in diabetic mice [22]. Another study has also reported the neuronal antiapoptotic and neuroprotective effects of AR-A014418 in the traumatic spinal cord injury model in rats [12]. In addition, AR-A014418 has also shown growth suppressive effects in pancreatic cancer cells [27]. However, to the best of our knowledge, this is the first study describing the stress attenuating potential of AR-A014418, a selective GSK-3β inhibitor, in immobilization stress of varying duration in mice.
In the present investigation, a single exposure of short as well as long immobilization stress-induced behavioral deficits was accompanied by a decrease in p-GSK-3β-S9, without any alteration in the total GSK-3β levels in the prefrontal cortex, suggesting that the changes in GSK-3β activity may be secondary to change in its phosphorylation state. An earlier study has shown that the exposure of acute restraint stress is associated with the decrease in p-GSK-3β-Ser9 in the hippocampus region of mice [28]. GSK-3β, a serine/threonine kinase, is an essential downstream effector of Akt and its activity is significantly reduced by Akt-mediated phosphorylation [29]. However, in the present study, acute stress exposure did not change the levels of total GSK-3β in the prefrontal cortex, suggesting that stress-induced changes in the GSK-3β activity may be due to a change in its phosphorylation state. Accordingly, it is possible to suggest that acute stress exposure may inhibit the Akt activity, which in turn may decrease the phosphorylation of GSK-3β to increase its activity. An earlier study has shown the decrease in Akt activity in the brain regions in response to acute stress exposure [30] (Figures 9 and 10 ).

The assessment of the p-GSK-3β-S9 levels for investigating the stress adaptive effect of long immobilization stress and its pharmacological modulation with AR-AR014418.
Values are expressed as mean±SD. Data were analyzed by one-way ANOVA followed by post hoc analysis using Tukey’s multiple comparison test. ap<0.05 as compared with normal control; bp<0.05 as compared with acute immobilization stress.

The representation of the signaling cascade involving GSK-3β during stress induction.

The representation of the signaling cascade involving GSK-3β during stress adaptation.
Our previous studies described the potential stress adaptive role of angiotensin II in short duration immobilization stress and the role of opioids in long duration immobilization stress [25, 31]. However, the present study results demonstrate the involvement of the same intracellular signaling cascade, i.e. GSK-3β in the induction of stress adaptation irrespective of duration of immobilization stress (short as well as long). Accordingly, it may be proposed that although the upstream modulators may be different, i.e. angiotensin II and the opioids in immobilization stress of variable duration, yet the downstream regulatory system may converge and the same signaling cascade may be involved in inducing stress adaptive behavior. The previous study from our laboratory described the involvement of the GSK-3β signaling cascade in electric foot shock stress-induced stress adaptation [32]. Based on this, it may be proposed that irrespective of stressor duration (short or long) or stressor type (immobilization or electric foot shock), the down-stream signaling cascade is the same and may possibly involve GSK-3β.
Conclusions
Acute immobilization stress exposure of short and long duration was associated with a decrease in p-GSK-3β-S9 levels in the prefrontal cortex, suggesting that the up-regulation of GSK-3β signaling may possibly contribute to the development of behavioral and biochemical alterations and its normalization contributes to stress adaptive behavior in response to repeated immobilization stress-subjected mice. The anti-stress effects of AR-A014418 projects GSK-3β as an important target in modulating stress associated behavior.
Acknowledgments
The authors are grateful to the Department of Pharmaceutical Sciences and Drug Research, Punjabi University, Patiala, India for supporting this study and providing technical facilities for the work.
Author contributions: All the authors have accepted responsibility for the entire content of this submitted manuscript and approved submission.
Research funding: None declared.
Employment or leadership: None declared.
Honorarium: None declared.
Competing interests: The funding organization(s) played no role in the study design; in the collection, analysis, and interpretation of data; in the writing of the report; or in the decision to submit the report for publication.
References
1. Rasheed N, Ahmad A, Pandey CP, Chaturvedi RK, Lohani M, Palit G. Differential response of central dopaminergic system in acute and chronic unpredictable stress models in rats. Neurochem Res 2010;35:22–32.10.1007/s11064-009-0026-5Search in Google Scholar
2. Agrawal A, Jaggi AS, Singh N. Pharmacological investigations on adaptation in rats subjected to cold water immersion stress. Physiol Behav 2011;103:321–9.10.1016/j.physbeh.2011.02.014Search in Google Scholar
3. Chauhan E, Bali A, Singh N, Singh Jaggi A. Pharmacological investigations on cross adaptation in mice subjected to stress immobilization. Life Sci 2015;127:98–105.10.1016/j.lfs.2015.02.015Search in Google Scholar
4. Pfister HP, King MG. Adaptation of the glucocorticosterone response to novelty. Physiol Behav 1976;17:43–46.10.1016/0031-9384(76)90267-5Search in Google Scholar
5. Rabasa C, Muñoz-Abellán C, Daviu N, Nadal R, Armario A. Repeated exposure to immobilization or two different footshock intensities reveals differential adaptation of the hypothalamic-pituitary-adrenal axis. Physiol Behav 2011;103:125–33.10.1016/j.physbeh.2011.02.022Search in Google Scholar
6. Cohen RM, Aulakh CS, Murphy DL. Long-term clorgyline treatment antagonizes the eating and motor function responses to m-chlorophenylpiperazine. Eur J Pharmacol 1983;94:175–9.10.1016/0014-2999(83)90459-4Search in Google Scholar
7. GouldTD,ManjiHK. Glycogen synthase kinase-3: a putative molecular target for lithium mimetic drugs. Neuropsychopharmacology 2005;30:1223–37.10.1038/sj.npp.1300731Search in Google Scholar PubMed
8. GouldTJ,Rukstalis M,Lewis MC. Atomoxetine and nicotine enhance prepulse inhibition of acoustic startle in C57BL/6 mice. Neurosci Lett2005;377:85–90.10.1016/j.neulet.2004.11.073Search in Google Scholar PubMed
9. Sutton LP, Honardoust D, Mouyal J, Rajakumar N, Rushlow WJ. Activation of the canonical Wnt pathway by the antipsychotics haloperidol and clozapine involves dishevelled-3. J Neurochem 2007;102:153–69.10.1111/j.1471-4159.2007.04527.xSearch in Google Scholar PubMed
10. Chen YC, Tan QR, Dang W, Wang HN, Zhang RB, Li ZY, et al. The effect of citalopram on chronic stress-induced depressive-like behavior in rats through GSK3β/β-catenin activation in the medial prefrontal cortex. Brain Res Bull 2012;88:338–44.10.1016/j.brainresbull.2012.03.004Search in Google Scholar PubMed
11. Seo MK, Lee CH, Cho HY, You YS, Lee BJ, Lee JG, et al. Effects of antipsychotic drugs on the expression of synapse-associated proteins in the frontal cortex of rats subjected to immobilization stress. Psychiatry Res 2015;229:968–74.10.1016/j.psychres.2015.05.098Search in Google Scholar PubMed
12. Gould TD, Einat H, Bhat R, Manji HK. AR-A014418, a selective GSK-3 inhibitor, produces antidepressant-like effects in the forced swim test. Int J Neuropsychopharmacol 2004;7:387–90.10.1017/S1461145704004535Search in Google Scholar PubMed
13. Zhang K, Song X, Xu Y, Li X, Liu P, Sun N, et al. Continuous GSK-3β overexpression in the hippocampal dentate gyrus induces prodepressant-like effects and increases sensitivity to chronic mild stress in mice. J Affect Disord 2013;146:45–52.10.1016/j.jad.2012.08.033Search in Google Scholar PubMed
14. Huang P, Li C, Fu T, Zhao D, Yi Z, Lu Q, et al. Flupirtine attenuates chronic restraint stress-induced cognitive deficits and hippocampal apoptosis in male mice. Behav Brain Res 2015;288:1–10.10.1016/j.bbr.2015.04.004Search in Google Scholar PubMed
15. Chiu CT, Scheuing L, Liu G, Liao HM, Linares GR, Lin D, et al. The mood stabilizer lithium potentiates the antidepressant-like effects and ameliorates oxidative stress induced by acute ketamine in a mouse model of stress. Int J Neuropsychopharmacol 2014;18. doi: 10.1093/ijnp/pyu102.10.1093/ijnp/pyu102Search in Google Scholar PubMed PubMed Central
16. Ma XC, Dang YH, Jia M, Ma R, Wang F, Wu J, et al. Long-lasting antidepressant action of ketamine, but not glycogen synthase kinase-3 inhibitor SB216763, in the chronic mild stress model of mice. PLoS One 2013;8:e56053.10.1371/journal.pone.0056053Search in Google Scholar PubMed PubMed Central
17. Ricart-Jané D, Rodríguez-Sureda V, Benavides A, Peinado-Onsurbe J, López-Tejero MD, Llobera M. Immobilization stress alters intermediate metabolism and circulating lipoproteins in the rat. Metabolism 2002;51:925–31.10.1053/meta.2002.33353Search in Google Scholar PubMed
18. Kvetnansky R, Mikulaj L. Adrenal and urinary catecholamines in rats during adaptation to repeated immobilization stress. Endocrinology 1970;87:738–43.10.1210/endo-87-4-738Search in Google Scholar PubMed
19. Bhatia N, Jaggi AS, Singh N, Anand P, Dhawan R. Adaptogenic potential of curcumin in experimental chronic stress and chronic unpredictable stress-induced memory deficits and alterations in functional homeostasis. J Nat Med 2011;65:532–43.10.1007/s11418-011-0535-9Search in Google Scholar PubMed
20. Lapiz-Bluhm MD, Bondi CO, Doyen J, Rodriguez GA, Bédard-Arana T, Morilak DA. Behavioural assays to model cognitive and affective dimensions of depression and anxiety in rats. J Neuroendocrinol 2008;20:1115–37.10.1111/j.1365-2826.2008.01772.xSearch in Google Scholar PubMed PubMed Central
21. Kaur R, Jaggi AS, Singh N. Studies on effect of stress preconditioning in restrain stress-induced behavioral alterations. Yakugaku Zasshi 2010;130:215–21.10.1248/yakushi.130.215Search in Google Scholar PubMed
22. King MR, Anderson NJ, Guernsey LS, Jolivalt CG. Glycogen synthase kinase-3 inhibition prevents learning deficits in diabetic mice. J Neurosci Res 2013;91:506–14.10.1002/jnr.23192Search in Google Scholar PubMed PubMed Central
23. García A, Martí O, Vallès A, Dal-Zotto S, Armario A. Recovery of the hypothalamic-pituitary-adrenal response to stress. Effect of stress intensity, stress duration and previous stress exposure. Neuroendocrinology 2000;72:114–25.10.1159/000054578Search in Google Scholar PubMed
24. Bali A, Jaggi AS. Preclinical experimental stress studies: protocols, assessment and comparison. Eur J Pharmacol 2015;746:282–92.10.1016/j.ejphar.2014.10.017Search in Google Scholar PubMed
25. Bali A, Jaggi AS. Investigations in foot shock stress of variable intensity in mice: Adaptation and role of angiotensin II. Eur J Pharmacol 2015;761:86–94.10.1016/j.ejphar.2015.04.037Search in Google Scholar PubMed
26. Kumar N, Singh N, Jaggi AS. Anti-stress effects of cilnidipine and nimodipine in immobilization subjected mice. Physiol Behav 2012;105:1148–55.10.1016/j.physbeh.2011.12.011Search in Google Scholar PubMed
27. Kunnimalaiyaan S, Gamblin TC, Kunnimalaiyaan M. Glycogen synthase kinase-3 inhibitor AR-A014418 suppresses pancreatic cancer cell growth via inhibition of GSK-3-mediated Notch1 expression. HPB (Oxford) 2015;17:770–6.10.1111/hpb.12442Search in Google Scholar PubMed PubMed Central
28. Jin Y, Kanno T, Nishizaki T. Acute restraint stress impairs induction of long-term potentiation by activating GSK-3β. Neurochem Res 2015;40:36–40.10.1007/s11064-014-1462-4Search in Google Scholar PubMed
29. Jope RS, Johnson GV. The glamour and gloom of glycogen synthase kinase-3. Trends Biochem Sci 2004;29:95–102.10.1016/j.tibs.2003.12.004Search in Google Scholar PubMed
30. Demarchi F, Bertoli C, Sandy P, Schneider C. Glycogen synthase kinase-3 beta regulates NF-kappa B1/p105 stability. J Biol Chem 2003;278:39583–90.10.1074/jbc.M305676200Search in Google Scholar PubMed
31. Bali A, Jaggi AS. Differential role of angiotensin neuropeptides in repeated exposure of immobilization stress of varying duration in mice. Life Sci 2015;141:90–98.10.1016/j.lfs.2015.09.005Search in Google Scholar PubMed
32. Bali A, Jaggi AS. Investigations on GSK-3β/NF-kB signaling in stress and stress adaptive behavior in electric foot shock subjected mice. Behav Brain Res 2016;302:1–10.10.1016/j.bbr.2016.01.014Search in Google Scholar PubMed
©2017 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Frontmatter
- Editorial
- The double face of light effects: circadian adjustment or disruption
- Review
- Artificial light-at-night – a novel lifestyle risk factor for metabolic disorder and cancer morbidity
- Behavior and Neuroprotection
- Anti-stress effects of a GSK-3β inhibitor, AR-A014418, in immobilization stress of variable duration in mice
- Cardiovascular Function
- Addition of omega-3 fatty acid and coenzyme Q10 to statin therapy in patients with combined dyslipidemia
- Oxidative Stress
- Protective effect of Moringa oleifera oil against HgCl2-induced hepato- and nephro-toxicity in rats
- Effects of peripherally and centrally applied ghrelin on the oxidative stress induced by renin angiotensin system in a rat model of renovascular hypertension
- Investigation of the role of α-lipoic acid on fatty acids profile, some minerals (zinc, copper, iron) and antioxidant activity against aluminum-induced oxidative stress in the liver of male rats
- Metabolism
- The choice of freely preferred cadence by trained nonprofessional cyclists may not be characterized by mechanical efficiency
- Immune Response
- Ascorbic acid does not modulate potassium currents in cultured human lymphocytes
- Phytotherapy
- Anti-inflammatory activity of Elaeagnus angustifolia fruit extract on rat paw edema
- Histopathological and biochemical assessments of Costus afer stem on alloxan-induced diabetic rats
- In vitro inhibition of phosphodiesterase-5 and arginase activities from rat penile tissue by two Nigerian herbs (Hunteria umbellata and Anogeissus leiocarpus)
- Antioxidant and antiproliferative potentials of methanol extract of Xylopia aethiopica (Dunal) A. Rich in PC-3 and LNCaP cells
Articles in the same Issue
- Frontmatter
- Editorial
- The double face of light effects: circadian adjustment or disruption
- Review
- Artificial light-at-night – a novel lifestyle risk factor for metabolic disorder and cancer morbidity
- Behavior and Neuroprotection
- Anti-stress effects of a GSK-3β inhibitor, AR-A014418, in immobilization stress of variable duration in mice
- Cardiovascular Function
- Addition of omega-3 fatty acid and coenzyme Q10 to statin therapy in patients with combined dyslipidemia
- Oxidative Stress
- Protective effect of Moringa oleifera oil against HgCl2-induced hepato- and nephro-toxicity in rats
- Effects of peripherally and centrally applied ghrelin on the oxidative stress induced by renin angiotensin system in a rat model of renovascular hypertension
- Investigation of the role of α-lipoic acid on fatty acids profile, some minerals (zinc, copper, iron) and antioxidant activity against aluminum-induced oxidative stress in the liver of male rats
- Metabolism
- The choice of freely preferred cadence by trained nonprofessional cyclists may not be characterized by mechanical efficiency
- Immune Response
- Ascorbic acid does not modulate potassium currents in cultured human lymphocytes
- Phytotherapy
- Anti-inflammatory activity of Elaeagnus angustifolia fruit extract on rat paw edema
- Histopathological and biochemical assessments of Costus afer stem on alloxan-induced diabetic rats
- In vitro inhibition of phosphodiesterase-5 and arginase activities from rat penile tissue by two Nigerian herbs (Hunteria umbellata and Anogeissus leiocarpus)
- Antioxidant and antiproliferative potentials of methanol extract of Xylopia aethiopica (Dunal) A. Rich in PC-3 and LNCaP cells