
Insights into caudate amphibian skin secretions with a focus on the chemistry and bioactivity of derived peptides
-
Lorena Kröner
Lorena Kröner received her M.Ed. in Chemistry and Mathematics from University of Koblenz in 2023. Currently, she is a Ph.D. student in Bioorganic Chemistry at the University of Koblenz. Her current research, under the supervision of Jun.-Prof. Marie-T. Hopp, is focused on the investigation of amphibian skin molecules and their physiological role.
Stefan Lötters is a professor of Biogeography at Trier University, leading an Ecology, Evolution & Systematics Group. Using amphibians as a main model group, his research is focused on speciation and character evolution with a particular geographic focus on the Neotropics, as well as the response to new pathogens. The latter is a serious conservation issue in many amphibians including salamanders and newts in Europe.
Marie-T. Hopp obtained her master’s degree in Chemistry and Biology (2017) at the University of Koblenz-Landau. In 2021, she completed her PhD in Pharmaceutical Biochemistry and Bioanalytics at the University of Bonn. In 2023, she joined the University of Koblenz as a junior professor for Bioorganic Chemistry with a focus on peptide chemistry. She aims to characterize heme-protein interactions in the blood coagulation system as well as to explore the chemistry and bioactivity of amphibian skin peptides at molecular level.



Abstract
Amphibians are well-known for their ability to produce and secrete a mixture of bioactive substances in specialized skin glands for the purpose of antibiotic self-protection and defense against predators. Some of these secretions contain various small molecules, such as the highly toxic batrachotoxin, tetrodotoxin, and samandarine. For some time, the presence of peptides in amphibian skin secretions has attracted researchers, consisting of a diverse collection of – to the current state of knowledge – three to 104 amino acid long sequences. From these more than 2000 peptides many are known to exert antimicrobial effects. In addition, there are some reports on amphibian skin peptides that can promote wound healing, regulate immunoreactions, and may serve as antiparasitic and antioxidative substances. So far, the focus has mainly been on skin peptides from frogs and toads (Anura), eclipsing the research on skin peptides of the ca. 700 salamanders and newts (Caudata). Just recently, several novel observations dealing with caudate peptides and their structure-function relationships were reported. This review focuses on the chemistry and bioactivity of caudate amphibian skin peptides and their potential as novel agents for clinical applications.
1 Introduction
Modern amphibians are of Jurassic origin with three extant orders, Gymnophiona (caecilians), Caudata (salamander and newts), and Anura (frogs and toads) with about 8800 species. They are the only tetrapods with a free larval stage that commonly lives in an aquatic environment, while amphibian adults are mostly terrestrial (Brown and Cai 2007; Çömden et al. 2023). The changing environment during the biphasic life cycle confronts especially their skin with many challenges due to environmental factors, such as predators, microorganisms, and several physical factors (Çömden et al. 2023). As a response, the skin shows special features such as multicellular exocrine glands (i.e., mucous and granular glands) for the production, storage, and release of secretions (Çömden et al. 2023; Pereira et al. 2018; Toledo and Jared 1995). While the smaller mucous glands dispense a sticky mucus over the skin as a lubricant with benefits for thermoregulation, water balance, and cutaneous respiration among others, the granular glands are responsible for the production and/or storage of bioactive substances, like toxins for defense against predators (Çömden et al. 2023; Toledo and Jared 1995). Chemically, the small molecule components of amphibian skin secretions are well characterized, such as the potent toxic alkaloid batrachotoxin in poison frogs (Phyllobates) or tetrodotoxin (TTX) and its analogs in, for example, harlequin toads (Atelopus) or rough-skinned newts (Taricha) (Daly et al. 2005; Hanifin 2010; Saporito et al. 2012). After the discovery of the first peptides [e.g., bradykinins (Anastasi et al. 1965), bombinin (Csordaś and Michl 1970), and bombesin (Anastasi et al. 1971)] in anuran skin secretions in the 1960s and 1970s, meanwhile > 2000 peptides have been isolated and characterized from amphibian skin (Xu and Lai 2015). These were described to exert different functions, among them antimicrobial, wound-healing, antioxidant, opioid-like, antiviral, pheromonal, and antiparasitic activities (Xu and Lai 2015). The majority of these peptides have been found on the skin of anurans. Comparatively little is known about skin peptides from caudates. Despite this, there are reports about different mucus components, like bioactive skin peptides, from salamanders and newts with high biopharmaceutical potential. Thus, the goal of this review is to present an overview of the skin components of caudates with a focus on the chemistry and bioactivity of skin peptides.
2 Chemical composition of caudate amphibian skin secretions: a general overview
2.1 Lipids: fatty acids, steroids, and derivatives
So far, the presence of lipids in skin secretions (Figure 1) was only confirmed for a few species of three families, Salamandra atra, Salamandra infraimmaculata, Salamandra salamandra, and Triturus carnifex, all of the Salamandridae, Plethodon cinereus of the Plethodontidae, as well as Hynobius leechii of the Hynobiidae.
![Figure 1:
Chemical composition of caudate amphibian skin secretions. On the skin of caudates, several different classes of substances were identified, encompassing fatty acids and cholesterol derivatives (i.e., 1: Palmitoleic acid, 2: myristoleic acid, 3: linolenic acid, 4: cholesterol, 5: cholesteryl stearate, 6: cholesteryl oleate, 7: cholesteryl palmitate, 8: (2α,3β)-2,3-bis[(4-carboxy-3,3-dimethylbutanoyl) oxy]olean-12-en-28-oic acid, 9: dihydronandrolone, 10: (15Z)-9,12,13-trihydroxy-15-octadecenoic acid, 11: nebrosteroid L, 12: androstenedione, 13: 16-alpha-17-beta-estriol, 14: minabeolide-8, 15: taurochenodeoxycholic acid, 16: cholic acid, 17: palmitoyl glycine, 18: N-arachidonoyl serine, 19: 1α,25-dihydroxy-11-(3-hydroxy-1-propynyl)-9,11-didehydro-vitamin D3, 20: testolic acid, 21: caldorin, 22: lophachinin D, 23: 15(R)-PGF2α), alkaloids (i.e., 24: samandarine, 25: samandaridine, 26: samandarone, 27: samanine, 28: samanone, 29: samandenone, 30: samandinine, 31: cycloneosamandione, 32: isocycloneosamandarine, 33: O-acetyl-samandarine, 34: O-(S)-3-hydroxybutanoylsamandarine, 35: (1-methyl-2-azepanyl)[4-(4-morpholinylcarbonyl)-1,4′-bipiperidin-1′-yl]methanone, 36: anatoxin-a (ATX-a), 37: TTX, 38: 6-epi-TTX, 39: 4-epi-TTX, 40: 4,9-anhydro-TTX, 41: 4,9-anhydro-6-epi-TTX, 42: 11-deoxy-TTX), biogenic amines (i.e., 43: tryptamine, 44: serotonin, 45: l-DOPA, 46: alanine, 47: γ-aminobutyric acid, 48: aspartic acid, 49: glutamic acid, 50: histidine, 51: isoleucine, 52: leucin, 53: phenylalanine, 54: proline, 55: serine, 56: tryptophane, 57: tyrosine, 58: valine, 59: pyroglutamic acid), proteins, and peptides. GG, granular gland; MG, mucous gland; SG, stratum germinativum; SS, stratum spongiosum; SC, stratum compactum.](/document/doi/10.1515/hsz-2024-0035/asset/graphic/j_hsz-2024-0035_fig_001.jpg)
Chemical composition of caudate amphibian skin secretions. On the skin of caudates, several different classes of substances were identified, encompassing fatty acids and cholesterol derivatives (i.e., 1: Palmitoleic acid, 2: myristoleic acid, 3: linolenic acid, 4: cholesterol, 5: cholesteryl stearate, 6: cholesteryl oleate, 7: cholesteryl palmitate, 8: (2α,3β)-2,3-bis[(4-carboxy-3,3-dimethylbutanoyl) oxy]olean-12-en-28-oic acid, 9: dihydronandrolone, 10: (15Z)-9,12,13-trihydroxy-15-octadecenoic acid, 11: nebrosteroid L, 12: androstenedione, 13: 16-alpha-17-beta-estriol, 14: minabeolide-8, 15: taurochenodeoxycholic acid, 16: cholic acid, 17: palmitoyl glycine, 18: N-arachidonoyl serine, 19: 1α,25-dihydroxy-11-(3-hydroxy-1-propynyl)-9,11-didehydro-vitamin D3, 20: testolic acid, 21: caldorin, 22: lophachinin D, 23: 15(R)-PGF2α), alkaloids (i.e., 24: samandarine, 25: samandaridine, 26: samandarone, 27: samanine, 28: samanone, 29: samandenone, 30: samandinine, 31: cycloneosamandione, 32: isocycloneosamandarine, 33: O-acetyl-samandarine, 34: O-(S)-3-hydroxybutanoylsamandarine, 35: (1-methyl-2-azepanyl)[4-(4-morpholinylcarbonyl)-1,4′-bipiperidin-1′-yl]methanone, 36: anatoxin-a (ATX-a), 37: TTX, 38: 6-epi-TTX, 39: 4-epi-TTX, 40: 4,9-anhydro-TTX, 41: 4,9-anhydro-6-epi-TTX, 42: 11-deoxy-TTX), biogenic amines (i.e., 43: tryptamine, 44: serotonin, 45: l-DOPA, 46: alanine, 47: γ-aminobutyric acid, 48: aspartic acid, 49: glutamic acid, 50: histidine, 51: isoleucine, 52: leucin, 53: phenylalanine, 54: proline, 55: serine, 56: tryptophane, 57: tyrosine, 58: valine, 59: pyroglutamic acid), proteins, and peptides. GG, granular gland; MG, mucous gland; SG, stratum germinativum; SS, stratum spongiosum; SC, stratum compactum.
As such, 24 fatty acids, among these myristoleic, linolenic, and palmitoleic acid (1–3, Figure 1), were identified on the skin of P. cinereus (Rickrode et al. 1986). Apart from the classical role of skin lipids to retard water evaporation (Toledo and Jared 1993), Rickrode et al. (1986) observed an antibiotic function of myristoleic, linolenic, and palmitoleic acid. In addition, the steroid compound cholesterol (4; ∼0.27–0.65 % of dried secretions; Figure 1) was found in skin secretions of S. atra, S. salamandra, and T. carnifex (Croce and Bolognani 1975). Among cholesterol (4), cholesterol esters (i.e., cholesteryl palmitate, stearate, and oleate; 5–7, Figure 1) were found in the subspecies S. salamandra bernardezi (Habermehl 1965; under the name Salamandra maculosa taeniata). Furthermore, various steroids and cholesterol derivatives (i.e., (2α,3β)-2,3-bis[(4-carboxy-3,3-dimethylbutanoyl) oxy]olean-12-en-28-oic acid (8), dihydronandrolone (9), (15Z)-9,12,13-trihydroxy-15-octadecenoic acid (10), nebrosteroid L (11), androstenedione (12), 16-alpha-17-beta-estriol (13), minabeolide-8 (14), taurochenodeoxycholic acid (15), and cholic acid (16)), fatty acid-amino acid conjugates (i.e., palmitoyl glycine (17) and N-arachidonoyl serine (18)), secosteroids (i.e., 1α,25-dihydroxy-11-(3-hydroxy-1-propynyl)-9,11-didehydro-vitamin D3 (19) and testolic acid (20)), the polyketide caldorin (21), and the diterpene lophachinin D (22) were observed as components in skin mucus of S. infraimmaculata by applying a non-targeted metabolomics analysis (Figure 1; Degani et al. 2023a,b). Finally, four prostaglandins from F-series (e.g., 15(R)-PGF2α (23)) were identified as arachidonic acid-derived sex pheromones in skin secretions from male and ovulated female H. leechii (Eom et al. 2009).
2.2 Carbohydrates
Although for anuran amphibians it is known that the skin mucus contains glycosaminoglycans (e.g., chondroitin sulfates) that reduce skin water loss (Toledo and Jared 1995), these were not yet confirmed in caudate skin secretions. There are only reports on a low carbohydrate content (0.41 % of dried secretions) in the adhesive skin secretions from the plethodontid salamander Plethodon shermani (Von Byern et al. 2017), which was earlier described as the presence of neutral and acidic carbohydrates (Largen and Woodley 2008).
2.3 Alkaloids: samandarine-type toxins, TTX derivates, and others
In the course of toxic skin secretion characterization from S. salamandra, Zalesky (1866) isolated the first alkaloid ever from caudates, which he introduced as “samandarine” (24, Figure 1). This oxazolidine derivate shows with a LD50 of ∼1.5 mg/kg (subcutaneous injection into mice) an extremely high toxicity which can cause convulsions, hypertension, and hyperventilation, eventually leading to death (Becker 1986). Several following studies revealed the presence of 10 other related alkaloids (25–34, Figure 1) in salamanders’ toxic secretions, comprising besides samanine (27) and samanone (28) six oxazolidine (i.e., samandaridine (25), samandarone (26), samandenone (29), samandinine (30), O-acetylsamandarine (33), and O-(S)-3-hydroxy-butanoyl-samandarine (34)) and two carbinolamine (i.e., cycloneosamandione (31) and isocycloneosamandaridine (32)) derivatives (Barros et al. 2022; Knepper et al. 2019; Lüddecke et al. 2018). As such, samandarone (26) and O-acetylsamandarine (33) were presented as the actually pronounced alkaloids in S. salamandra, and samandarine (24) as the major alkaloid in skin secretions from the related species Salamandra atra and S. lanzai (Barros et al. 2022; Knepper et al. 2019; Lüddecke et al. 2018). Recently, the presence of samandarone (26), samanine (27), and cycloneosamandione (“samandridine”, 31) was also approved in skin secretions of an additional relative, S. infraimmaculata (Degani et al. 2023a). It has been demonstrated that all Salamandra species contain samandarine (24) and/or samandarone (26) in their secretions, which was confirmed for species of the sister group, Lyciasalamandra (Vences et al. 2014). In addition, molecules, like (1-Methyl-2-azepanyl)[4-(4-morpholinyl-carbonyl)-1,4′-bipiperidin-1′-yl]methanone (35), were identified in the skin secretions of S. infraimmaculata, whose functions have not been unraveled yet (Degani et al. 2023a). Interestingly, the bicyclic amine alkaloid anatoxin-a (36; ATX-a, “very fast death factor”), which is well-known as a cyanobacterial neurotoxin, has been recently found in the secretions of S. infraimmaculata as well (Degani et al. 2023a). As an agonist of the nicotinic acetylcholine receptor, it causes convulsions and eventually death, explaining its high toxicity with a LD50 of ∼100 μg/kg upon intravenous injection into mice (Fawell et al. 1999). From the series of Salamandra toxins, samandarone (26) was also detected in low concentrations in other species of the family Salamandridae, namely of the genera Calotriton, Euproctus, Lissotriton, and Triturus (Vences et al. 2014). In addition, various salamandrid newt species produce and/or sequester and secrete the neurotoxin TTX (37, Figure 1), which is a highly potent antagonist of voltage-gated sodium ion channels with the consequence of paralysis (LD50 ∼ 12.5–16 μg/kg upon subcutaneous administration into mice) (Abal et al. 2017; Kudo et al. 2020; Yotsu-Yamashita et al. 2007). In some individuals of Ichthyosaura alpestris, Triturus cristatus, Lissotriton helveticus, and Lissotriton vulgaris, small amounts of TTX (37; up to 9 μg/g), 6-epi-TTX (38; up to 17 μg/g), and other TTX derivatives (i.e., 4-epi-TTX (39), 4,9-anhydro-TTX (40), and 4,9-anhydro-6-epi-TTX (41)) were detected (Yotsu-Yamashita et al. 2007). In another study, the additional presence of 11-deoxy-TTX (42, Figure 1) was confirmed in I. alpestris and L. vulgaris (Yotsu et al. 1990). Plethodontid newts of the genus Taricha, such as Taricha granulosa, were even shown to be able to produce TTX on their own without the need of dietary precursors (Gall et al. 2022). In addition, they protect their eggs from predators through loading the eggs with the neurotoxin (Hanifin et al. 2003). Apart from TTX, also the derivatives 6-epi-TTX (38) and 11-deoxy-TTX (39) were identified in skin secretions from T. granulosa (Yotsu et al. 1990). The same alkaloids were also found in other genera of Salamandridae, such as in the genera Cynops (i.e., C. ensicauda, C. pyrrhogaster, and C. orientalis), Paramesotriton (i.e., P. hongkongensis, P. chinensis, P. deloustali, and P. guangxiensis), and Notophthalmus (i.e., N. viridescens). In addition, the same applies to Ambystoma tigrinum, a species from the family Ambystomatidae (Yotsu et al. 1990; Yotsu-Yamashita et al. 2017). Furthermore, in the body of C. ensicauda popei, the derivatives 2-epi-TTX, 8-epi-TTX, 8-epi-5,6,11-trideoxy-TTX, 4,9-anhydro-8-epi-5,6,11-trideoxy-TTX, 1-hydroxy-8-epi-5,6,11-trideoxy-TTX, and 1-hydroxy-4,4a-anhydro-8-epi-5,6,11-trideoxy-TTX were identified, revealing potential insights into the biosynthesis and/or degradation of TTX in newts (Kudo et al. 2012; Kudo and Yotsu-Yamashita 2019).
Finally, caudate alkaloids are mainly known from the family of true salamanders (Salamandridae) with a focus on skin secretions from adults, emphasizing the need for further research on species from other families as well as from different metamorphosis states. In case of S. salamandra terrestris, there is already evidence for the complete absence of alkaloids in its larvae, even when taken out from oviducts of gravid females (Mebs and Pogoda 2005). In addition, there are also already species known, like Plethodon shermani, which have no alkaloids in their secretions at all – not even as adults (Von Byern et al. 2017).
2.4 Biogenic amines
In species of the genus Salamandra (Salamandridae), indolealkylamines, such as tryptamine (43) and serotonin (44), were identified as components of the skin secretions (Lüddecke et al. 2018; Roseghini et al. 1989). Varying concentrations between the species were detected, ranging from ∼3 to 5 µg serotonin/tryptamine (S. atra) to 10 µg serotonin and 55 µg tryptamine (S. salamandra) per gram extracted skin (Roseghini et al. 1989). In addition, low levels of L-3,4-dihydroxyphenylalanine (l-DOPA; 45) were found in the adhesive secretions of the glue-producing salamanders Plethodon shermani, family Plethodontidae (∼0.12 % of dried secretions; Von Byern et al. 2017) and Hynobius dunni, family Hynnobiidae (Von Byern et al. 2021). In case of H. dunni, the secretion of l-DOPA was confirmed to occur via granular glands (Von Byern et al. 2021). Finally, several amino acids (i.e., alanine (46), γ-aminobutyric acid (47), aspartic acid (48), glutamic acid (49), histidine (50), isoleucine (51), leucin (52), phenylalanine (53), proline (54), serine (55), tryptophane (56), tyrosine (57), and valine (58)) were detected in the skin secretions of various newt species of the genera Cynops, Lissotriton, and Triturus (Bachmayer and Michl 1965; at that time all referred to the genus Triturus) and pyroglutamic acid (59) in those of Salamandra infraimmaculata (Degani et al. 2023a).
2.5 Proteins and peptides
Proteins were ranked as the highest portion (>70 %) in caudate amphibian skin secretions of some species, encompassing, for example, the newts Lissotriton vulgaris and Triturus ivanbureschi (Salamandridae) as well as the salamanders Plethodon shermani and Ambystoma macrodactylum (Plethodontidae and Ambystomatidae, respectively) (Hopkins and Migabo 2010; Karış et al. 2018; Von Byern et al. 2017). In L. vulgaris (18 protein fractions) and T. ivanbureschi (20 protein fractions), not further characterized proteins in the range of ∼9 kDa–∼272 kDa were found and correlated with a cytotoxic, antimicrobial, and hemolytic activity of the skin secretions (Karış et al. 2018). In L. vulgaris, T. cristatus as well as in the newt C. pyrrhogaster, four proteins were classified as hemolytically active enzymes, namely an amylase, a leucylnaphtylamide-cleaving enzyme, a phosphatase, and a phospholipase (Bachmayer and Michl 1965). Similar to the number of proteins in the secretions of Triturus species, in the salamander glue of P. shermani ∼18 proteins (10–160 kDa) were found and characterized by a high glycine amount as well as 52 % polar, 48 % non-polar, and 38 % hydrophobic amino acids (Von Byern et al. 2017). For Ambystoma species it is so far only known that the adhesiveness of the skin secretions is caused by the protein content without knowing the actual participating proteins (Hamning et al. 2000; Hopkins and Migabo 2010). In addition, disulfide bond crosslinking was suggested to play a crucial role for the consistency of the secretions (Hamning et al. 2000). In Salamandra salamandra, proteins with masses of 10–60 kDa were observed and characterized as fibrinogenolytically, hydrolytically (collagenolytically), and gelatinolytically active enzymes (Nikolaieva et al. 2018). Through their proteolytic activities these enzymes were suggested to cause the generation of further small bioactive molecules in the secretions (Nikolaieva et al. 2018). A higher variety of proteins (155 in total) was identified in the mucus of the Chinese giant salamander Andrias davidianus, family Cryptobranchidae (Geng et al. 2015). Among these proteins, mainly sticky fiber and glycoproteins were found by an MS-based proteomic analysis (Geng et al. 2015), highlighting proteins involved in extracellular matrix organization (e.g., collagens α-1(I, XI), collagen α-2(IX), collagenase 3, procollagen galactosyltransferase 1-A/B, and protein disulfide-isomerase), cell adhesion/junction (e.g., annexin-B11, apolipoprotein A-IV, galectins-2/7, talin-2, and xin actin-binding repeat-containing protein 2), cytoskeleton organization (e.g., keratin 9, beta-actin, talin-2, xin actin-binding repeat-containing protein 2, dystrophin-1, profilin), immune response (e.g., galectin-2, cGMP-dependent 3′,5′-cyclic phosphodiesterase, Ig κ chain V-II region WOL, apolipoprotein A-IV, MHC class I heavy chain maturation peptide H–2K (D), and MHC class I heavy chain), and wound healing (e.g., β-actin, cGMP-dependent 3′,5′-cyclic phosphodiesterase, osteonectin, profilin, heat shock cognate 71 kDa protein, and protein hook homolog 1) (Geng et al. 2015). In particular, Qu et al. (2014) isolated and characterized the ∼17 kDa protein “lectin” out of the mucus of the skin of A. davidianus. The protein was only partially sequenced and showed antibacterial activity in vitro (Qu et al. 2014). Furthermore, in another study the presence of lysozymes was confirmed in the skin of A. davidianus and the axolotl Ambystoma mexicanum (Yang et al. 2017a; Yu et al. 2013). Although the antibacterial activity of these lysozymes was already confirmed, it is still under investigation whether these lysozymes are also subject of secretion and might thus be involved in skin protection (Yang et al. 2017a; Yu et al. 2013). Secretomics, as extensively performed for anuran species (e.g., Ong et al. 2021; Thompson et al. 2007), are required for a complete identification of proteins from caudate amphibian skin secretions.
During mating season, male caudates also secrete proteins and/or peptides as courtship pheromones from specific glands which are then distributed through water to or actively displayed onto female’s skin (Woodley and Staub 2021). In contrast to mucus proteins, these protein and peptide pheromones are better characterized, as evident, for example, by the well-studied sodefrin precursor-like factor (SPF) system that consists of ∼20 kDa glycosylated proteins (Janssenswillen et al. 2014). SPF isoforms were found, for example, in species of the families Plethodontidae (e.g., Desmognathus ocoee and Karsenia koreana), Salamandridae (e.g., I. alpestris, L. helveticus, and N. viridescens), and Ambystomatidae (A. mexicanum) (DeBruin et al. 2023; Hall et al. 2016; Houck et al. 2008; Janssenswillen et al. 2015; Treer et al. 2017). Persuasins, ∼15 kDa proteins with a characteristic disulfide bond pattern, were found in the courtship elution profile of I. alpestris and L. helveticus, thereby exerting a synergistic effect together with the SPF pheromone system (Maex et al. 2018). Beyond the SPF protein pheromones, the 22 kDa cytokine-homolog plethodon receptivity factor (PRF) has been isolated from e.g., P. shermani, and was shown to increase female receptivity in most plethodontid salamanders but exclusively in the genus Plethodon (Rollmann et al. 1999). An opposite effect to PRF was described to be induced by the 7 kDa protein “plethodontid modulating factor” (PMF), which reduces female receptivity. However, in combined presence with PRF an increase of female receptivity was observed (Houck et al. 2007).
Finally, already some peptide pheromones and bioactive peptides were isolated from the skin of urodeles, whose extraction, chemistry, and bioactivities are described in detail in the following sections.
3 Strategies for peptide extraction and isolation from caudate amphibian skin
Exocrine cutaneous glands produce a variety of substances in the gland alveolus by secreting epithelium. The secretions are then transported through the duct of the glands and released onto the skin. However, granular glands are surrounded by a contractile myoepithelial layer that supports active secretion of the granular components (Çömden et al. 2023). As such, several techniques for stimulation of the skin secretion and collection of skin peptides were established for anurans, but for caudates only poorly developed. In the following, the techniques from already successful investigations on caudate amphibian skin peptides are presented, including the stimulation, extraction, and isolation of peptides (Figure 2).
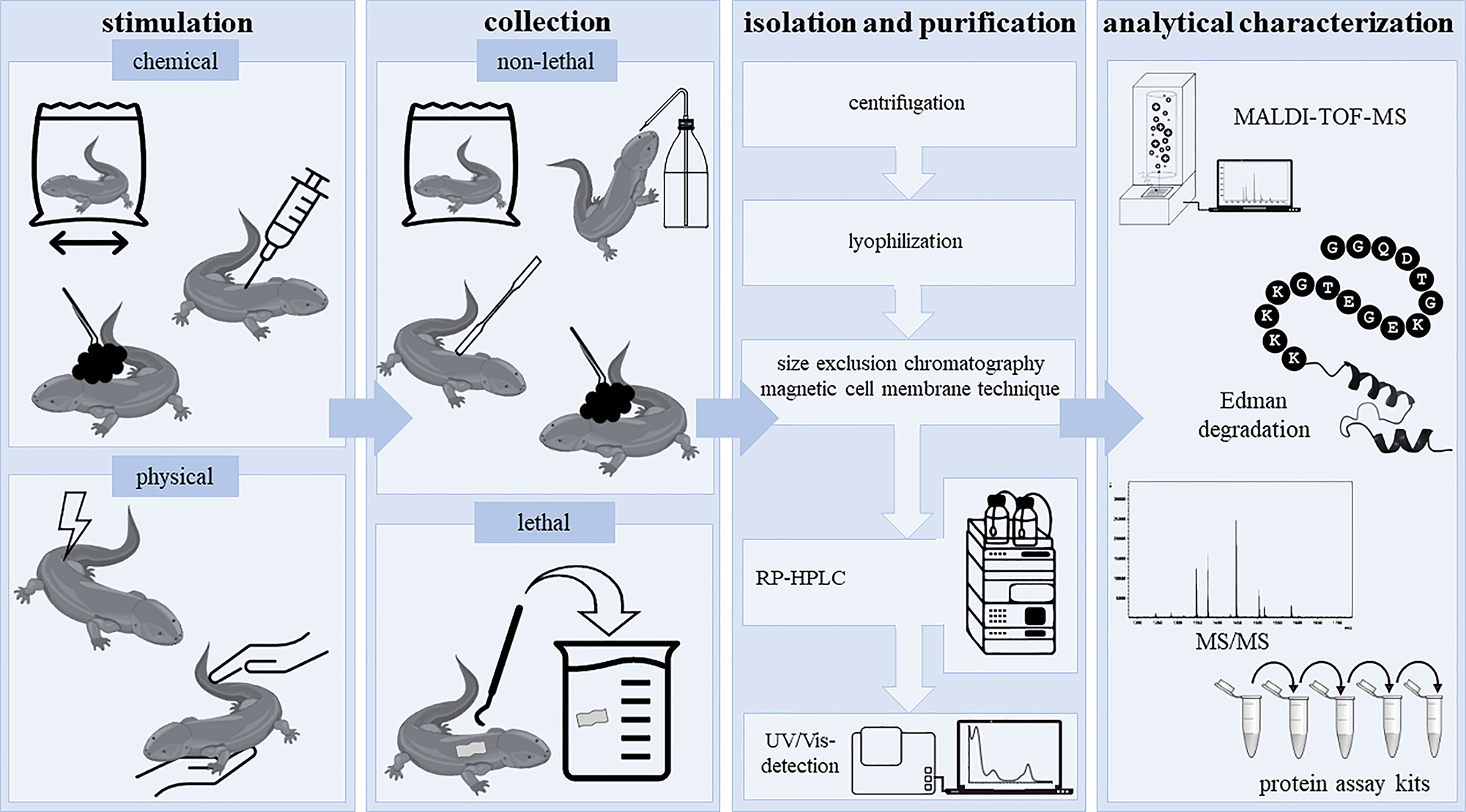
Stimulation, collection, extraction, and analytical characterization of caudate amphibian secretions. The so far reported collection protocols for secretions from caudate amphibians are summarized. After chemical or physical stimulation, the secretions can be collected from living caudates (e.g., by rinsing and swabbing). When using the lethal collection method skin is cut off followed by a homogenization step. Homogenates and/or secretions are then centrifuged, lyophilized, separated and analytically characterized (e.g., by MALDI-TOF-MS and Edman sequencing).
3.1 Stimulation of secretion and collection of skin peptides
Traditionally, the extraction of skin secretions was performed from excised skins of dead caudates without any need for prior secretion stimulation (Fredericks and Dankert 2000; Teranishi et al. 2004; Yang et al. 2017b). In part, this method is still used today to isolate skin peptides from caudates. As such, researchers anaesthetized individuals of the salamander Tylototriton verrucosus (Salamandridae) with isofluorane (2.5 %) and cut off the gland-rich dorsal skin (Jiang et al. 2015; Mu et al. 2014). Subsequently, the skin was homogenized in slightly acidic phosphate-buffered saline (PBS; including a protease inhibitor mixture; pH 6.0), centrifuged for 10 min, and lyophilized (Jiang et al. 2015; Mu et al. 2014). Another method was presented by Luo et al. (2021), who anaesthetized Tylototriton kweichowensis with ice and destroyed brain and spinal marrow with a needle for euthanasia. The dorsal skin was then dissected and stored in liquid nitrogen prior to further use (Luo et al. 2021). Similarly, Fredericks and Dankert (2000) homogenized skin from the plethodontid species Plethodon cinereus in sodium acetate buffer (50 mM, including a protease inhibitor mixture), centrifuged the homogenate for 20 min, and stored it at −70°C. Today, strategies for the stimulation of active secretion from amphibian cutaneous glands of living amphibians are preferred and encompass chemical as well as physical techniques (Figure 2) (Pereira et al. 2021; Xiao et al. 2011). Thereby, chemical stimulation is conducted either by injection or immersion (Pereira et al. 2021; Xiao et al. 2011). In case of injection, typically neurotransmitters (e.g., acetylcholine, adrenaline, noradrenaline) are applied into the lymphatic system of anaesthetized amphibians (Xiao et al. 2011). Inspired by a protocol used for the stimulation of peptide release in anurans (Rollins-Smith et al. 2002), Sheafor et al. (2008) injected noradrenaline into larvae and adults of the salamander A. tigrinum and subsequently transferred the animals into buffer solutions (25 mM sodium chloride, 25 mM ammonium acetate; pH 7.0) for 15 min to allow for extraction of the skin peptides. Although subcutaneous acetylcholine injection was shown to effectively lead as well to granular gland discharge, for example, in T. cristatus, this procedure is rarely used for caudates, since their lymphatic system is not that easily accessible as in anurans and requires several skin injury provoking injections (Hedrick et al. 2013; Hoffman and Dent 1977; Pereira et al. 2021). In contrast to the injection-based stimulation, the immersion strategy (without anesthesia) is more commonly used. In a recent study of Pereira et al. (2021), salamanders (N. viridescens and Desmognathus ochrophaeus) were individually immersed in a plastic bag containing collection buffer (50 mM sodium chloride, 25 mM sodium acetate; no pH given) with neurotransmitters [i.e., noradrenaline (1 mM) or acetylcholine (1 mM)] and carefully moved back and forth for 10 min (Pereira et al. 2021). Another chemical approach is direct swabbing of the animals with cotton containing chemicals, such as ether, and subsequent rinsing or scrapping the skin to collect the released substances (Xiao et al. 2011). However, this has not been applied yet to collect peptides from caudate amphibian skin. In fact, swabbing without any chemical stimulation is a classical method of choice for the analysis of mucus (Degani et al. 2023a,b; Umile et al. 2014). As such, Degani et al. (2023a,b) used, for example, filter paper to collect the mucous secretions of Salamandra infraimmaculata, which was afterwards used for further extraction of the substances. Prior to further usage, the swabbing material is then transferred into, for example, methanol, vortexed for 10 min, and finally filtered through a 0.22 µm membrane (Degani et al. 2023a,b). Another non-invasive method displays immersion of individuals in a plastic bag containing buffer as well as continuously and firmly massaging the animal for 10 min, as has been performed for several salamanders (Ambystoma maculatum, Amphiuma tridactylum, D. ochrophaeus, Desmognathus fuscus, Eurycea bislineata, E. longicauda, Necturus beyeri, Necturus maculosus, N. viridescens, and Siren intermedia) (Pereira et al. 2021, 2023). As a result of the comparative study of Pereira et al. (2021), the authors described that this kind of physical stimulation seems to be more effective for the stimulation of peptide release from caudate amphibian skin than immersion in neurotransmitter solution (Pereira et al. 2021). Since neither noradrenaline nor acetylcholine showed any support with respect to peptide recovery from the salamanders’ skin, the use of both is even critically discussed with respect to a potential effect on skin peptide quantification (Pereira et al. 2021).
Finally, the most common and appropriate technique for the stimulation of secretions with the aim of peptide isolation is the electrical stimulation, which is recommended especially for species with compact glands (Pereira et al. 2021; Xiao et al. 2011). As such, Meng et al. (2013) treated, for example, the salamander Cynops fudingensis for 3–5 s with an alternating electric current of 3 V, followed by rinsing the animals with buffer (phosphate-buffered saline including a protease inhibitor mixture) to obtain the secretions. With this method, Meng et al. (2013) were the first one identifying and characterizing an antimicrobial peptide in salamander skin secretions. For the stimulation of skin secretion in Salamandra salamandra, an even higher electrical stimulation of 9 V was applied for a few seconds and secretions were collected in water (Plácido et al. 2020).
In future, a systematic analysis of stimulation and collection methods is required to characterize the most appropriate method for caudate amphibian skin secretions, and thus allow for a complete mapping of skin peptides and their physiological function.
3.2 Isolation and purification of caudate amphibian skin peptides
Commonly, secretions and homogenates are centrifuged for at least 10 min, then lyophilized and finally transferred into the freezer (Figure 2) (Degani et al. 2023a,b; Fredericks and Dankert 2000; Meng et al. 2013; Pereira et al. 2021; Plácido et al. 2020). Lyophilized extracts are then, for example, dissolved in buffers (e.g., PBS, pH 6.0) and separated by size exclusion chromatography, followed by purification via reverse-phase high-performance liquid chromatography (RP-HPLC) equipped with a UV/Vis detector (Jiang et al. 2015; Meng et al. 2013; Mu et al. 2014; Pereira et al. 2023; Plácido et al. 2020; Teranishi et al. 2004). As usual for peptides, water and acetonitrile with 0.1 % trifluoroacetic acid (TFA) as a modifier are used as eluents (Meng et al. 2013; Mu et al. 2014).
Alternatively, Pei and Jiang (2017) recently reported a magnetic cell membrane separation technique using magnetic nanoliposomes as a purification step of lyophilized mucus before RP-HPLC separation.
3.3 Qualitative and quantitative assessment of caudate amphibian skin peptides
So far, peptides from caudate amphibian skin secretions were in part characterized by matrix-assisted laser desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) and (automatic) Edman sequencing or MS/MS for analysis of the amino acid sequences (Figure 2) (Meng et al. 2013; Mu et al. 2014; Pei and Jiang 2017; Plácido et al. 2020; Teranishi et al. 2004). In a few studies, the sequence and molecular masses of the peptides were – so far – not further characterized after HPLC analysis of the secretions and were just tested for their bioactivity as part of the collected sample or as a separated fraction (e.g., Fredericks and Dankert 2000; Pereira et al. 2023).
Quantification of components, such as peptides, from mucous or granular secretions is still difficult to perform since many parameters, such as rate of production and discharge, are not known. For initial quantification, the peptide concentrations in caudate amphibian skin secretions were (so far) determined by the application of protein assay kits, such as the BCA protein assay kit (Pereira et al. 2021, 2023; Sheafor et al. 2008). A peptide amount of ∼325–400 μg/g (body weight) was for example found on the skin of A. tigrinum larvae (Ambystomatidae) (Sheafor et al. 2008). Interestingly, in the past, no difference of the peptide amount in skin secretions of captive and free-living salamanders was observed (Pereira and Woodley 2021).
4 Chemical features of skin peptides from caudates
In total, 14 peptides (P1–P14) from three caudate families were so far described in detail, either as components of granular and mucous skin secretions (i.e., P1–P7, P9, P13, P14) or as pheromones [i.e., silefrin (P8), sodefrin (P10), aonirin (P11), and imorin (P12)] (Table 1). In the following, the chemical characteristics, such as length, sequence, amino acid composition, charge, and secondary structure of the so far identified caudate skin peptides are presented (Figure 3A–D).
Overview of the chemical features and the bioactivity of caudate amphibian skin peptides.
| No. | Name | Sequence | Species | Net charge at pH 7.0 | IEP | Structural features | Bioactivity |
|---|---|---|---|---|---|---|---|
| Plethodontidae | |||||||
|
|
|||||||
| P1 | Ramosin | H-GWFSLVRKVVGGVGSL-NH2 | Bolitoglossa ramosi | +3 | 11.0 | α-helix* | AM |
|
|
|||||||
| Salamandridae | |||||||
|
|
|||||||
| P2 | Infraim-I | H-fLX-NH-CH2CH3 | Salamandra infraimmaculata | +1 | n.d. | Random coil | n.d. |
| P3 | Antillatoxin B | cyclo-[A(homoF)-(-N-methyl)-G-(1-oxa-(2-(1E,3E)-1-methyl-3-methyl-5,5-diemthyl-1,3-hexadienyl)-3-methyl-4-methylidene-hexan-6-on)] | S. infraimmaculata | 0 | n.d. | Cyclic | AM, NT |
| P4 | Salamandrin-I | H-FAVWGCADYRGY-NH2 | S. salamandra | +1 | 8.8 | α-helix, random coil* | AO, CT, MT |
| P5 | Tk-Cath | H-GGQDTGKEGETGKKKKSDNWFMNLLNKFLELIGLKEAGDDSEPFCFTCIFDMFSQ-OH | Tylototriton kweichowensis | −3 | 4.7 | α-helix, random coil | HL, AO, AI, WH |
| P6 | CCK-TV | H-DYMGWMDF-NH2 | T. verrucosus | −1 | 3.8 | Random coil | MT |
| P7 | Tylotoin | H-KCVRQNNKRVCK-OH (1× –S–S–) | T. verrucosus | +5 | 10.3 | β-sheet | WH |
| P8 | Silefrin | H-SILSKDAQLK-OH | Cynops ensicauda | +1 | 8.3 | Random coil | PM |
| P9 | CFBD-1 | H-FAVWGCADYRGYCRAACFAFEYSLGPKGCTEGYVCCVPNTF-OH (3× –S–S–) | C. fundigensis | 0 | 6.1 | β-sheet, random coil | AM, HL |
| P10 | Sodefrin | H-SIPSKDALLK-OH | C. pyrrhogaster | +1 | 8.3 | α-helix, random coil | PM |
| P11 | Aonirin | H-SIPSKDAVLK-OH | C. pyrrhogaster | +1 | 9.9 | Random coil | PM |
| P12 | Imorin | H-AEF-OH | C. pyrrhogaster | −1 | 3.3 | Random coil | PM |
|
|
|||||||
| Cryptobranchidae | |||||||
|
|
|||||||
| P13 | Ad-Cath | H-RPKKVQGRKAEKDNGDGTTAANASGKKKSSNVFK-OH | Andrias davidianus | +7 | 10.5 | Random coil | AM, CT |
| P14 | Andricin-I | H-AIGHCLGATL-OH | A. davidianus | 0 | 6.8 | α-helix, random coil* | AM |
-
Caudate amphibian skin peptides P1 to P14 are listed including their amino acid sequence, modifications such as disulfide bonds (bold) and amidated C-temini (–NH2), species origin, net charge at pH 7.0, isoelectric point (IEP), structural features, and bioactivities. The information concerning structural features are derived from predictions by AlphaFold and experimental results (if available; “*”). IEP, isoelectric point; n.d., not determined; X, l-norvaline; AM, antimicrobial; CT, cytotoxic; NT, neurotoxic; HL, hemolytic; AO, antioxidant; AI, antiinflammatory; WH, wound-healing; PM, pheromonal; MT, myotropic.
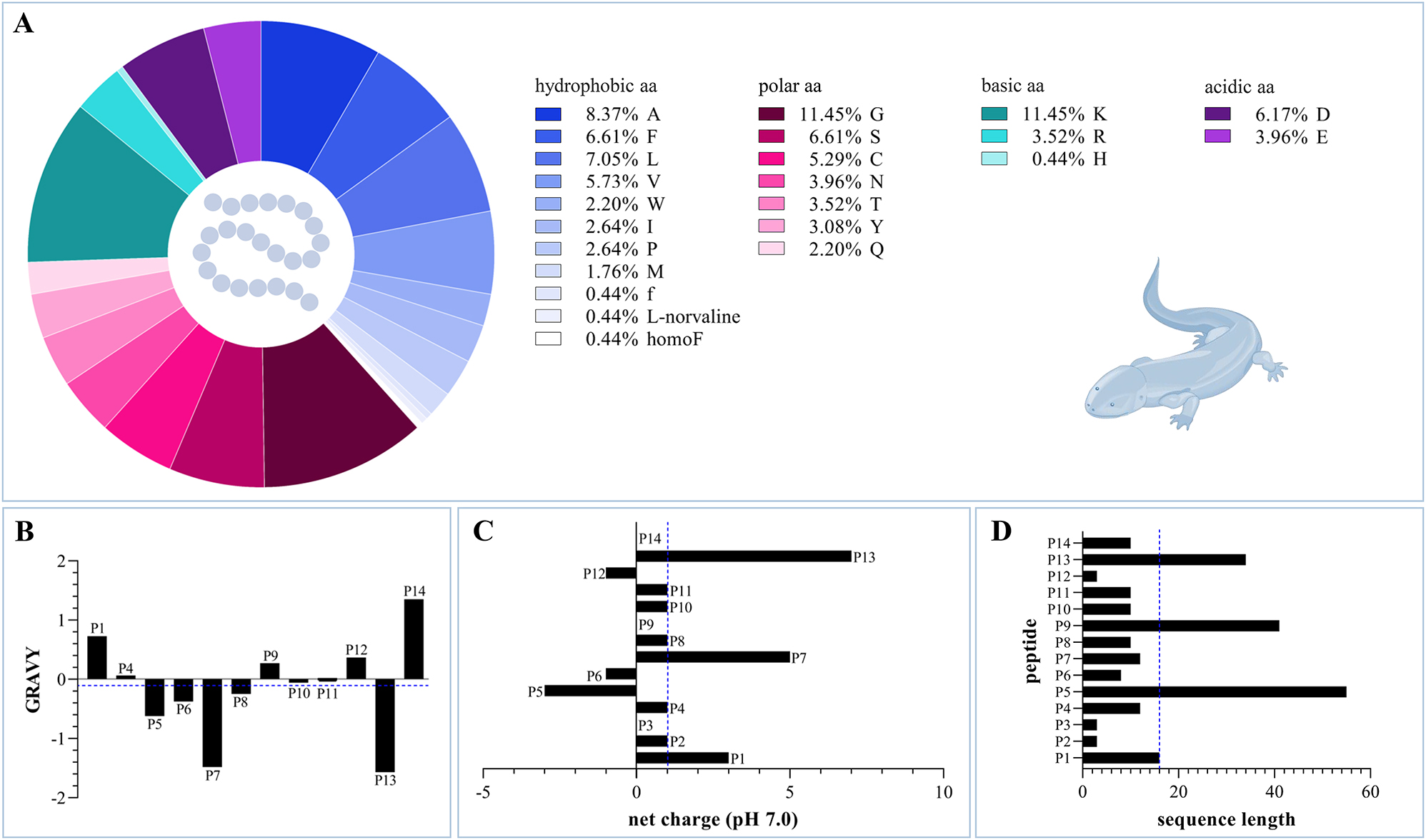
Chemical properties of caudate skin peptides P1–P14. (A) The amino acid composition of the 14 caudate amphibian skin peptides is shown, consisting of 23 amino acids. (B) The GRAVY values are displayed, with exception of the highly modified peptides P2 and P3. The average value is −0.102, which slightly tends towards predominantly hydrophilic properties. (C) The net charges at pH 7.0 of all the fourteen peptides P1 to P14 are given with an average value of +1. (D) The sequence length of the peptides ranges from three to 55 amino acids with an average of 16 amino acids. All average values are marked with a blue dashed line. aa, amino acid; GRAVY, grand average of hydropathicity.
4.1 Sequence length
With an average length of 16 amino acids, the sequence length of the caudate skin peptides varies so far between three and 55 amino acids (Table 1; Figure 3D), which is in the same range as found for anuran skin peptides (Xu and Lai 2015). Among these, the shortest peptide is the pheromone “imorin” (P12) from C. pyrrhogaster (Salamandridae), only consisting of three amino acid residues (Nakada et al. 2017), followed by two peptides from mucus of Salamandra infraimmaculata. Both of them, d-phenylalanyl-l-leucyl-N-ethyl-l-norvalinamide (P2; in the following designated as infraim-I) and the cyclic antillatoxin B (P3), display strongly modified peptides built out of three amino acids (Degani et al. 2023a,b). The peptide next in size, with eight residues, is CCK-TV from T. verrucosus (Salamandridae), and was assigned to the family of cholecystokinins (Jiang et al. 2015). Sodefrin (P10) and aonirin (P11), 10 residue long peptides, were found in C. pyrrhogaster, which display – as the above-mentioned imorin – peptide pheromones in the newt (Kikuyama et al. 1995). A similar long peptide, “andricin-I” (P14; formerly known as “andricin 01”), has been identified in A. davidianus of the family Cryptobranchidae (Pei and Jiang 2017). The newt, C. ensicauda, releases also a peptide pheromone with a length of 10 amino acids, called “silefrin” (P8) (Yamamoto et al. 2000), followed by two peptides from two different species producing 12 amino acids long skin peptides. As such, the peptide tylotoin (P7) was isolated from the skin of the Himalayan salamander (Mu et al. 2014), and salamandrin-I (P4) from secretions of the fire salamander S. salamandra (Plácido et al. 2020). Bolitoglossa ramosi, family Plethodontidae, releases ramosin (P1), which is 16 residues long (Medina et al. 2022). In contrast, with some distance, peptides found on the skin of A. davidianus, Cynops fundigensis, and T. kweichowensis display the largest peptides with amino acid sequences of 34–55 residues, comprising Ad-Cath (P13; 34 amino acids), CFBD-1 (P9; 41 amino acids), and Tk-Cath (P5; 55 amino acids) (Luo et al. 2021; Meng et al. 2013; Yang et al. 2017b). From these, Ad-Cath from A. davidianus and Tk-Cath from T. kweichowensis belong to the group of cathelicidins (Luo et al. 2021; Yang et al. 2017b).
4.2 Amino acid composition
Looking at the entirety of skin peptides that were isolated from caudates so far, they are consisting of 23 different amino acids (Figure 3A), including apart from the 20 proteinogenic amino acids also three non-proteinogenic amino acids (i.e., d-phenylalanine, l-homophenylalanine, and l-norvaline). Among them, the sequences contain ∼38 % hydrophobic amino acids (i.e., 8.37 % l-alanine, 5.73 % l-valine, 1.76 % l-methionine, 7.05 % l-leucine, 2.63 % l-isoleucine, 2.63 % l-proline, 2.20 % l-tryptophan, 6.61 % l-phenylalanine, 0.44 % d-phenylalanine, 0.44 % l-homophenylalanine, and 0.44 % l-norvaline). Polar amino acids (i.e., 3.01 % l-tyrosine, 3.52 % l-threonine, 2.20 % l-glutamine, 11.45 % l-glycine, 6.61 % l-serine, 5.29 % l-cysteine, and 3.97 % l-asparagine) are represented with a total percentage of ∼36 %. Amino acids with basic side chains occur with a percentage of ∼15 %, made up by 11.45 % l-lysine, 3.52 % l-arginine, and 0.44 % l-histidine. Finally, acidic amino acids (i.e., 3.97 % l-glutamic acid and 6.17 % l-aspartic acid) are less represented by in total ∼10 %. Thus, in the so far identified peptides, the most common amino acids are the small l-glycine, the basic l-lysine, and the hydrophobic l-alanine.
4.3 Hydropathicity, net charge, and isoelectric points
Calculation of the grand average of hydropathicity (GRAVY; Gasteiger et al. 2005) revealed andricin-I (P14; 1.35) as the most hydrophobic peptide and Ad-Cath (P13; −1.57) as the most hydrophilic peptide among the caudate skin peptides (Figure 3B). Ramosin (P1; 0.73), CFDB-1 (P9; 0.27), and salamandrin-I (P4; 0.06) belong to the peptides with more hydrophobic characteristics as well, whereas tylotoin (P7; −1.48), silefrin (P8; −0.25), sodefrin (P10; −0.06), aonirin (P11; −0.02), Tk-Cath (P5; −0.62), and CCK-TV (P6; −0.38) are more hydrophilic. With a GRAVY of 0, the small peptide imorin (P12) possesses balancing hydrophobic and hydrophilic residues. The two highly modified peptides from Salamandra infraimmaculata, antillatoxin B (P3) and infraim-I (P2), are both hydrophobic peptides. Antillatoxin B even belongs to the class of lipopeptides, actually well known as a toxin from marine cyanobacteria (Nogle et al. 2001).
The net charges of the peptides at pH 7.0 range from −3 to +7 (Table 1; Figure 3C). Negatively charged peptides are Tk-Cath (P5; net charge of −3), CCK-TV and imorin (P6 and P12; both with a net charge of −1). CFBD-1 (P9), antillatoxin B (P3), and andricin-I (P14) possess no net charge at pH 7.0, and the peptides salamandrin-I, silefrin, sodefrin, aonirin, infraim-I (P2, P4, P8, P10, and P11; all five with a net charge of +1), ramosin (P1; net charge of +3), tylotoin (P7; net charge of +5), and Ad-Cath (P13; net charge of +7) belong to the cationic peptides and display to the current state the bigger part of the caudate skin peptides.
Finally, the isoelectric points of these peptides show high variety in the range of 3.3 (imorin; P12) – 11.0 (ramosin; P1) (Table 1).
4.4 Sequence alignment
As already emphasized by others, salamandrin-I (P4) shares 100 % identity with the defensin CFBD-1 (P9). However, truncation of the defensin seems to be unlikely as the origin of salamandrin-I due to its C-terminal amide modification (Plácido et al. 2020). In addition, the peptide pheromones silefrin (P8), sodefrin (P10), and aonirin (P11; also known as “[Val8]-sodefrin”) differ only by one to two amino acids (80–90 % identity). Otherwise, sequence alignment of the already known caudate skin peptides demonstrates that the data amount is too small. Even an intra-familiar sequence alignment could not lead to the identification of further interspecies similarities of the peptides, highlighting the need for further research on caudate skin peptides in future.
4.5 Modifications
Three of the caudate skin peptides [i.e., ramosin (P1), salamandrin-I (P4), and CCK-TV (P6)] are modified by an amidated C-terminus (Table 1) (Jiang et al. 2015; Medina et al. 2022; Plácido et al. 2020). Besides the masking of charge, the amidation usually increases the stability due to the protection from proteolytic cleavage (Kim and Seong 2001). Tylotoin (P7), Tk-Cath (P5), and CFBD-1 (P9) may form intramolecular disulfide bonds (Luo et al. 2021; Meng et al. 2013; Mu et al. 2014). Tylotoin has one proven disulfide bond, Tk-Cath could potentially form one disulfide bond as well but this has not been proven so far. CFBD-1 belongs to the class of β-defensins, which are well known for their typical disulfide bond patterns (Luo et al. 2021; Mu et al. 2014). As such, it was suggested that CFBD-1 forms three intramolecular disulfide bonds (Meng et al. 2013).
As already stated before (see Section 4.2), the two mucus peptides (P2 and P3) from Salamandra infraimmaculata contain amino acid derivatives and non-proteinogenic amino acids, which also supports biological stability of these peptides (Degani et al. 2023a,b) (Figure 3). It is worth highlighting that antillatoxin B is a cyclic lipopeptide with hydrophobic properties, usually accounting for the ability of penetrating cell membranes (Degani et al. 2023b; Dougherty et al. 2019).
4.6 Secondary structure
The only caudate skin peptides that were also structurally characterized are ramosin (P1), andricin-I (P14) and salamandrin-I (P4) (Medina et al. 2022; Pei and Jiang 2017; Plácido et al. 2020). Circular dichroism (CD) spectroscopy was used to analyze the secondary structural elements of these peptides. For andricin-I and salamandrin-I, minima at ∼195 nm and a flat maximum above 210 nm indicated a random coiled structure (Pei and Jiang 2017; Plácido et al. 2020). While a 3D structure of andricin-I was calculated and modelled from the CD data, the 3D structure of salamandrin-I was predicted using the in silico tool PEP-FOLD (Pei and Jiang 2017; Plácido et al. 2020; Shen et al. 2014). Due to its relation to cathelicidins, Ad-Cath was speculated to possess an α-helical structure but structural analysis was not yet performed (Yang et al. 2017b). In contrast to these two peptides, CD spectroscopic analysis of the amphipathic ramosin allowed for the assumption of an α-helical structure (Medina et al. 2022), which was further confirmed by structural prediction through I-Tasser (Yang and Zhang 2015).
Since none of the caudate skin peptides’ 3D structures were experimentally determined so far, the AlphaFold tool in UCSF ChimeraX (Jumper et al. 2021; Meng et al. 2023) was herein applied to predict them and, thus, to get a first comparative impression of their overall structural features and similarities (Figure 4). In fact, the peptides can be sorted into four classes: infraim-I (P2), CCK-TV (P6), silefrin (P8), aonirin (P11), imorin (P12), and Ad-Cath (P13) show only a random coiled structure, while ramosin (P1), salamandrin-I (P4), and andricin-I (P14) comprise α-helical elements (Figure 4). In particular, the α-helical structure was correlated with an antimicrobial function in case of, for example, many anuran skin peptides, and should thus be correlated with observed bioactivities (see Section 5). Structures with combined random coiled and α-helical elements were additionally found for Tk-Cath (P5), and sodefrin (P10). The only peptides forming β-strand elements are tylotoin (P7), which is further stabilized by a disulfide bond, and CFBD-1 (P9) (Figure 4). Finally, antillatoxin B differs from the other peptides through its cyclic structure and the long aliphatic side chain (P3; Figure 4).
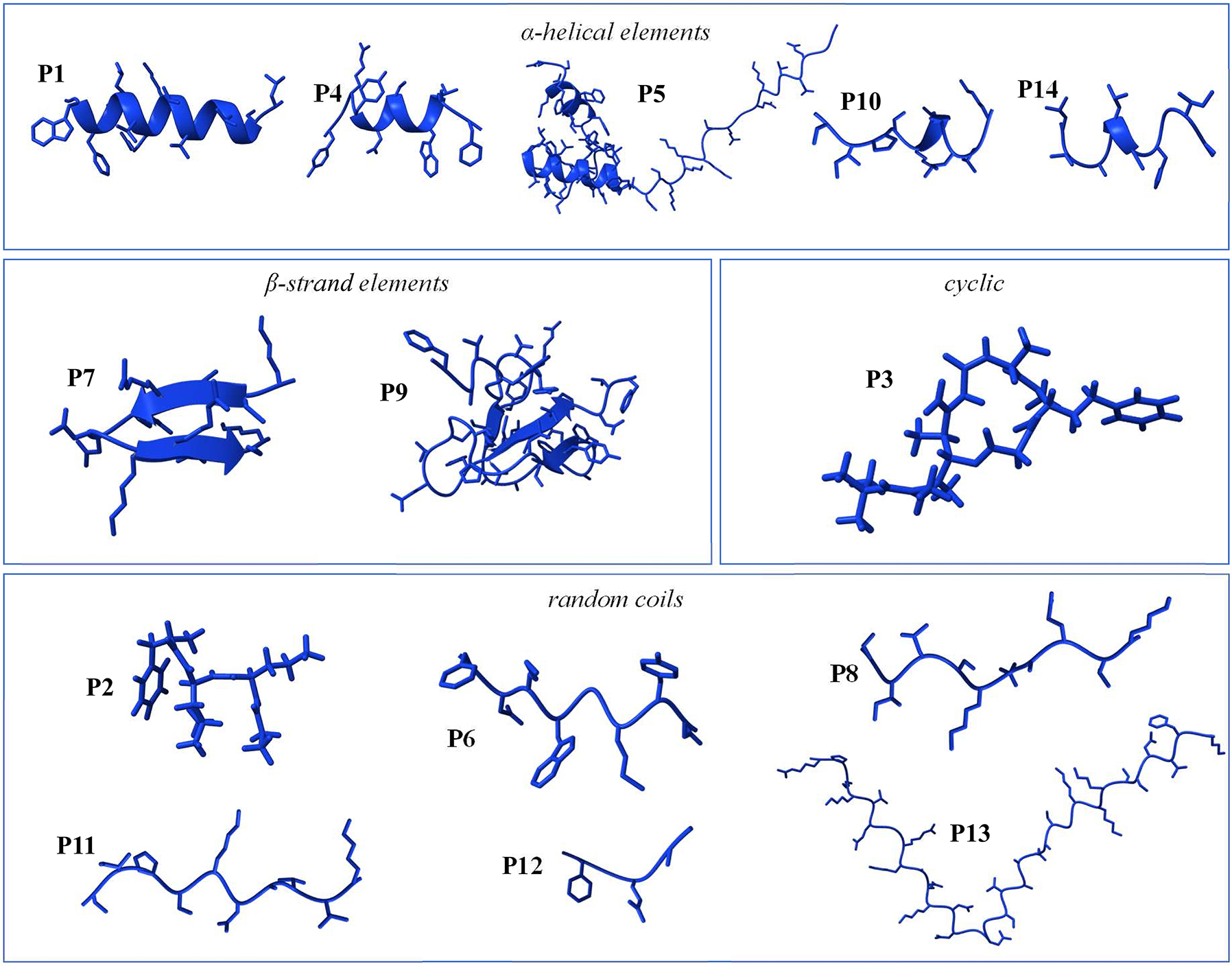
Predicted secondary structures of caudate amphibian skin peptides. As predicted by the AlphaFold tool in UCSF ChimeraX (Jumper et al. 2021; Meng et al. 2023), the secondary structures of skin peptides P1 to P14 are displayed and can be categorized by the presence of specific structural elements, involving α-helical elements (P1, P4, P5, P10, and P14), β-strand elements (P7 and P9), cyclic (P3), and random coils (P2, P6, P8, P11, P12, and P13).
4.7 Other peptides
Apart from the listed peptides (Table 1), there are also a few studies on peptides that were identified in several species, in some cases even purified by HPLC, and finally tested for their bioactivity but not further characterized with respect to, for example, their amino acid sequence and length (Figure 5). As such, peptides (undefined number) were found in the secretions of: Ambystomatidae: A. maculatum, A. tigrinum; Amphiumidae: A. tridactylum; Cryptobranchidae: Cryptobranchus alleganiensis; Plethodontidae: D. fuscus, D. ochrophaeus, D. ocoee, E. bislineata, E. longicauda, Plethodon cinereus, P. shermani; Proteidae: N. beyeri, N. maculosus; Plethodon cinereus, P. shermani; Salamandridae: T. cristatus; Sirenidae: S. intermedia (Bachmayer and Michl 1965; Fredericks and Dankert 2000; Pereira et al. 2018, 2023; Sheafor et al. 2008). Partially, a bioactivity could be already correlated with single peptide fractions (see Section 5; Fredericks and Dankert 2000; Pereira et al. 2018, 2023; Sheafor et al. 2008).
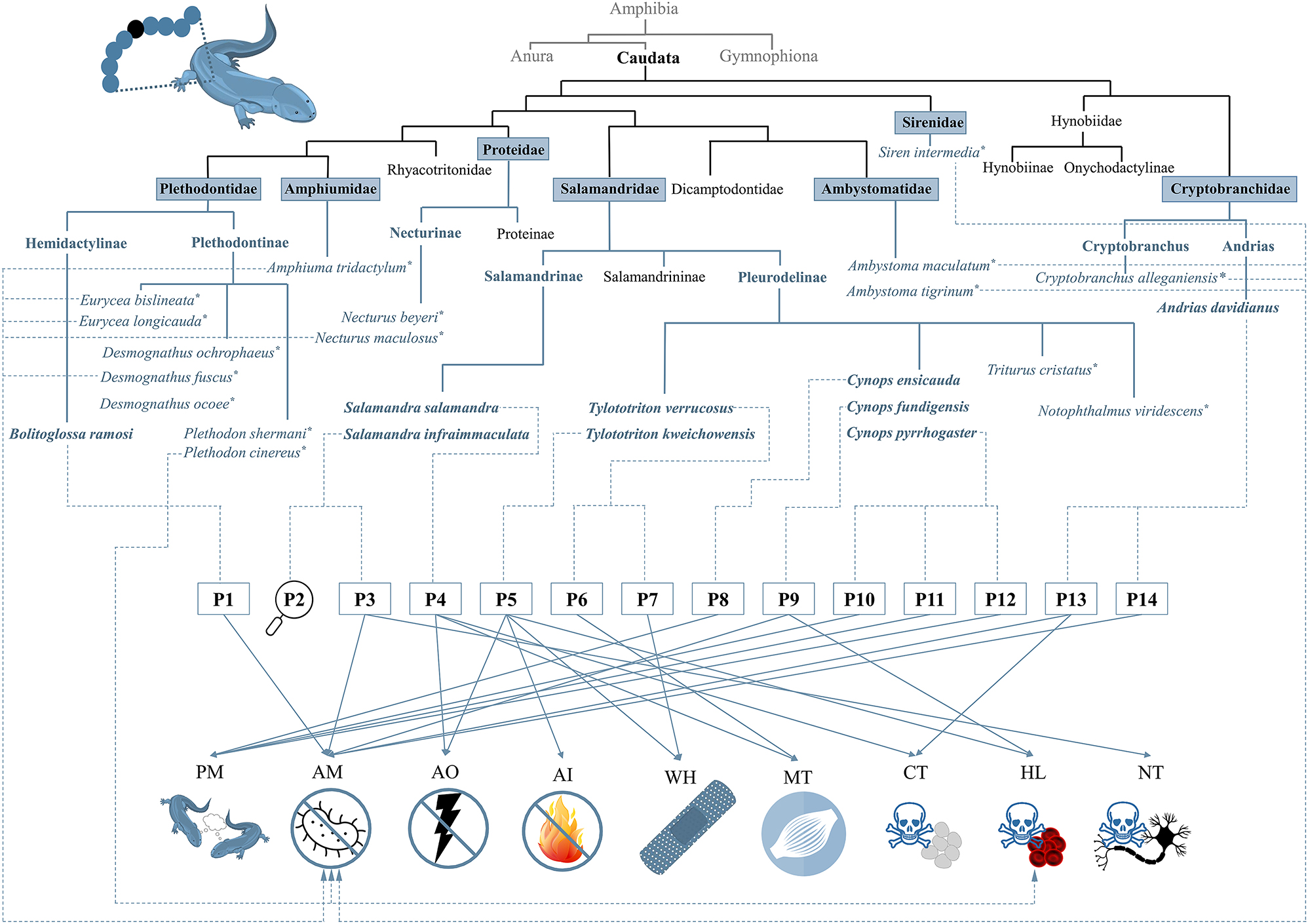
Overview of caudates with confirmed skin peptides and their so far shown bioactivities. According to the current state of knowledge, the species that were found to have peptides on their skin belong to the seven families Ambystomatidae, Amphiumidae, Cryptobranchidae, Plethodontidae, Proteidae, Salamandridae, and Sirenidae. Species, in which only the presence of peptides was confirmed but without further, detailed analysis are highlighted with “*”. Four skin peptides are amphibian pheromones (i.e., P8, P10, P11, and P12), two peptides (i.e., P1 and P14) show exclusively an antimicrobial activity, and one peptide acts only myotropically (P6). Antillatoxin B (P3) is an antimicrobial and neurotoxic peptide, while CFBD-1 (P9) shows antimicrobial and hemolytic activity, and Ad-Cath (P13) is antimicrobial and cytotoxic. Tylotoin (P7) shows its wound-healing bioactivity. The other wound-healing peptide, Tk-Cath (P5), shows in addition hemolytic, antioxidant, and antiinflammatory activities. For salamandrin-I (P4), an antioxidant, cytotoxic and myotropic bioactivity is known. The bioactivity of infraim-I (P2) has not yet been elucidated. AM, antimicrobial; CT, cytotoxic; NT, neurotoxic; HL, hemolytic; AO, antioxidant; AI, antiinflammatory; WH, wound-healing; PM, pheromonal; MT, myotropic.
5 Bioactivity of skin peptides from caudates
For the so far known caudate skin peptides a variety of bioactivities was analyzed and reported, including antimicrobial, antioxidant, antiinflammatory, wound-healing, and pheromonal, but also cytotoxic, hemolytic, and neurotoxic properties (Table 1 and Figure 5).
5.1 Antimicrobial activity
Antimicrobial peptides (AMPs) are often characterized by an α-helical structure with cationic and amphiphilic features. This supports channel-like pore formation within microbial lipid membranes eventually causing cell death (Huan et al. 2020). From the already reported caudate skin peptides, five peptides (i.e., P1, P3, P9, P13, and P14) were described to exert antimicrobial effects against Gram-negative and Gram-positive bacteria (Dussault et al. 2016; Medina et al. 2022; Meng et al. 2013; Nogle et al. 2001; Pei and Jiang 2017; Yang et al. 2017b). In an endpoint assay, ramosin (P1) decreased the viability of the Gram-negative bacteria Escherichia coli and Pseudomonas aeruginosa in a concentration-dependent manner (Medina et al. 2022). While the decrease in viability of E. coli was already significant after incubation with 17.5 µM ramosin (and higher concentrations up to 70 µM), the inhibition of P. aeruginosa occurred starting from 35 µM ramosin. In a time-dependent bacterial growth experiment, ramosin was shown to act bactericidally against both, E. coli and P. aeruginosa. Ramosin exhibited a bacteriostatic effect against P. aeruginosa (Medina et al. 2022). In contrast, the antimicrobial activity of ramosin was less efficient against Gram-positive bacteria (i.e., Staphylococcus aureus and Bacillus cereus). Bacterial growth was inhibited at high concentrations of ramosin, which was ≥40 µM (S. aureus) and 70 µM (B. cereus). The activity of ramosin against both Gram-positive bacteria was characterized as a bactericidal effect in the higher concentrations, whereas at lower concentrations a bacteriostatic effect was observed (Medina et al. 2022). In case of Enterococcus faecalis, no decrease of cell viability could be observed with the highest applied concentration of ramosin (70 µM). Thus, the authors suggested that a higher level of ramosin might be required to observe an inhibitory effect against E. faecalis (Medina et al. 2022). With scanning electron microscopy (SEM), it was further confirmed that the antimicrobial effect of ramosin (20 µM) against S. aureus was caused by an interaction of the peptide with the bacterial membrane, which resulted in a significant disturbance of the bacterial surface (Medina et al. 2022). In contrast, a change of the membrane of E. coli could not be observed in the presence of ramosin, which might suggest another mechanism of action (Medina et al. 2022).
The cyclic peptide antillatoxin B (P3) was tested against the Gram-positive bacteria B. cereus and Listeria monocytogenes, as well as the Gram-negative Salmonella Typhimurium, showing inhibitory effects in the concentration range of ∼14–884 µM with the highest potency against B. cereus (Dussault et al. 2016).
Tested against the Gram-positive bacterium S. aureus and the Gram-negative bacteria E. coli and Bacillus subtilis, CFBD-1 (P9) was demonstrated to possess an antimicrobial activity against all tested bacteria (Meng et al. 2013). The minimal inhibitory concentrations (MIC) of CFBD-1 were ∼14,4 µM (S. aureus), ∼29.9 µM (B. subtilis), and ∼35.4 µM (E. coli). In addition, the fungus Candida albicans was sensitive towards CFBD-1 with a MIC of ∼44.2 µM (Meng et al. 2013). Although 100 % identity overlap with CFBD-1, salamandrin-I did not show any antibacterial activity against a selection of Gram-negative (i.e., E. coli and P. aeruginosa) and Gram-positive bacteria (i.e., S. aureus and E. faecalis) up to an applied concentration of ∼728 µM (Plácido et al. 2020).
The defensin Ad-Cath (P13) was demonstrated to exert antimicrobial activity against both, Gram-negative and Gram-positive bacteria, as well (Yang et al. 2017b). With MICs of 0–0.75 µM (E. coli) and 0.75–1.53 µM (Aeromonas hydrophila) against Gram-negative bacteria, as well as MICs of 0.75–1.53 µM (B. subtilis and Micrococcus luteus) and 1.53–3.05 µM (Staphylococcus epidermidis) it shows higher antimicrobial efficiency than the other caudate peptides (Yang et al. 2017b). SEM analysis revealed a clear damage of the bacterial membrane of E. coli and A. hydrophila in the presence of ∼6 µM Ad-Cath after 2 h incubation (Yang et al. 2017b), thus suggesting pore formation through the incorporation of the peptide into the membrane and subsequent leakage of cytoplasm as the mechanism of Ad-Cath-induced antimicrobial effects. In contrast to Ad-Cath (P13), Tk-Cath (P5; up to ∼32 µM tested) was shown to neither act antimicrobially against Gram-negative nor Gram-positive bacteria (Luo et al. 2021). Andricin-I (P14) also promotes antimicrobial processes against several Gram-positive and Gram-negative bacteria (Pei and Jiang 2017). The activity of andricin-I against Gram-negative bacteria is characterized by MICs of ∼8.3 µM (different E. coli strains, Alcaligenes faecalis, Serratia marcescens, and Salmonella Paratyphi) and ∼4.2 µM (P. aeruginosa and Enterobacter cloacae). The inhibitory action of andricin-I against Gram-positive bacteria was in most cases slightly lower with MICs of 33.5 µM (B. subtilis, S. aureus, and Listeria innocua) and 8.3 µM (Bacillus megaterium). However, andricin-I did not exhibit any antifungal activity against Aspergillus niger, C. albicans, and Saccharomyces cerevisiae (Pei and Jiang 2017).
Peptide fractions (not further characterized) from secretions of Plethodon cinereus (see Section 4.7) showed antibacterial activity against S. aureus and E. coli, while those of A. tridactylum were effective against various bacteria (i.e., Acinetobacter baumannii, Klebsiella aerogenes, K. pneumoniae, P. aeruginosa, S. aureus, and S. marcescens) (Fredericks and Dankert 2000; Pereira et al. 2023). The peptide secretions of C. alleganiensis showed also a broad antibacterial activity (against A. baumannii, Enterococcus faecium, K. pneumoniae, P. aeruginosa, and S. aureus), whereas those of D. fuscus, E. bislineata, E. longicauda, N. maculosus, S. intermedia, A. tridactylum each only killed bacteria of either S. aureus, K. aerogenes or P. aeruginosa (Pereira et al. 2023).
Interestingly, in Ambystoma triginum, peptide fractions were collected that exhibited an inhibitory effect not only against S. aureus and Klebsiella species but also against the fungal skin pathogen Batrachochytrium dendrobatidis (Sheafor et al. 2008). Similar observations were made with peptide fractions from secretions of A. tridactylum and A. maculatum that were able to inhibit growth of the fungal pathogens B. dendrobatidis and B. salamandrivorans (Pereira et al. 2018, 2023; Pereira and Woodley 2021).
5.2 Antioxidant function
In an ABTS- and DPPH-based assay, a free radical scavenging activity of salamandrin-I (P4) and Tk-Cath (P5) was demonstrated (Luo et al. 2021; Plácido et al. 2020). In salamandrin-I (P4), a crucial role of its tryptophan side chain for the antioxidant function was suggested, which was based on in silico studies (Plácido et al. 2020). In case of Tk-Cath (P5), the antioxidant function was proposed to support its wound-healing activity (see Section 5.6) (Luo et al. 2021).
5.3 Cytotoxicity
The cytotoxicity of ramosin (P1), salamandrin-I (P4), Tk-Cath (P5), Ad-Cath (P13), and andricin-I (P14) was tested by the 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) and/or resazurin-based cell viability assay (Luo et al. 2021; Medina et al. 2022; Pei and Jiang 2017; Plácido et al. 2020; Silva-Carvalho et al. 2023; Yang et al. 2017b). Upon exposure with ∼2.4 µM Ad-Cath (P13), >80 % of HEK 293T cells survived, whereas it showed significant cytotoxicity above a concentration of 4.9 µM (Yang et al. 2017b). Salamandrin-I (P4; up to ∼100 µM) had no significant cytotoxic effect on human microglia cells (HMC3) after 24 h incubation (Plácido et al. 2020). Injection (10–40 mg/kg) of salamandrin-I into Galleria mellonella larvae did also not affect their physiological condition, supporting the non-toxicity of the peptide in vivo (Plácido et al. 2020). However, recent studies revealed that salamandrin-I decreases the cell viability of human leukemia (HL)-60 cells with a half-maximal inhibitory concentration (IC50) of 27 µM and cell death was observed (Silva-Carvalho et al. 2023). The proliferation reduction was correlated with diminished CDK1 transcription levels, an increased release of LDH, and an impaired mitochondrial membrane potential (Silva-Carvalho et al. 2023). Cell death induction by salamandrin-I also involves the inflammasome as observed by increased NLRP1/3, CASP-1, and IL-1β expression upon salamandrin-I exposure (Silva-Carvalho et al. 2023).
In contrast, ramosin (P1; 0.5–50 µM) did not show any cytotoxicity against three human cell lines (i.e., HaCat, MCF-7, and PC-3) after 24 h treatment. Flow cytometry even confirmed a non-toxic effect upon exposure with up to 200 µM ramosin (Medina et al. 2022). Similar results were obtained with andricin-I (P14) (Pei and Jiang 2017). It was also considered as “relatively safe” due to >80 % viability of the human cell lines HL-7702 and HREpiC cells after incubation with ∼106 µM andricin-I (Pei and Jiang 2017). Exposure of HaCat cells with up to ∼32 µM Tk-Cath (P5) induced no significant cell death (only in case of 16.6 %) (Luo et al. 2021). In contrast, it even promoted cell proliferation of HaCat cells at lower concentrations applied (i.e., 1–2 µM) (Luo et al. 2021).
5.4 Hemolytic activity
CFBD-1 (P9) has been characterized as a hemolytic peptide by using human and rabbit red blood cells (RBCs) (Meng et al. 2013). By spectroscopic monitoring of the amount of released hemoglobin, a hemolysis rate of ∼2.5 % and 3.2 % with human and rabbit RBCs could be determined in the presence of ∼88.5 µM CFBD-1, respectively. Similar results were obtained with Tk-Cath (P5; up to ∼32 µM), which induced hemolysis with a rate <2 % in mouse RBCs (Luo et al. 2021). Non-characterized peptide fractions from secretions of Plethodon cinereus (Plethodontidae) (see Section 4.7) were also demonstrated to induce hemolysis of guinea pig RBCs up to a lysis level of ∼84 % after ∼1 h (Fredericks and Dankert 2000).
In contrast, ramosin (P1; up to ∼128 µM), salamandrin-I (P4; up to 100 µM), and andricin-I (P14; up to ∼52 µM) were shown to be non-hemolytic against human RBCs (Medina et al. 2022; Pei and Jiang 2017; Plácido et al. 2020). Similarly, the peptide fractions from secretions of the following species did not show any hemolytic potential against human RBCs: Ambystomatidae: A. maculatum; Amphiumidae: A. tridactylum; Cryptobranchidae: C. alleganiensis; Plethodontidae: D. fuscus, E. bislineata, E. longicauda; Proteidae: N. beyeri, N. maculosus; Sirenidae: S. intermedia (Pereira et al. 2023).
5.5 Antiinflammatory activity
So far, Tk-Cath (P5) is the only reported caudate skin peptide exerting a strong antiinflammatory activity (Luo et al. 2021). In fact, Tk-Cath is a potent inhibitor of lipopolysaccharide (LPS)-induced proinflammatory Toll-like receptor 4 (TLR4) signaling in a mouse macrophage cell line (RAW 264.7) (Luo et al. 2021). The inhibition was observed to occur in a dose-dependent manner (∼0.8–3.2 µM) as evident by the level of effector protein phosphorylation (i.e., of ERK, JNK, and p38). In leukocytes of the African clawed frog, the antiinflammatory activity of Tk-Cath towards LPS-induced signaling was confirmed through the significantly reduced expression profiles of tumor necrosis factor (TNF; by 37 %), interleukin 1β (by 54 %), and the inducible nitric oxide synthase (iNOS; by 59 %) and the decreased presence of the cytokines TNF (by 33 %) and interleukin 6 (by 68 %) (Luo et al. 2021).
5.6 Wound healing-promoting activity
So far, two caudate skin peptides [i.e., Tk-Cath (P5) and tylotoin (P7)] were described to exert wound healing-promoting functions (Luo et al. 2021; Mu et al. 2014). Due to their chemotactic features, these are directly cross-linked with the antiinflammatory effects (see Section 5.5.). Tk-Cath (P5) promotes wound healing by improving the keratinocyte migration, as has been observed in a cell scratch assay upon exposure of HaCaT cells with ∼3 µM Tk-Cath. After 48 h, a repair rate of ∼82.5 % was reached (Luo et al. 2021). In addition, keratinocyte proliferation was enhanced in a dose-dependent manner in the presence of Tk-Cath (∼0.81–6.45 µM). In a mouse full-thickness wound model, Tk-Cath was thus able to significantly accelerate wound healing. On molecular level, this wound-healing capability of Tk-Cath was associated with enhanced production of TNF, the chemokines MCP-1 and CXCL1 as well as the growth factor TGF-β1 (Luo et al. 2021). Furthermore, Tk-Cath induced the mitogen-activated protein kinase (MAPK) signaling pathway in RAW 264.7 cells, further supporting the wound healing process (Luo et al. 2021). Similar effects were observed for the peptide tylotoin (P7; ∼1.4–14 µM), which increased proliferation of keratinocytes (HaCat) but also of fibroblasts (HSFs) and endothelial cells (HUVECs) (Mu et al. 2014). In the wound healing scratch assay, tylotoin enhanced keratinocyte migration. After 48 h, a repair rate of ∼80 % was reached (Mu et al. 2014). In contrast, exposure of HUVECs with tylotoin resulted in endothelial cell tube formation (Mu et al. 2014). Furthermore, as in case of Tk-Cath, also tylotoin induces MAPK signaling pathway in RAW264-7 macrophage cells, resulting in cytokine (i.e., TGF- β1 and interleukin 6) secretion (Luo et al. 2021; Mu et al. 2014). However, the secretion of interleukin 1β, EGF, and TNF was not affected by tylotoin. Finally, topical administration of tylotoin led to significant acceleration of wound healing along with rapid macrophage migration in a mouse full-thickness wound model (Mu et al. 2014).
5.7 Neurotoxicity
Antillatoxin B (P3) was shown to activate voltage-gated sodium channels in mouse neuro-2a neuroblastoma cells with a half maximal effective concentration (EC50) of 1.77 µM, resulting in its neurotoxic action due to continuous stimulation (Nogle et al. 2001). This led to a highly ichthyotoxicity in goldfish with a lethal dose of 1.0 µM (Nogle et al. 2001).
5.8 Myotropic activity
In in vivo studies with salamandrin-I (P4), a histological analysis of the larvae of G. mellonella revealed after injection of the peptide (40 mg/kg) a slight change of the muscle tissue morphology suggesting an increase muscle activity and, thus, a potential myotropic effect of the peptide (Plácido et al. 2020). In contrast, a myotropic effect of CCK-TV (P6) was already proven (Jiang et al. 2015). As tested by the use of porcine gallbladder strips, the peptide was capable of inducing muscle contractions in a dose-dependent manner characterized by a low half-maximal effective concentration (EC50) of 13.6 nM.
5.9 Pheromone activity
The pheromone signals of caudate amphibians were proposed to be based on a multicomponent system (see Section 2.5), thereby also involving peptides [i.e., silefrin (P8), sodefrin (P10), aonirin (P11), and imorin (P12)]. Sodefrin (P10) exerts a species-specific female-attracting activity, which requires at least a concentration of 0.1–1.0 pM in water to attract female individuals of the salamandrid Cynops pyrroghaster (Kikuyama et al. 1995). Males secrete sodefrin from abdominal glands and are recognized by females through binding to receptors at the vomeronasal epithelium (Kikuyama et al. 1995). In the related C. ensicauda, sodefrin was shown to not attract females. In contrast, a variant of sodefrin, silefrin (P8), was found to act similarly but also in a species-specific way (Kikuyama et al. 1995; Yamamoto et al. 2000). While the peptide pheromones silefrin (P8) and sodefrin (P10) are produced by male newts, the peptide imorin (P12) was detected on the skin of female C. pyrroghaster acting as a male attractant thereby stimulating the vomeronasal epithelial cells of the male newts (Nakada et al. 2017).
Beside species-specific peptide pheromones, also already one region-specific pheromone, aonirin, a variant of sodefrin, was identified in C. pyrroghaster (Nakada et al. 2007). Although only differing by one amino acid residue at position 8 (Leu8Val), the pheromone was either much less or not attractive for individuals from other areas, which might lead to the development of a separate species due to reproductive isolation in future (Nakada et al. 2007).
6 Potential future applications and perspectives
While components of secretions from other amphibians (e.g., bufotenine and bombesin) have been considered as drug candidates for targeting various pathophysiological states (Rodriguez et al. 2017; Sousa et al. 2017), the potential of caudate amphibian skin secretions and especially of skin peptides seems to be still inconspicuous and underestimated.
Based on the observed bioactivities, skin peptides from caudates were already discussed as novel therapeutic agents for the treatment of bacterial infections, wounds, and cancer as well as potential antiaging agents (Luo et al. 2021; Medina et al. 2022; Mu et al. 2014; Pei and Jiang 2017; Plácido et al. 2020; Silva-Carvalho et al. 2023; Yang et al. 2017b). Among these, tylotoin is probably the most widely studied peptide in terms of pharmaceutical applications. Due to its wound-healing properties the application of a slightly modified tylotoin with the prolonged sequence RKCVRQNNKRVCK (“cathelicidin-TV2”) for skin repair with reduced scar generation has been proposed (pending Chinese patent CN104288750A). Efforts were also made to design a drugable version of tylotoin by coupling with a cell-penetrating peptide or incorporation in nanoparticles, which improved the cellular uptake of the peptide and, thus, enabled more targeted treatment (Horn and Neundorf 2018; Wang et al. 2022).
In addition, the skin secretions of A. davidianus (Cryptobranchidae) were proposed as an outstanding adhesive raw biomaterial with the potential of embedding active substances for administration on wounds for faster cell proliferation (Liang et al. 2023). Similar potential usage was suggested for the adhesive skin secretions of Plethodon shermani (Plethodontidae) and Ambystoma opacum (Ambystomatidae) (Von Byern et al. 2017). Finally, these examples demonstrate the capacity and applicability of caudate amphibian skin peptides for biomedical purposes in future.
About the authors

Lorena Kröner received her M.Ed. in Chemistry and Mathematics from University of Koblenz in 2023. Currently, she is a Ph.D. student in Bioorganic Chemistry at the University of Koblenz. Her current research, under the supervision of Jun.-Prof. Marie-T. Hopp, is focused on the investigation of amphibian skin molecules and their physiological role.

Stefan Lötters is a professor of Biogeography at Trier University, leading an Ecology, Evolution & Systematics Group. Using amphibians as a main model group, his research is focused on speciation and character evolution with a particular geographic focus on the Neotropics, as well as the response to new pathogens. The latter is a serious conservation issue in many amphibians including salamanders and newts in Europe.

Marie-T. Hopp obtained her master’s degree in Chemistry and Biology (2017) at the University of Koblenz-Landau. In 2021, she completed her PhD in Pharmaceutical Biochemistry and Bioanalytics at the University of Bonn. In 2023, she joined the University of Koblenz as a junior professor for Bioorganic Chemistry with a focus on peptide chemistry. She aims to characterize heme-protein interactions in the blood coagulation system as well as to explore the chemistry and bioactivity of amphibian skin peptides at molecular level.
-
Research ethics: Not applicable.
-
Author contributions: The manuscript was written and reviewed through the contribution of all authors. All authors have accepted responsibility for the entire content of the submitted manuscript and approved submission.
-
Competing interests: The authors declare no conflicts of interest regarding this article.
-
Research funding: Not applicable.
-
Data availability: Not applicable.
References
Abal, P., Louzao, M.C., Antelo, A., Alvarez, M., Cagide, E., Vilariño, N., Vieytes, M.R., and Botana, L.M. (2017). Acute oral toxicity of tetrodotoxin in mice: determination of lethal dose 50 (LD50) and no observed adverse effect level (NOAEL). Toxins 9: 75, https://doi.org/10.3390/toxins9030075.Search in Google Scholar
Anastasi, A., Erspamer, V., and Bertaccini, G. (1965). Occurrence of bradykinin in the skin of Rana temporaria. Comp. Biochem. Physiol. 14: 43–52, https://doi.org/10.1016/0010-406x(65)90007-1.Search in Google Scholar
Anastasi, A., Erspamer, V., and Bucci, M. (1971). Isolation and structure of bombesin and alytesin, 2 analogous active peptides from the skin of the European amphibians Bombina and Alytes. Experientia 27: 166–167, https://doi.org/10.1007/bf02145873.Search in Google Scholar
Bachmayer, H. and Michl, H. (1965). Über hämolytisch wirksame Stoffe in Molchgiften. Monatsh. Chem. 96: 1166–1172, https://doi.org/10.1007/bf00904262.Search in Google Scholar
Barros, A.L.A.N., Hamed, A., Marani, M., Moreira, D.C., Eaton, P., Plácido, A., Kato, M.J., and Leite, J.R.S.A. (2022). The arsenal of bioactive molecules in the skin secretion of urodele amphibians. Front. Pharmacol 12: 810821, https://doi.org/10.3389/fphar.2021.810821.Search in Google Scholar
Becker, H. (1986). Inhaltsstoffe von Feuer- und Alpensalamander. Pharm. unserer Zeit 15: 97–128, https://doi.org/10.1002/pauz.19860150402.Search in Google Scholar
Brown, D.D. and Cai, L. (2007). Amphibian metamorphosis. J. Dev. Biol. 306: 20–33, https://doi.org/10.1016/j.ydbio.2007.03.021.Search in Google Scholar
Çömden, E.A., Yenmiş, M., and Çakır, B. (2023). The complex bridge between aquatic and terrestrial life: skin changes during development of amphibians. J. Dev. Biol. 11: 6, https://doi.org/10.3390/jdb11010006.Search in Google Scholar
Croce, G. and Bolognani, L. (1975). Lipid components in the skin secretions of amphibia – I. Cholesterol. Comp. Biochem. Physiol. B. 52: 307–309, https://doi.org/10.1016/0305-0491(75)90069-3.Search in Google Scholar
Csordaś, A. and Michl, A. (1970). Isolierung und Strukturaufklärung eines hämolytisch wirkenden polypeptides aus dem Abwehrsekret europäischer Unken. Monatsh. Chem. 101: 182–189.10.1007/BF00907538Search in Google Scholar
Daly, J.W., Spande, T.F., and Garraffo, H.M. (2005). Alkaloids from amphibian skin: a tabulation of over eight-hundred compounds. J. Nat. Prod. 68: 1556–1575, https://doi.org/10.1021/np0580560.Search in Google Scholar
DeBruin, J.H., Wilburn, D.B., Feldhoff, R.C., and Staub, N.L. (2023). Presence of sodefrin precursor-like factor pheromone candidates in mental and dorsal tail base glands in the plethodontid salamander, Karsenia koreana. PLoS One 18: e0289296, https://doi.org/10.1371/journal.pone.0289296.Search in Google Scholar
Degani, G., Am, G.I., Yatom, N., Khatib, S., and Musa, S. (2023a). Eastern fire salamander (Salamandra infraimmaculata) skin mucus metabolites – LC-MS/MS analysis. Environ. Anal. Eco. Stud. 10: 1187–1194.10.31031/EAES.2023.10.000747Search in Google Scholar
Degani, G., Peretz, E., and Musa, S. (2023b). Skin mucus metabolites in Salamandra infraimmaculata from various habitats. Endocrinol. Metab. Nurtr. 2: 1–9, https://doi.org/10.33425/2833-0307.1014.Search in Google Scholar
Dougherty, P.G., Sahni, A., and Pei, D. (2019). Understanding cell penetration of cyclic peptides. Chem. Rev. 119: 10241–10287, https://doi.org/10.1021/acs.chemrev.9b00008.Search in Google Scholar
Dussault, D., Vu, K.D., Vansach, T., Horgen, F.D., and Lacroix, M. (2016). Antimicrobial effects of marine algal extracts and cyanobacterial pure compounds against five foodborne pathogens. Food Chem. 199: 114–118, https://doi.org/10.1016/j.foodchem.2015.11.119.Search in Google Scholar
Eom, J., Jung, Y.R., and Park, D. (2009). F-series prostaglandin function as sex pheromones in the Korean salamander Hynobius leechii. Comp. Biochem. Physiol. 154: 61–69, https://doi.org/10.1016/j.cbpa.2009.05.006.Search in Google Scholar
Fawell, J.K., Mitchell, R.E., Hill, R.E., and Everett, D.J. (1999). The toxicity of cyanobacterial toxins in the mouse: II Anatoxin-a. Hum. Exp. Toxicol. 18: 168–173, https://doi.org/10.1177/096032719901800306.Search in Google Scholar
Fredericks, L.P. and Dankert, J.R. (2000). Antibacterial and hemolytic activity of the skin of the terrestrial salamander, Plethodon cinereus. J. Exp. Zool. 287: 340–345, https://doi.org/10.1002/1097-010x(20001001)287:5<340::aid-jez2>3.0.co;2-9.10.1002/1097-010X(20001001)287:5<340::AID-JEZ2>3.3.CO;2-0Search in Google Scholar
Gall, B.G., Stokes, A.N., Brodie III, E.D., and Brodie, Jr., E.D. (2022). Tetrodotoxin levels in lab-reared rough-skinned newts (Taricha granulosa) after 3 years and comparison to wild-caught juveniles. Toxicon 213: 7–12, https://doi.org/10.1016/j.toxicon.2022.04.007.Search in Google Scholar
Gasteiger, E., Hoogland, C., Gattiker, A., Duvaud, S., Wilkins, M.R., Appel, R.D., and Bairoch, A. (2005). Protein identification and analysis tools on the Expasy server. In: Walker, J.M. (Ed.). The proteomics protocols handbook. Humana Press, Totowa.10.1385/1-59259-890-0:571Search in Google Scholar
Geng, X., Wei, H., Shang, H., Zhou, M., Chen, B., Zhang, F., Zang, X., Li, P., Sun, J., Che, J., et al.. (2015). Proteomic analysis of the skin of Chinese giant salamander (Andrias davidianus). J. Proteomics 119: 196–208, https://doi.org/10.1016/j.jprot.2015.02.008.Search in Google Scholar
Habermehl, G. (1965). Cholesterin und Cholesterinester aus dem Hautdrüsensekret von Salamandra maculosa taeniata. Adv. Cycloaddit. 680: 104–107, https://doi.org/10.1002/jlac.19646800113.Search in Google Scholar
Hall, K.W., Eisthen, H.L., and Williams, B.L. (2016). Proteinaceous pheromone homologs identified from the cloacal gland transcriptome of a male axolotl, Ambystoma mexicanum. PLoS One 11: e0146851, https://doi.org/10.1371/journal.pone.0146851.Search in Google Scholar
Hamning, V.K., Yanites, H.L., and Peterson, N.L. (2000). Characterization of adhesive and neurotoxic components in skin granular gland secretions of Ambystoma tigrinum. Copeia 2000: 856–859, https://doi.org/10.1643/0045-8511(2000)000[0856:coaanc]2.0.co;2.10.1643/0045-8511(2000)000[0856:COAANC]2.0.CO;2Search in Google Scholar
Hanifin, C.T. (2010). The chemical and evolutionary ecology of tetrodotoxin (TTX) toxicity in terrestrial vertebrates. Mar. Drugs 8: 577–593, https://doi.org/10.3390/md8030577.Search in Google Scholar
Hanifin, C.T., Brodie III, E.D., and Brodie Jr, E.D. (2003). Tetrodotoxin levels in eggs of the rough-skin newt, Taricha granulosa, are correlated with female toxicity. J. Chem. Ecol. 29: 1729–1739, https://doi.org/10.1023/a:1024885824823.Search in Google Scholar
Hedrick, M.S., Hillman, S.S., Drewes, R.C., and Withers, P.C. (2013). Lymphatic regulation in nonmammalian vertebrates. J. Appl. Physiol. 115: 297–308, https://doi.org/10.1152/japplphysiol.00201.2013.Search in Google Scholar
Hoffman, C.W. and Dent, J.N. (1977). Effects of neurotransmitters upon the discharge of secretory product from the cutaneous glands of the red-spotted newt. J. Exp. Zool. 202: 155–161, https://doi.org/10.1002/jez.1402020203.Search in Google Scholar
Hopkins, G.R. and Migabo, S.W. (2010). Antipredator skin secretions of the long-toed salamander (Ambystoma macrodactylum) in its northern range. J. Herpetol. 44: 627–633, https://doi.org/10.1670/09-216.1.Search in Google Scholar
Horn, M. and Neundorf, I. (2018). Design of a novel cell-permeable chimeric peptide to promote wound healing. Sci. Rep. 8: 16279, https://doi.org/10.1038/s41598-018-34684-1.Search in Google Scholar
Houck, L.D., Palmer, C.A., Watts, R.A., Arnold, S.J., Feldhoff, P.W., and Feldhoff, R.C. (2007). A new vertebrate courtship pheromone, PMF, affects female receptivity in a terrestrial salamander. Anim. Behav. 73: 315–320, https://doi.org/10.1016/j.anbehav.2006.07.008.Search in Google Scholar
Houck, L.D., Watts, R.A., Mead, L.M., Palmer, C.A., Arnold, S.J., Feldhoff, P.W., and Feldhoff, R.C. (2008). A candidate vertebrate pheromone, SPF, increases female receptivity in a salamander. In: Hurst, J.L., Beynon, R.J., Roberts, S.C., and Wyatt, T.D. (Eds.). Chemical signals in vertebrates 11. Springer, New York.Search in Google Scholar
Huan, Y., Kong, Q., Mou, H., and Yi, H. (2020). Antimicrobial peptides: classification, design, application and research progress in multiple fields. Front. Microbiol. 11: 582779, https://doi.org/10.3389/fmicb.2020.582779.Search in Google Scholar
Janssenswillen, S., Vandebergh, W., Treer, D., Willaert, B., Maex, M., Van Bocxlaer, I., and Bossuyt, F. (2014). Origin and diversification of a salamander sex pheromone system. Mol. Biol. Evol. 32: 472–480, https://doi.org/10.1093/molbev/msu316.Search in Google Scholar
Janssenswillen, S., Willaert, B., Treer, D., Vandebergh, W., Bossuyt, F., and Van Bocxlaer, I. (2015). High pheromone diversity in the male cheek gland of the red-spotted newt Notophthalmus viridescens (Salamandridae). BMC Evol. Biol. 15: 54, https://doi.org/10.1186/s12862-015-0333-1.Search in Google Scholar
Jiang, W., Hakim, M., Luo, L., Li, B., Yang, S., Song, Y., Lai, R., and Lu, Q. (2015). Purification and characterization of cholecystokinin from the skin of salamander Tylototriton verrucosus. Zool. Res. 36: 174–177.Search in Google Scholar
Jumper, J., Evans, R., Pritzel, A., Green, T., Figurnov, M., Ronneberger, O., Tunyasuvunakool, K., Bates, R., Žídek, A., Potapenko, A., et al.. (2021). Highly accurate protein structure prediction with AlphaFold. Nature 596: 583–589, https://doi.org/10.1038/s41586-021-03819-2.Search in Google Scholar
Karış, M., Şener, D., Yalçın, H.T., Nalbantsoy, A., and Göçmen, B. (2018). Major biological activities and protein profiles of skin secretions of Lissotriton vulgaris and Triturus ivanbureschi. Turkish J. Biochem. 43: 605–612, https://doi.org/10.1515/tjb-2017-0306.Search in Google Scholar
Kikuyama, S., Toyoda, F., Ohmiya, Y., Matsuda, K., Tanaka, S., and Hayashi, H. (1995). Sodefrin: a female-attracting peptide pheromone in newt cloacal glands. Science 267: 1643–1645, https://doi.org/10.1126/science.7886452.Search in Google Scholar
Kim, K.-H. and Seong, B.L. (2001). Peptide amidation: production of peptide hormones in vivo and in vitro. Biotech. Bioproc. Eng. 6: 244–251, https://doi.org/10.1007/bf02931985.Search in Google Scholar
Knepper, J., Lüddecke, T., Preißler, K., Vences, M., and Schulz, S. (2019). Isolation and identification of alkaloids from poisons of fire salamanders (Salamandra salamandra). J. Nat. Prod. 82: 1319–1324, https://doi.org/10.1021/acs.jnatprod.9b00065.Search in Google Scholar
Kudo, Y., Hanifin, C.T., Kotaki, Y., and Yotsu-Yamashita, M. (2020). Structures of N-hydroxy-type tetrodotoxin analogues and bicyclic guanidinium compounds found in toxic newts. J. Nat. Prod. 83: 2706–2717, https://doi.org/10.1021/acs.jnatprod.0c00623.Search in Google Scholar
Kudo, Y., Yasumoto, T., Konoki, K., Cho, Y., and Yotsu-Yamashita, M. (2012). Isolation and structural determination of the first 8-epi-type tetrodotoxin analogs from the newt, Cynops ensicauda popei, and comparison of tetrodotoxin analogs profiles of this newt and the puffer fish, Fugu poecilonotus. Mar. Drugs 10: 655–667, https://doi.org/10.3390/md10030655.Search in Google Scholar
Kudo, Y. and Yotsu-Yamashita, M. (2019). Isolation and biological activity of 8-epitetrodotoxin and the structure of a possible biosynthetic shunt product of tetrodotoxin, Cep-226A, from the newt Cynops ensicauda popei. J. Nat. Prod. 82: 1656–1663, https://doi.org/10.1021/acs.jnatprod.9b00178.Search in Google Scholar
Largen, W. and Woodley, S.K. (2008). Cutaneous tail glands, noxious skin secretions, and scent marking in a terrestrial salamander (Plethodon shermani). Herpetologica 64: 270–280, https://doi.org/10.1655/08-010.1.Search in Google Scholar
Liang, H., Wang, X.-T., Ge, W.-Y., Zhang, R., Liu, J., Chen, L.-L., Xi, X.-L., Guo, W.-H., and Yin, D.-C. (2023). Andrias davidianus mucus-based bioadhesive with enhanced adhesion and wound healing properties. ACS Appl. Mater. Interfaces 15: 49931–49942, https://doi.org/10.1021/acsami.3c04148.Search in Google Scholar
Lüddecke, T., Schulz, S., Steinfartz, S., and Vences, M. (2018). A salamander’s toxic arsenal: review of skin poison diversity and function in true salamanders, genus Salamandra. Sci. Nat. 105: 56, https://doi.org/10.1007/s00114-018-1579-4.Search in Google Scholar
Luo, X., Ouyang, J., Wang, Y., Zhang, M., Fu, L., Xiao, N., Gao, L., Zhang, P., Zhou, J., and Wang, Y. (2021). A novel anionic cathelicidin lacking direct antimicrobial activity but with potent anti-inflammatory and wound healing activities from salamander Tylototriton kweichowensis. Biochimie 191: 37–50, https://doi.org/10.1016/j.biochi.2021.08.007.Search in Google Scholar
Maex, M., Treer, D., De Greve, H., Proost, P., Van Bocylaer, I., and Bossuyt, F. (2018). Exaptation as a mechanism for functional reinforcement of an animal pheromone system. Curr. Biol. 28: 2955–2960, https://doi.org/10.1016/j.cub.2018.06.074.Search in Google Scholar
Mebs, D. and Pogoda, W. (2005). Variability of alkaloids in the skin secretion of the European fire salamander (Salamandra salamadra terrestris). Toxicon 45: 603–606, https://doi.org/10.1016/j.toxicon.2005.01.001.Search in Google Scholar
Medina, L., Guzmán, F., Álvarez, C., Delgado, J.P., and Carbonell-M, B. (2022). Ramosin: the first antibacterial peptide identified on Bolitoglossa ramosi Colombian salamander. Pharmaceutics 14: 2579, https://doi.org/10.3390/pharmaceutics14122579.Search in Google Scholar
Meng, E.C., Goddard, T.D., Pettersen, E.F., Couch, G.S., Pearson, Z.J., Morris, J.H., and Ferrin, T.E. (2023). UCSF ChimeraX: tools for structure building and analysis. Protein Sci. 32: e4792, https://doi.org/10.1002/pro.4792.Search in Google Scholar
Meng, P., Yang, S., Shen, C., Jiang, K., Rong, M., and Lai, R. (2013). The first salamander defensin antimicrobial peptide. PLoS One 8: e83044, https://doi.org/10.1371/journal.pone.0083044.Search in Google Scholar
Mu, L., Tang, J., Liu, H., Shen, C., Rong, M., Zhang, Z., and Lai, R. (2014). A potential wound-healing-promoting peptide from salamander skin. FASEB J. 28: 3919–3929, https://doi.org/10.1096/fj.13-248476.Search in Google Scholar
Nakada, T., Toyoda, F., Iwata, T., Yamamoto, K., Conlon, J.M., Kato, T., and Kukuyama, S. (2007). Isolation, characterization and bioactivity of a region-specific pheromone, [Val8]sodefrin from the newt Cynops pyrrhogaster. Peptides 28: 774–780, https://doi.org/10.1016/j.peptides.2006.12.006.Search in Google Scholar
Nakada, T., Toyoda, F., Matsuda, K., Nakakura, T., Hasunuma, I., Yamamoto, K., Onoue, S., Yokosuka, M., and Kikuyama, S. (2017). Imorin: a sexual attractiveness pheromone in female red-bellied newts (Cynops pyrrhogaster). Sci. Rep. 7: 41334, https://doi.org/10.1038/srep41334.Search in Google Scholar
Nikolaieva, I., Dudkina, Y., Oliinyk, D., Oskyrko, O., Marushchak, O., Halenova, T., and Savchuk, O. (2018). Amphibian skin secretions: a potential source of proteolytic enzymes. Biotechnol. Acta 11: 42–48, https://doi.org/10.15407/biotech11.05.042.Search in Google Scholar
Nogle, L.M., Okino, T., and Gerwick, W.H. (2001). Antillatoxin B, a neurotoxic lipopeptide from the marine cyanobacterium Lyngbya majuscula. J. Nat. Prod. 64: 983–985, https://doi.org/10.1021/np010107f.Search in Google Scholar
Ong, D., Ismail, M.N., and Shahrudin, S. (2021). Protein composition and biomedical potential of the skin secretion of Hylarana erythraea (Schlegel, 1837) (Anura: Ranidae) from Langkawi Archipelago, Kedah, Peninsular Malaysia. Int. J. Pept. Res. Ther. 27: 2125–2133, https://doi.org/10.1007/s10989-021-10240-w.Search in Google Scholar
Pei, J. and Jiang, L. (2017). Antimicrobial peptide from mucus of Andrias davidianus: screening and purification by magnetic cell membrane separation technique. Int. J. Antimicrob. Agents 50: 41–46, https://doi.org/10.1016/j.ijantimicag.2017.02.013.Search in Google Scholar
Pereira, K.E., Cava, K.A., and Woodley, S.K. (2021). An evaluation of immersive and handling methods for collecting salamander skin peptides. J. Herpetol. 55: 318–324, https://doi.org/10.1670/20-122.Search in Google Scholar
Pereira, K.E., Crother, B.I., Sever, D.M., Fontenot, C.L., Pojman, J.A., Wilburn, D.B., and Woodley, S.K. (2018). Skin glands of an aquatic salamander vary in size and distribution and release antimicrobial secretions effective against chytrid fungal pathogens. J. Exp. Biol. 221: jeb183707, https://doi.org/10.1242/jeb.183707.Search in Google Scholar
Pereira, K.E., Deslouches, J.T., Deslouches, B., and Woodley, S.K. (2023). In vitro investigation of the antibacterial activity of salamander skin peptides. Curr. Microbiol. 80: 214, https://doi.org/10.1007/s00284-023-03320-1.Search in Google Scholar
Pereira, K.E. and Woodley, S.K. (2021). Skin defenses of North American salamanders against a deadly salamander fungus. Anim. Conserv. 24: 552–567, https://doi.org/10.1111/acv.12666.Search in Google Scholar
Plácido, A., Bueno, J., Barbosa, E.A., Moreira, D.C., Dias, J.d.N., Cabral, W.F., Albuquerque, P., Bessa, L.J., Freitas, J., Kuckelhaus, S.A.S., et al.. (2020). The antioxidant peptide salamandrin-I: first bioactive peptide identified from skin secretion of Salamandra genus (Salamandra salamandra). Biomolecules 10: 512, https://doi.org/10.3390/biom10040512.Search in Google Scholar
Qu, M., Tong, C., Kong, L., Yan, X., Chernikov, O., Lukyanov, P.A., Jin, Q., and Li, W. (2014). Purification of a secreted lectin from Andrias davidianus skin and its antibacterial activity. Comp. Biochem. Physiol. C. Toxicol. Pharmacol. 167: 140–146, https://doi.org/10.1016/j.cbpc.2014.08.008.Search in Google Scholar
Rickrode, T.E., Mueller, C.F., and Tylor, D. (1986). Identification and antibiotic activity of fatty acids in dermal secretions of Plethodon cinereus. Am. Midl. Nat. 115: 198–200, https://doi.org/10.2307/2425850.Search in Google Scholar
Rodriguez, C., Rollins-Smith, L., Ibáñez, R., Durant-Archibold, A.A., and Gutiérrez, M. (2017). Toxins and pharmacologically active compounds from species of the family Bufonidae (Amphibia, Anura). J. Ethnopharmacol. 198: 235–254, https://doi.org/10.1016/j.jep.2016.12.021.Search in Google Scholar
Rollins-Smith, L.A., Reinert, L.K., Miera, V., and Conlon, J.M. (2002). Antimicrobial peptide defense of the Tarahumara frog, Rana tarahumarae. Biochem. Biophys. Res. Commun. 297: 361–367, https://doi.org/10.1016/s0006-291x(02)02217-9.Search in Google Scholar
Rollmann, S.M., Houck, L.D., and Feldhoff, R.C. (1999). Proteinaceous pheromone affecting female receptivity in a terrestrial salamander. Science 285: 1907–1909, https://doi.org/10.1126/science.285.5435.1907.Search in Google Scholar
Roseghini, M., Erspamer, F., Severini, C., and Simmaco, M. (1989). Biogenic amines and active peptides in extracts of thirty-two European amphibian species. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 94: 455–460, https://doi.org/10.1016/0742-8413(89)90097-2.Search in Google Scholar
Saporito, R.A., Donnelly, M.A., Spande, T.F., and Garraffo, H.M. (2012). A review of chemical ecology in poison frogs. Chemoecol 22: 159–168, https://doi.org/10.1007/s00049-011-0088-0.Search in Google Scholar
Sheafor, B., Davidson, E.W., Parr, L., and Rollins-Smith, L. (2008). Antimicrobial peptide defenses in the salamander, Ambystoma tigrinum, against emerging amphibian pathogens. J. Wildl. Dis. 44: 226–236, https://doi.org/10.7589/0090-3558-44.2.226.Search in Google Scholar
Shen, Y., Maupetit, J., Derreumaux, P., and Tufféry, P. (2014). Improved PEP-FOLD approach for peptide and miniprotein structure prediction. J. Chem. Theory Comput. 10: 4745–4758, https://doi.org/10.1021/ct500592m.Search in Google Scholar
Silva-Carvalho, A.E., de Oliveira, N.N., Machado, J.V.L., Moreira, D.C., Brand, G.D., Leite, J.R.S.A., Plácido, A., Eaton, P., and Saldanha-Araujo, F. (2023). The peptide salamandrin-I modulates components involved in pyroptosis and induces cell death in human leukemia cell line HL-60. Pharmaceutics 15: 1864, https://doi.org/10.3390/pharmaceutics15071864.Search in Google Scholar
Sousa, L.Q., Machado, K.D., Oliveira, S.F., Araújo, L.D., Monção-Filho, E.D., Melo-Cavalcante, A.A., Vieira-Júnior, G.M., and Ferreira, P.M. (2017). Bufadienolides from amphibians: a promising source of anticancer prototypes for radical innovation, apoptosis triggering and Na+/K+-ATPase inhibition. Toxicon 127: 63–76, https://doi.org/10.1016/j.toxicon.2017.01.004.Search in Google Scholar
Teranishi, H., Muneoka, Y., Takao, T., Shimonishi, Y., and Kojima, M. (2004). Isolation and characterization of four VIP-related peptides from red-bellied newt, Cynops pyrrhogaster. Regul. Pep. 123: 173–179, https://doi.org/10.1016/j.regpep.2004.04.024.Search in Google Scholar
Thompson, A.H., Bjourson, A.J., Orr, D.F., Shaw, C., and Mcclean, S. (2007). Amphibian skin secretomics: application of parallel quadrupole time-of-flight mass spectrometry and peptide precursor cDNA cloning to rapidly characterize the skin secretory peptidome of Phyllomedusa hypochondrialis azurea: discovery of a novel peptide family, the hyposins. J. Proteome Res. 6: 3604–3613, https://doi.org/10.1021/pr0702666.Search in Google Scholar
Toledo, R.C. and Jared, C. (1993). Cutaneous adaptations to water balance in amphibians. Comp. Biochem. Physiol. 105: 593–608, https://doi.org/10.1016/0300-9629(93)90259-7.Search in Google Scholar
Toledo, R.C. and Jared, C. (1995). Cutaneous granular glands and amphibian venoms. Comp. Biochem. Physiol. 111: 1–29, https://doi.org/10.1016/0300-9629(95)98515-i.Search in Google Scholar
Treer, D., Maex, M., Van Bocxlaer, I., Proost, P., and Bossuyt, F. (2017). Divergence of species-specific protein sex pheromone blends in two related, nonhybridizing newts (Salamandridae). Mol. Ecol. 27: 508–519, https://doi.org/10.1111/mec.14398.Search in Google Scholar
Umile, T.P., McLaughlin, P.J., Johnson, K.R., Honarvar, S., Blackman, A.L., Burzynski, E.A., Davis, R.W., Teotonio, T.L., Hearn, G.W., Hughey, C.A., et al.. (2014). Nonlethal amphibian skin swabbing of cutaneous natural products for HPLC fingerprinting. Anal. Methods 6: 3277–3284, https://doi.org/10.1039/c4ay00566j.Search in Google Scholar
Vences, M., Sanchez, E., Hauswaldt, J.S., Eikelmann, D., Rodríguez, A., Carranza, S., Donaire, D., Gehara, M., Helfer, V., Lötters, S., et al.. (2014). Nuclear and mitochondrial multilocus phylogeny and survey of alkaloid content in true salamanders of the genus Salamandra (Salamandridae). Mol. Phylogenet. Evol. 73: 208–216, https://doi.org/10.1016/j.ympev.2013.12.009.Search in Google Scholar
Von Byern, J., Farkaš, R., Steinort, D., Greistorfer, S., Eckhard, M., and Cyran, N. (2021). Perspective for a new bioinspired permanent adhesive for dry conditions - insights in the glue producing Japanese art of defence system of the oita salamander Hynobius dunni. Front. Mech. Eng. 7: 667857, https://doi.org/10.3389/fmech.2021.667857.Search in Google Scholar
Von Byern, J., Grunwald, I., Koskok, M., Saporito, R.A., Dicke, U., Wetjen, O., Thiel, K., Borcherding, K., Kowalik, T., and Marchetti-Deschmann, M. (2017). Chemical characterization of the adhesive secretions of the salamander Plethodon shermani (Caudata, Plethodontidae). Sci. Rep. 7: 6647, https://doi.org/10.1038/s41598-017-05473-z.Search in Google Scholar
Wang, Y., Guo, L., Liu, J., Huang, X., Wang, X., Guo, X., You, X., Li, W., Sun, W., Sun, T., et al.. (2022). Chitosan/PLGA shell nanoparticles as tylotoin delivery platform for advanced wound healing. Int. J. Biol. Macromol. 220: 395–405, https://doi.org/10.1016/j.ijbiomac.2022.07.244.Search in Google Scholar
Woodley, S.K. and Staub, N.L. (2021). Pheromonal communication in urodelan amphibians. Cell Tissue Res. 383: 327–345, https://doi.org/10.1007/s00441-020-03408-1.Search in Google Scholar
Xiao, Y., Liu, C., and Lai, R. (2011). Antimicrobial peptides from amphibians. BioMol. Concepts 2: 27–38, https://doi.org/10.1515/bmc.2011.006.Search in Google Scholar
Xu, X. and Lai, R. (2015). The chemistry and biological activities of peptides from amphibian skin secretions. Chem. Rev. 115: 1760–1846, https://doi.org/10.1021/cr4006704.Search in Google Scholar
Yamamoto, K., Kawai, Y., Hayashi, T., Ohe, Y., Hayashi, H., Toyoda, F., Kawahara, G., Iwata, T., and Kikuyama, S. (2000). Silefrin, a sodefrin-like pheromone in the abdominal gland of the sword-tailed newt. Cynops ensicauda. FEBS Lett. 472: 267–270, https://doi.org/10.1016/s0014-5793(00)01455-1.Search in Google Scholar
Yang, H., Liu, R., Cui, D., Liu, H., Xiong, D., Liu, X., and Wang, L. (2017a). Analysis on the expression and function of a chicken-type and goose-type lysozymes in Chinese giant salamanders Andrias davidianus. Dev. Comp. Immunol. 72: 69–78, https://doi.org/10.1016/j.dci.2017.02.017.Search in Google Scholar
Yang, H., Lu, B., Zhou, D., Zhao, L., Song, W., and Wang, L. (2017b). Identification of the first cathelicidin gene from skin of Chinese giant salamanders Andrias davidianus with its potent antimicrobial activity. Dev. Comp. Immunol. 77: 141–149, https://doi.org/10.1016/j.dci.2017.08.002.Search in Google Scholar
Yang, J. and Zhang, Y. (2015). I-TASSER server: new development for protein structure and function predictions. Nucleic Acids Res. 43: W174–W181, https://doi.org/10.1093/nar/gkv342.Search in Google Scholar
Yotsu, M., Iorizzi, M., and Yasumoto, T. (1990). Distribution of tetrodotoxin, 6-epitetrodotoxin, and 11-deoxytetrodotoxin in newts. Toxicon 28: 238–241, https://doi.org/10.1016/0041-0101(90)90419-8.Search in Google Scholar
Yotsu-Yamashita, M., Mebs, D., Kwet, A., and Schneider, M. (2007). Tetrodotoxin and its analogue 6-epitetrodotoxin in newts (Triturus spp.; Urodela, Salamandridae) from southern Germany. Toxicon 50: 306–309, https://doi.org/10.1016/j.toxicon.2007.03.019.Search in Google Scholar
Yotsu-Yamashita, M., Toennes, S.W., and Mebs, D. (2017). Tetrodotoxin in Asian newts (Salamandridae). Toxicon 134: 14–17, https://doi.org/10.1016/j.toxicon.2017.05.014.Search in Google Scholar
Yu, H., Gao, J., Lu, Y., Guang, H., Cai, S., Zhang, S., and Wang, Y. (2013). Molecular cloning, sequence analysis and phylogeny of first Caudata G-type lysozyme in axolotl (Ambystoma mexicanum). Zool. Sci. 30: 938–943, https://doi.org/10.2108/zsj.30.938.Search in Google Scholar
Zalesky, S. (1866). Über das Samandarin. Das Gift der Salamandra maculata. Med chem. Untersuch. Hoppe-Seyler 1: 85–116.Search in Google Scholar
© 2024 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- Highlight: GBM Young Investigators Part 5
- Highlight: young research groups in Germany – 5th edition
- Protein persulfidation in plants: mechanisms and functions beyond a simple stress response
- Pathological and physiological roles of ADP-ribosylation: established functions and new insights
- Implications of TRPM3 and TRPM8 for sensory neuron sensitisation
- The complex regulation of Slo1 potassium channels from a structural perspective
- The TOM complex from an evolutionary perspective and the functions of TOMM70
- Insights in caveolae protein structure arrangements and their local lipid environment
- Insights into caudate amphibian skin secretions with a focus on the chemistry and bioactivity of derived peptides
- Analysis of cell cycle stage, replicated DNA, and chromatin-associated proteins using high-throughput flow cytometry
- A tailored cytochrome P450 monooxygenase from Gordonia rubripertincta CWB2 for selective aliphatic monooxygenation
Articles in the same Issue
- Frontmatter
- Highlight: GBM Young Investigators Part 5
- Highlight: young research groups in Germany – 5th edition
- Protein persulfidation in plants: mechanisms and functions beyond a simple stress response
- Pathological and physiological roles of ADP-ribosylation: established functions and new insights
- Implications of TRPM3 and TRPM8 for sensory neuron sensitisation
- The complex regulation of Slo1 potassium channels from a structural perspective
- The TOM complex from an evolutionary perspective and the functions of TOMM70
- Insights in caveolae protein structure arrangements and their local lipid environment
- Insights into caudate amphibian skin secretions with a focus on the chemistry and bioactivity of derived peptides
- Analysis of cell cycle stage, replicated DNA, and chromatin-associated proteins using high-throughput flow cytometry
- A tailored cytochrome P450 monooxygenase from Gordonia rubripertincta CWB2 for selective aliphatic monooxygenation