Anti-inflammatory mediators for molecular imaging of atherosclerosis
-
Gunter Almer
Gunter Almer studied Microbiology at the Karl-Franzens University of Graz which he finished with a Master’s thesis in Phospholipid Biology in 2006. From 2004 to 2006 he was a scientific co-worker for a clinical study focusing on juvenile obesity at the Clinical Institute for Medical and Chemical Laboratory Diagnosis, Medical University of Graz and later on, until 2007, also at the Center for Medical Research (ZMF) in Graz. In 2007 he started a PhD thesis in Preclinical Atherosclerotic (AS-)imaging using biomarkers and nanotechnology at the ZMF, which he finished in 2011. From 2011 to 2012 he was a scientific co-worker at the IBN, Austrian Academy of Science, focusing on AS-imaging with liposomal nanoparticles and on imaging brown and white adipose tissue. Since 2013 he works as PostDoc for the EU project NanoAthero continuing focusing on the tasks AS-imaging in animal models of atherosclerosis with various nanoparticles.
Peter Opriessnig is currently working as PhD student at the Institute of Medical Engineering, Graz University of Technology (Austria). The topic of his PhD thesis and his research interest focuses on the preclinical evaluation of atherosclerotic lesions at the molecular level using contrast enhanced magnetic resonance imaging (CE-MRI).
Ruth Prassl received her PhD degree in Chemistry and Physics at the Karl-Franzens University of Graz in 1986. Since 2012 she holds a position as Associate Professor at the Institute of Biophysics of the Medical University of Graz. Previously she was leader of the working group “Lipoproteins and Nanomedicine” at the Institute of Biophysics and Nanosystems Research of the Austrian Academy of Sciences. Her research interests are in molecular biophysics with special focus on structure analysis of lipoproteins and membrane associated proteins. A second major thrust of her research addresses the design and development of self-assembling nanostructures including liposomal nanocarriers for diagnosis and therapy.
Harald Mangge is a Medical Doctor and Professor at the Department of Laboratory Medicine at the Medical University of Graz, Austria. He conducts the research unit on lifestyle and inflammation-associated biomarkers. His research focuses on immune-mediated inflammation in cardiovascular and metabolic diseases. Another focus is nanomedicine, where an improved diagnosis and treatment of atherosclerosis is investigated (
http://www.nanoathero.eu/ ). Harald Mangge conducts the STYJOBS/EDECTA cohort project, a large prospective, observational study to improve the understanding of metabolic and cardiovascular risk in obesity (http://clinicaltrials.gov/ct2/show/NCT00482924 ). Further, Harald Mangge holds the function of a Vicespeaker of the Cardiovascular Research Field of the Medical University of Graz, and he is the MUG managing director of the BioTechMed initiative, an interdisciplinary strategic joint project of the three large universities at the location of Graz (http://biotechmedgraz.at/de/ ). With respect to his research activities in cardiovascular diseases Harald Mangge holds the function of a Vicespeaker of the Cardiovascular Research Field of the Medical University of Graz. Further, he is MUG Coordinator of the BioTechMed Initiative a large strategic joint project of the three large universities at the location of Graz (Medical-/Karl Franzens-/Technical-University).

Abstract
Nanomedicine, a young and innovative field, offers interesting approaches for diagnosis and treatment in personalized medicine. Myocardial infarction and stroke belong to the most important challenges in this context because an improved early diagnosis of individuals well before fatal clinical endpoints occur is urgently needed. The underlying cause of myocardial infarction and stroke is atherosclerosis, a chronic immune-mediated inflammation of the vascular wall involving monocytes, macrophages, T-lymphocytes, and arterial wall cells. Hence, an immense number of pro-inflammatory mediators have been investigated in the context of nanomedicine and atherosclerosis but, interestingly, only few anti-inflammatory biomarkers. Nevertheless, the anti-inflammatory axis is always present as a negative feedback if a critical inflammatory perpetuation destabilizes atherosclerotic lesions. Hence, we could show that the immune-modulating, anti-inflammatory molecules, adiponectin and interleukin-10, are useful for molecular imaging of AS plaques. Based on recent publications in animal models of atherosclerosis, we strongly assume that the inflammatory “brake” mechanisms may represent an interesting new tool to specifically target the scenario of culprit AS-lesions. In this review article we discuss the potential of adiponectin, interleukin-10 and other anti-inflammatory active molecules like targeted liposomes and high dense lipoproteins towards this.
Introduction
Despite considerable therapeutic advances over the past 50 years, cardiovascular events are the leading causes of death worldwide. This is primarily due to the increasing prevalence of atherosclerosis (AS) which is associated with a sedentary and obesogenic life style increasingly present in the so-called Western, industrialized world. Atherosclerosis is a sub-acute inflammatory condition of the vascular wall, characterized by the infiltration of macrophages and T-cells which interact with one another and with the arterial wall cells (1). The chronic inflammatory process leads to the formation, progression and rupture of vascular lesions, called AS-plaques (2).
The early identification and the ability to follow the development of AS-plaques are still unsolved challenges for medical imaging, which is limited not only by the performance of present imaging techniques, but also by the availability of specific molecules for targeted recognition (3). Unfortunately, so far AS is only diagnosed at the advanced stages of the disease: either by directly measuring the degree of stenosis, or by evaluating the effect of arterial stenosis on organ perfusion (4). Nevertheless, over the past few years, advances have been made in imaging techniques that enable the visualization and monitoring of AS-lesions’ progression or regression (5). However, a reliable, non-invasive technique to detect different stages of AS for an applicable, clinical characterization of AS-plaques is still missing (6). With time, an immense number of mediators involved in the inflammatory scenario of AS progression have been suggested, and tested to recognize AS-lesions (7, 8). Interestingly, the great majority of all studies focused on pro-inflammatory molecules, practically none on anti-inflammatory ones (3).
To close this gap, we review herein the potential of anti-inflammatory candidate molecules for an improved molecular imaging of AS lesions. The majority of the data is based on recent observations in animal models of AS by which anti-inflammatory molecules uncoupled, or coupled with nanoparticles (NPs) have turned out to be promising for an improved diagnosis of vulnerable AS lesions (9–12). Vulnerable AS plaques are characterized by a thin fibrous cap, a large, necrotic lipid core, an increased number of activated macrophages, and less collagen and smooth muscle cells content (13). Refering to the utmost importance of inflammatory activation for the generation of vulnerability, the diagnostic use of anti-inflammatory molecules implicates also potential theranostic effects.
Liposomes – “nanofat” carriers for anti-inflammatory action
Nanoparticles offer new possibilities as homing devices for various drugs or contrast agents. Although cancer therapy and cancer diagnostics are still dominating the field, the detection of AS plaques is an equally attractive endeavor in the prevention of cardiovascular diseases (CVD) (4, 7, 14). Liposomes belong to the most widely studied class of NPs in use. They are composed of biologically degradable phospholipid molecules and in aqueous solution they typically assemble to closed bilayer structures (15). They are easy to handle and offer a unique variety of possible combinations concerning their overall size and charge. The surface of liposomes is often coated with polyethylene glycol (PEG-) molecules to yield “stealth” particles which can elude the reticulo-endothelial system (RES), and thus, can have prolonged circulation times ranging from hours to days (16). Such liposomes are easily modifiable with signal-emitting groups to be detected by different imaging modalities. For magnetic resonance imaging (MRI) iron oxide nanoparticles or lipid conjugated gadolinium (Gd3+) ion chelates are widely used. Gadolinium-based products, however, should be used with caution, since free Gd3+ is highly toxic and intracellular gadolinium is known to induce cell apoptosis. Especially in patients with renal insufficiency the risk for inducing nephrogenic systemic fibrosis is significantly increased after exposure to gadolinium based contrast agents (17). Using liposomes for MRI requires a balance between blood circulation time, time required for accumulation and retention of nanoparticles in the plaque and blood clearance rates. The latter is important to reduce background signal for imaging (18). As gadolinium complexes have relaxivities between 3 and 15 mM–1s–1 at 3 Tesla and a tissue relaxation rate of 1 s–1 a very high concentration of about 50 nmol Gd3+ per gram of tissue is required for detectable signal alterations. In contrast, iron oxide nanoparticle loaded liposomes produce local magnetic susceptibility, which results in a distance dependent dephasing of hydrogen nuclei. Such superparamagnetic liposomal contrast agents have a thousand times higher magnetic moments compared to paramagnetic gadolinium containing ones, therefore improving sensitivity tremendously (19). For nuclear medicine, liposomes are radiolabeled with 64Cu, 68Ga or 18F for positron emission tomography (PET), while 67Ga and 111In are frequently used radiolabels for single photon emission computed tomography (SPECT) (19).
Besides incorporation or labeling with diagnostic markers, functionalized groups can be attached to the distal end of the PEG-chains for coupling of molecules enabling recognition of specific biologic targets (20–22). The targeting moiety, such as an antibody, biomarker or specific ligand must ideally have a high affinity for the desired target molecule. Targeted probes tested for cardiovascular imaging typically consist of a targeting sequence linked to the NP which is fused to a contrast agent appropriate to the particular imaging technique applied (23). By choosing the proper ligand, liposomes can be specifically directed to inflammatory sites, for instance to monocyte derived inflammatory macrophages in advanced AS lesions.
Alternatively, stealth liposomes can be loaded with therapeutic substances with anti-inflammatory properties. In this context, a very promising setup was recently published by Lobatto et al. (24). First, they induced AS in a rabbit model by balloon angioplasty of the aorta. Then, they prepared long-circulating anti-inflammatory PEGylated liposomes by encapsulating the corticosteriod prednisolone in the aqueous interior of the liposomes. Large scale good manufacturing practice (GMP) production was achieved by using high-shear homogenization for liposome preparation instead of extrusion membranes. Shelf life stability studies with the liposomal corticosteroid product indicated that it remains stable for at least 2 years when stored between 2 and 8°C. The liposomes were injected into the animals, and a 18F-fluorodeoxyglucose positron emission tomography/computed tomography (FDG-PET/CT) showed an anti-inflammatory effect which persisted over one week after a single dose. The positive effect was also proved by MR imaging compared to no changes with the free corticosteroid (24). Thus, liposomes can be used to achieve strong anti-inflammatory effects – a strategy promising for the future treatment of AS in humans.
Disadvantages of liposomes for in vivo studies are that their performance strongly depends on the formulation, the targeting sequence, the applied imaging technique, and unfortunately also on the animal models, creating atherosclerotic lesions different to those in humans. This is one of the major limitations that hamper the translation from promising pre-clinical findings with diagnostic liposomes in animal models of atherosclerosis to clinical studies (25). Hence, no liposomal imaging agent for targeted imaging of AS is approved or in clinical trials. However, the capability to include high payloads of contrast generating materials into liposomes makes them highly attractive for diagnosis. Especially, ligand targeted liposomes show great promise to identify different components of the plaque scenario with high specificity. This can be especially valuable in the early phase of clinical trials to detect inflammatory changes before morphological changes occur (25, 26).
HDL – native anti-inflammatory nanoparticles
High density lipoprotein (HDL) particles are the smallest and most dense lipoprotein entities with diameters in a range from 8 to 10 nm. They contain paraoxonase, a potent antioxidant enzyme, and are rich on different amounts of exchangeable apolipoprotein species, of which the most prominent candidate is apolipoprotein AI (apo-AI) (27). Physiologically, HDL plays a significant role in reverse cholesterol transport and mediates antioxidant effects as well as anti-inflammatory and antithrombotic functions. In contrast to low density lipoprotein (LDL), where the entire particle is taken up, only the core lipids of HDL are shuttled into the cells. This uptake process is referred to as selective cholesterol uptake mediated by the scavenger receptor class B type I (SR-BI) (28). Cholesterol is off-loaded and either converted into a bile salt by hepatocytes or is directly excreted into the bile. A significant fraction of HDL cholesterol is also delivered to the liver by way of the LDL-receptor, after first being transferred from HDL to LDL by cholesterol ester transfer protein (CETP). As an inverse correlation exists between blood HDL levels and the risk to develop CVD, elevated serum concentrations of HDL are regarded as advantageous (29, 30).
Hence, the use of artificial HDL as a transport vehicle may be attractive for therapeutic and diagnostic applications. More recent papers underline this potential of HDL like NPs, so far most promising in cancer research (31). The small size of artificial HDL (<30 nm) allows it to maneuver deeply into target organs, e.g., tumors (32), a characteristic which is probably also useful for the management of AS lesions.
Reconstituted HDL
Besides the application in cancer, studies concentrate on the intrinsic anti-atherogenic and cardioprotective properties of HDL, and deal with the use of reconstituted (r)HDL as therapeutic agent (33). Repeated treatment with rHDL was shown to reduce inflammatory and atherogenic processes in different animal studies. Such preclinical studies have been refined to human clinical trials revealing that rHDL is well tolerated by patients. It was found that intravenous injections of rHDL significantly raises the plasma levels of HDL (34). Recently, Ibanez et al. (35) compared the anti-inflammatory and plaque stabilizing properties of wild type HDL and rHDL. They found that the infusion of rHDL(Milano) exerts superior positive effects than wild type HDL (35). rHDL can further be applied for the targeted delivery of lipophilic and poorly water soluble drugs, as well as for cholesterol modified siRNA (36–38). Modified rHDL has also been studied as contrast agent for AS imaging. Macrophage infiltration is positively correlated with lesion progression, plaque size and intimal thickness (39). Hence, HDL particles penetrate the plaque due to their small size and target macrophages. Based on this, HDL seems to be excellently suited for the recognition of atherosclerotic plaques. Regarding antioxidant and anti-inflammatory properties, HDL particles may also mediate disease-stabilizing effects. In a very recent paper Duivenvoorden et al. have developed statin-loaded rHDL NPs, which were identified as potent modulators of plaque inflammation. The statin-loaded rHDLs accumulated in atherosclerotic plaques where they were taken up by macrophages. For a long-term, low dose treatment with statin-loaded rHDLs, an inhibition of plaque inflammation progression was achieved, while a short-term, high dose treatment was able to markedly decrease inflammation in advanced atherosclerotic plaques (40).
Synthetic HDL
Over the last decade, the method for lipoprotein inspired imaging of AS by producing fully synthetic HDL-particles containing a spherical nanocrystalline core was established (41, 42). In synthetic HDLs the inner lipophilic core is surrounded by a monolayer of phospholipids with apo-AI molecules associated to the surface. The nanocrystalline core can be composed of either Gold-nanoparticles for computed tomography (CT) imaging, quantum dots (QD) for optical imaging (41) or ironoxide-nanoparticles for magnet resonance imaging (MRI) (42). Additionally, fluorescent dyes or phospholipid anchored Gadolinium (Gd)-chelates can be incorporated into the outer phospholipid monolayer of synthetic HDLs to construct a multimodal signal-emitting particle for molecular imaging. The synthetic HDL particles are spherical and highly homogeneous, having a diameter and lipid to protein content similar to natural HDL. In vivo experiments in the atherosclerotic apoE deficient mouse model revealed an accumulation of nanocrystalline HDL particles in the vessel wall and macrophages of atherosclerotic lesions (41).
In summary, artificial HDL assemblies may represent a flexible nanoplatform with future potential for molecular diagnostics and therapeutics, concurrently possessing intrinsic antiatherosclerotic and anti-inflammatory activities. Nevertheless, there are also substantial obstacles. HDL is strongly internalized into cells of the liver. If labeled with Gd the recirculation of GdrHDL particles will make nuclear imaging impossible due to the ever present background signal relative to the half-life of the isotope. Additionally, studies using GdDTPA-lipids show that the Gd label is liable and can transfer to other lipoproteins in the vascular phase (43). This fact limits clinical translation of the rHDL platform for MR diagnostic imaging. The use of iron oxides may be an alternative but changes in the charge and size of the synthetic HDL following incorporation of iron may again cause significant problems.
Interleukin-10 – the anti-inflammatory “master” cytokine
A broad spectrum of proinflammatory cytokines has been reported to be involved and to stimulate the progression of AS (11, 44–52), whereas, more recently, few were found to potentially aid in AS regression (53–55). Not the least, this may be caused by the fact that the investigation of the role of regulatory T cells (Tregs) dominated the discussion how to achieve a sufficient atheroprotective immune response (53, 56). Apart from this, Pinderski Oslund et al. found that activated T-lymphocytes overexpress the anti-inflammatory cytokine interleukin-10 (IL-10), and that this event is capable of blocking AS actions in vitro and in vivo (57, 58). Xie et al. showed in a cross-sectional study that decreased serum IL-10 concentrations were significantly associated with an increased likelihood of ischemic stroke (59). Further, IL-10 expression was found elevated in advanced and unstable AS-plaques (60), and it appears to contribute to the regulation of the local inflammatory response, and is involved in the control of cell death within plaques (60–63). This is underlined by the recent observation where IL-10 mediated the immunoregulatory response in conjugated linoleic acid-induced regression of AS (64). One of the most important facet of this immunological “brake” is represented by inhibition of pro-inflammatory cytokines and mediators from macrophages and dendritic cells (65–67). Hence, IL-10 is believed to be the most prominent anti-inflammatory cytokine. Albeit, this cytokine is neglected in the recent AS research.
Many cells produce IL-10. Major sources are T-helper cells (68) like CD8 positive T-cells (69), monocytes, macrophages (65), subsets of dendritic cells (66), human B lymphocytes (70) and mast cells (71). The spectrum of non-immune cells comprises keratinocytes, epithelial cells, and tumor cells (72, 73). IL-10 signals mainly through a two-receptor complex: IL-10 receptor 1 (IL-10R1) and IL-10 receptor 2 (IL-10R2) (74). Nevertheless, the IL-10-binding receptor complex is one of only a few regulating factors known so far.
As summarized, many characteristics suggest IL-10 as an interesting biomarker for AS. Hence, we investigated the potency of IL-10 as targeting molecule for molecular imaging of AS plaques. This was based on the assumption that where inflammatory fire is strongly present (in vulnerable AS lesions) there is increased need for “fire water” – i.e., an upregulated anti-inflammatory cascade most likely mediated by the anti-inflammatory “master” cytokine IL-10. Indeed, we could show that recombinant IL-10 preferentially accumulates in AS-plaque areas. Nanoconstructs of IL-10 and PEGylated liposomes (Figure 1) increased IL-10’s stability in vivo and the specificity of target recognition (12). Notably, no significant immune reactions were observed so far when the nanoconstruct was injected into wild type mice (12). To conclude, IL-10 combined with multifunctionalized liposomes is a promising candidate for multi-modal AS-plaque imaging (12). Further investigations are needed to clarify if this holds also true in the human system. Potential drawbacks may be given by the fact that IL-10, as an abundant molecule, may reduce the sensitivity of the imaging modality. This should be clarified in future investigations.
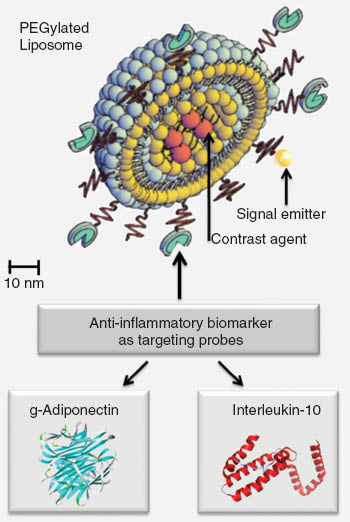
Shows a nanoconstruct consisting of a PEGylated stealth liposome and antiinflammatory biomarkers (i.e., globular adiponectin, interleukin-10) attached for specific targeting.
Adiponectin – an anti-inflammatory molecule of unconventional origin
It is well established now that adipose tissue is a highly active endocrine organ, also centrally involved in the regulation of inflammation (75–77). The most important anti-inflammatory adipokine is adiponectin (78), predominantly synthesized by adipocytes (79). The basic structure of the full-length form of adiponectin (fAd) is a 247 amino acid protein with four domains (80). In the bloodstream, adiponectin circulates at concentrations of 2–30 μg/mL (81) and in different molecular weight forms; high molecular weight (oligomer); medium molecular weight (hexamer); and low molecular weight (trimer) adiponectin (82, 83). These subfractions exert different biological functions (82–84). Adiponectin is thought to protect against diabetes and AS (11, 82, 83, 85–88). Li et al. showed that local treatment with adiponectin reduces atherosclerotic plaque size in rabbits by decreasing the expression of vascular cell adhesion molecule-1 (VCAM-1) and intercellular adhesion molecule-1 (ICAM-1) in the vascular walls (89). Recently, Okamoto et al. found that adiponectin reduces the thrombogenic potential of macrophages by inhibiting tissue factor expression and activity (90). Hypoadiponectinemia was found to be a significant predictor of endothelial dysfunction in both peripheral and coronary arteries (91, 92), and associated with preatherosclerotic symptoms of obese juveniles (83, 87). Adiponectin was also shown to protect endothelial progenitor cells from apoptosis (93). In subjects with metabolic syndrome, a significant negative correlation was observed between serum adiponectin concentrations, plaque volume, and highly sensitive C-reactive protein levels (94).
Moreover, Waki et al. showed that leukocyte elastase secreted from activated monocytes and/or neutrophils can cleave fAd (95). This cleavage might be the reason for the generation of the 17 kDa globular fragment of adiponectin (gAd), which is found at lower levels (about 1% of total adiponectin) in the circulation (96). Globular Adiponectin was shown to increase insulin-stimulated glucose uptake and to boost β-oxidation of fatty acids (97, 98), while other functions remain controversial (99, 100). Apart from the anti-atherosclerotic and anti-diabetic properties, adiponectin has potent anti-inflammatory effects (101). The latter are associated with the ability to stimulate the production of nitric oxide (NO)(102) which protects endothelial cells (103, 104). Endothelial damage plays a crucial role in the formation of AS leading to adhesion and penetration of monocytes followed by macrophage-to-foam cell transformation (105). Adya et al. found that gAd decreased glucose and C-reactive protein-induced angiogenesis with a concomitant reduction in matrixmetalloproteinase-2 (MMP-2), MMP-9, and vascular endothelial growth factor in human microvascular endothelial cells (106). Thus, gAd may support the regeneration of AS damaged vascular tissue. Two main receptors are known to bind adiponectin, namely adipoR1 and adipoR2 (107). AdipoR1 is expressed mainly on vascular smooth muscle cells and endothelial cells, predominantly binding gAd. AdipoR2 binds both gAd and fAd and is highly expressed on hepatocytes, and in smaller amounts in the hypothalamus and on brain endothelial cells (108). Further, fAd was shown to bind to T-cadherin (109) and to the multifunctional protein, calreticulin (110). With respect to AS, adiponectin has been shown to accumulate in the subendothelial space of vascular walls after injury of the endothelial barrier where it binds to different types of collagen of the vascular intima (111). In contrast, adiponectin was not detected in the subendothelial space of human atherosclerotic lesions covered by an intact endothelium (112).
We investigated i) the role of adiponectin and subfractions in obesity associated preatherosclerosis of humans (82, 83, 87, 88, 113), and ii) the potential of fluorescence-labeled gAd and fAd subfractions (fAd-Sfs) to bind to atherosclerotic lesions in apoE-deficient mice (9–11). By the animal studies, we found a low binding efficiency of fAd but a, probable inflammation-mediated, strong accumulation of gAd in the fibrous cap of atherosclerotic plaques (9, 10). Thus, gAd may be an appropriate targeting sequence for the molecular imaging of AS lesions (9). We developed also nanoconstructs between gAd and PEGylated stealth liposomes (10) (Figure 1). By means of the NPs, a higher payload of signal emitting molecules could be transported to the atherosclerotic plaques (10). Moreover, we investigated constructs between gAd and protamine-oligonucleotide NPs, called proticles. Proticles are biodegradable NPs, which have been previously developed for the delivery of various active compounds like antisense oligonucleotides or small peptides (114–117). They show a certain affinity to monocytes and macrophages which may be of additional interest for sequential AS plaque targeting. Limitations may be given by clinical unfavorable effects of gAd alone or in combination with NPs. The fact that Ad is an abundant molecule of the physiologic scenario argues against this. Nevertheless, this physiologic abundance can also reduce the sensitivity of the imaging modality. Further investigations are underway by our group to clarify this.
Taken together, our results indicate an interesting potential of gAd-targeted NPs for the imaging of atherosclerotic vascular lesions. Combinations of targeted NPs may be useful for the discrimination between stable and vulnerable plaques (10).
Conclusion
In the foregoing we have summarized evidence that anti-inflammatory mediators constitute a new innovative scope of application for diagnosis and therapeutic modification of atherosclerotic lesions. Early diagnosis and identification of individuals at risk by molecular imaging well before fatal clinical endpoints occur is warranted for an improved personalized medicine of AS. Interestingly, to the best of our knowledge, only few anti-inflammatory biomarkers have been investigated in the context of nanomedicine and AS. This is astonishing because anti-inflammatory activation is always present as a negative feedback mechanism, if inflammatory perpetuation of AS lesions towards clinical endpoints occurs. Thus, this inflammatory “brake” may probably more specifically target the scenario of vulnerable, culprit AS-lesions than the more unspecific systemic and local inflammation. Novel molecules with the ability to balance the inflammatory force, and safe as well as specific delivery are warranted. The implementation of multi-functional probes (theranostic particles for concomitant imaging and therapy) combined with the advantages of differential imaging (e.g., high spatial resolution, soft tissue contrast MRI, high sensitivity PET) will improve the diagnostic performance. The immune-modulating, anti-inflammatory characteristics of adiponectin and IL-10 are useful in this context, and will be further investigated by our group for their usability in humans.
Thus far, there is no robust blood test ready for clinical use to identify patients with active perpetuation of AS towards vulnerablility.
In addition to the Framingham risk score, a gender- specific algorithm used to estimate the 10-year cardiovascular risk of an individual, we must find distinct cardiovascular phenotypes and biomarker signatures that complement the clinical decision-making for an early preventive treatment of AS. It must be noted that relevant studies so far largely dealt with single adipokines, cytokines, chemokines and their receptors measured in limited numbers of patients. Proteomics for improved diagnostics can extend the portfolio of techniques and introduce new avenues. For example, potentially “druggable” microRNAs are of high interest (118, 119). Further, the relationships between oxidation-specific epitopes (OSE), apolipoprotein-A (apoA), progredient AS, and plaque vulnerability may be promising (120). Human atherosclerotic lesions showed a differential expression of OSEs and apoA as they progress, rupture, and become clinically symptomatic. These findings may provide a rationale for targeting OSE for biotheranostic applications in humans (120).
Taken together, the recognition of individual inflammatory/metabolic signatures by approaches of personalized medicine may give important and new insights towards early diagnosis and monitoring of AS, and may finally provide the basis of new therapeutic strategies. Anti-inflammatory feedback cascades will be an interesting part for completion of this mission.
About the authors

Gunter Almer studied Microbiology at the Karl-Franzens University of Graz which he finished with a Master’s thesis in Phospholipid Biology in 2006. From 2004 to 2006 he was a scientific co-worker for a clinical study focusing on juvenile obesity at the Clinical Institute for Medical and Chemical Laboratory Diagnosis, Medical University of Graz and later on, until 2007, also at the Center for Medical Research (ZMF) in Graz. In 2007 he started a PhD thesis in Preclinical Atherosclerotic (AS-)imaging using biomarkers and nanotechnology at the ZMF, which he finished in 2011. From 2011 to 2012 he was a scientific co-worker at the IBN, Austrian Academy of Science, focusing on AS-imaging with liposomal nanoparticles and on imaging brown and white adipose tissue. Since 2013 he works as PostDoc for the EU project NanoAthero continuing focusing on the tasks AS-imaging in animal models of atherosclerosis with various nanoparticles.

Peter Opriessnig is currently working as PhD student at the Institute of Medical Engineering, Graz University of Technology (Austria). The topic of his PhD thesis and his research interest focuses on the preclinical evaluation of atherosclerotic lesions at the molecular level using contrast enhanced magnetic resonance imaging (CE-MRI).

Ruth Prassl received her PhD degree in Chemistry and Physics at the Karl-Franzens University of Graz in 1986. Since 2012 she holds a position as Associate Professor at the Institute of Biophysics of the Medical University of Graz. Previously she was leader of the working group “Lipoproteins and Nanomedicine” at the Institute of Biophysics and Nanosystems Research of the Austrian Academy of Sciences. Her research interests are in molecular biophysics with special focus on structure analysis of lipoproteins and membrane associated proteins. A second major thrust of her research addresses the design and development of self-assembling nanostructures including liposomal nanocarriers for diagnosis and therapy.

Harald Mangge is a Medical Doctor and Professor at the Department of Laboratory Medicine at the Medical University of Graz, Austria. He conducts the research unit on lifestyle and inflammation-associated biomarkers. His research focuses on immune-mediated inflammation in cardiovascular and metabolic diseases. Another focus is nanomedicine, where an improved diagnosis and treatment of atherosclerosis is investigated (http://www.nanoathero.eu/). Harald Mangge conducts the STYJOBS/EDECTA cohort project, a large prospective, observational study to improve the understanding of metabolic and cardiovascular risk in obesity (http://clinicaltrials.gov/ct2/show/NCT00482924). Further, Harald Mangge holds the function of a Vicespeaker of the Cardiovascular Research Field of the Medical University of Graz, and he is the MUG managing director of the BioTechMed initiative, an interdisciplinary strategic joint project of the three large universities at the location of Graz (http://biotechmedgraz.at/de/). With respect to his research activities in cardiovascular diseases Harald Mangge holds the function of a Vicespeaker of the Cardiovascular Research Field of the Medical University of Graz. Further, he is MUG Coordinator of the BioTechMed Initiative a large strategic joint project of the three large universities at the location of Graz (Medical-/Karl Franzens-/Technical-University).
Acknowledgments
This work is supported by funding under the European FP7 program “NanoAthero”-NMP4-LA-2012-3099820.
References
1. Rocha VZ, Libby P. Obesity, inflammation, and atherosclerosis. Nat Rev Cardiol 2009;6:399–409.10.1038/nrcardio.2009.55Search in Google Scholar PubMed
2. Libby P, Okamoto Y, Rocha VZ, Folco E. Inflammation in atherosclerosis: transition from theory to practice. Circ J 2010;74:213–20.10.1253/circj.CJ-09-0706Search in Google Scholar
3. Libby P, Nahrendorf M, Weissleder R. Molecular imaging of atherosclerosis: a progress report. Tex Heart Inst J 2010;37:324–7.Search in Google Scholar
4. Sanz J, Fayad ZA. Imaging of atherosclerotic cardiovascular disease. Nature 2008;451:953–7.10.1038/nature06803Search in Google Scholar PubMed
5. Helft G, Worthley SG, Fuster V, Fayad ZA, Zaman AG, Corti R, et al. Progression and regression of atherosclerotic lesions: monitoring with serial noninvasive magnetic resonance imaging. Circulation 2002;105:993–8.10.1161/hc0802.104325Search in Google Scholar PubMed
6. Camici PG, Rimoldi OE, Gaemperli O, Libby P. Non-invasive anatomic and functional imaging of vascular inflammation and unstable plaque. Eur Heart J 2012;33:1309–17.10.1093/eurheartj/ehs067Search in Google Scholar PubMed
7. Nahrendorf M, Sosnovik DE, Weissleder R. MR-optical imaging of cardiovascular molecular targets. Basic Res Cardiol 2008;103:87–94.10.1007/s00395-008-0707-2Search in Google Scholar PubMed PubMed Central
8. Choudhury RP, Fisher EA. Molecular imaging in atherosclerosis, thrombosis, and vascular inflammation. Arterioscler Thromb Vasc Biol 2009;29:983–91.10.1161/ATVBAHA.108.165498Search in Google Scholar PubMed PubMed Central
9. Almer G, Saba-Lepek M, Haj-Yahya S, Rohde E, Strunk D, Frohlich E, et al. Globular domain of adiponectin: promising target molecule for detection of atherosclerotic lesions. Biologics: Targ Ther 2011;5:95–105.Search in Google Scholar
10. Almer G, Wernig K, Saba-Lepek M, Haj-Yahya S, Rattenberger J, Wagner J, et al. Adiponectin-coated nanoparticles for enhanced imaging of atherosclerotic plaques. Int J Nanomed 2011;6:1279–90.Search in Google Scholar
11. Mangge H, Almer G, Truschnig-Wilders M, Schmidt A, Gasser R, Fuchs D. Inflammation, adiponectin, obesity and cardiovascular risk. Current medicinal chemistry. 2010;17:4511–20.10.2174/092986710794183006Search in Google Scholar PubMed
12. Almer G, Frascione D, Pali-Scholl I, Vonach C, Lukschal A, Stremnitzer C, et al. Interleukin-10: an anti-inflammatory marker to target atherosclerotic lesions via PEGylated liposomes. Mole Pharm 2013;10:175–86.10.1021/mp300316nSearch in Google Scholar PubMed PubMed Central
13. Salagianni M, Galani IE, Lundberg AM, Davos CH, Varela A, Gavriil A, et al. Toll-like receptor 7 protects from atherosclerosis by constraining “inflammatory” macrophage activation. Circulation 2012;126:952–62.10.1161/CIRCULATIONAHA.111.067678Search in Google Scholar PubMed
14. Lindsay AC, Choudhury RP. Form to function: current and future roles for atherosclerosis imaging in drug development. Nat Rev Drug Discov 2008;7:517–29.10.1038/nrd2588Search in Google Scholar
15. Sessa G, Weissmann G. Phospholipid spherules (liposomes) as a model for biological membranes. J Lipid Res 1968;9:310–8.10.1016/S0022-2275(20)43097-4Search in Google Scholar
16. Immordino ML, Dosio F, Cattel L. Stealth liposomes: review of the basic science, rationale, and clinical applications, existing and potential. Int J Nanomed 2006;1:297–315.Search in Google Scholar
17. Thomsen HS, Morcos SK, Almen T, Bellin MF, Bertolotto M, Bongartz G, et al. Nephrogenic systemic fibrosis and gadolinium-based contrast media: updated ESUR Contrast Medium Safety Committee guidelines. Euro radiol 2013;23:307–18.10.1007/s00330-012-2597-9Search in Google Scholar PubMed
18. Petersen AL, Hansen AE, Gabizon A, Andresen TL. Liposome imaging agents in personalized medicine. Adv Drug Deliver Rev 2012;64:1417–35.10.1016/j.addr.2012.09.003Search in Google Scholar PubMed
19. Geraldes CF, Laurent S. Classification and basic properties of contrast agents for magnetic resonance imaging. Contrast Media Molec Imaging 2009;4:1–23.10.1002/cmmi.265Search in Google Scholar PubMed
20. Erdogan S. Liposomal nanocarriers for tumor imaging. J Biomed Nanotechnol 2009;5:141–50.10.1166/jbn.2009.1016Search in Google Scholar PubMed
21. Cormode DP, Skajaa T, Fayad ZA, Mulder WJ. Nanotechnology in medical imaging: probe design and applications. Arterioscler Thromb Vasc Biol 2009;29:992–1000.10.1161/ATVBAHA.108.165506Search in Google Scholar PubMed PubMed Central
22. Kelly C, Jefferies C, Cryan SA. Targeted liposomal drug delivery to monocytes and macrophages. J Drug Deliver 2011;2011:727241.10.1155/2011/727241Search in Google Scholar PubMed PubMed Central
23. Wickline SA, Neubauer AM, Winter PM, Caruthers SD, Lanza GM. Molecular imaging and therapy of atherosclerosis with targeted nanoparticles. J Magn Reson Im 2007;25:667–80.10.1002/jmri.20866Search in Google Scholar PubMed
24. Lobatto ME, Calcagno C, Metselaar JM, Storm G, Stroes ES, Fayad ZA, et al. Imaging the efficacy of anti-inflammatory liposomes in a rabbit model of atherosclerosis by non-invasive imaging. Method enzymol 2012;508:211–28.10.1016/B978-0-12-391860-4.00011-2Search in Google Scholar PubMed PubMed Central
25. Lobatto ME, Fuster V, Fayad ZA, Mulder WJ. Perspectives and opportunities for nanomedicine in the management of atherosclerosis. Nat Rev Drug discovery 2011;10:835–52.10.1038/nrd3578Search in Google Scholar
26. Levchenko TS, Hartner WC, Torchilin VP. Liposomes for cardiovascular targeting. Ther deliver 2012;3:501–14.10.4155/tde.12.18Search in Google Scholar
27. Mackness M, Durrington P, Mackness B. Paraoxonase 1 activity, concentration and genotype in cardiovascular disease. Current opin lipidol 2004;15:399–404.10.1097/01.mol.0000137227.54278.29Search in Google Scholar
28. Acton S, Rigotti A, Landschulz KT, Xu S, Hobbs HH, Krieger M. Identification of scavenger receptor SR-BI as a high density lipoprotein receptor. Science 1996;271:518–20.10.1126/science.271.5248.518Search in Google Scholar
29. Grundy SM, Balady GJ, Criqui MH, Fletcher G, Greenland P, Hiratzka LF, et al. Primary prevention of coronary heart disease: guidance from Framingham: a statement for healthcare professionals from the AHA Task Force on Risk Reduction. American Heart Association. Circulation 1998;97:1876–87.10.1161/01.CIR.97.18.1876Search in Google Scholar
30. Rosenson RS, Brewer HB, Jr., Chapman MJ, Fazio S, Hussain MM, Kontush A, et al. HDL measures, particle heterogeneity, proposed nomenclature, and relation to atherosclerotic cardiovascular events. Clin Chem 2011;57:392–410.10.1373/clinchem.2010.155333Search in Google Scholar
31. Ng KK, Lovell JF, Zheng G. Lipoprotein-inspired nanoparticles for cancer theranostics. Accounts chem res 2011;44:1105–13.10.1021/ar200017eSearch in Google Scholar
32. Liu X, Suo R, Xiong SL, Zhang QH, Yi GH. HDL drug carriers for targeted therapy. Clin Chim Acta 2013;415:94–100.10.1016/j.cca.2012.10.008Search in Google Scholar
33. Sirtori CR, Calabresi L, Franceschini G. Recombinant apolipoproteins for the treatment of vascular diseases. Atherosclerosis 1999;142:29–40.10.1016/S0021-9150(98)00247-0Search in Google Scholar
34. Murphy AJ, Remaley AT, Sviridov D. HDL therapy: two kinds of right? Curr Pharm Design 2010;16:4134–47.10.2174/138161210794519228Search in Google Scholar PubMed
35. Ibanez B, Giannarelli C, Cimmino G, Santos-Gallego CG, Alique M, Pinero A, et al. Recombinant HDL(Milano) exerts greater anti-inflammatory and plaque stabilizing properties than HDL(wild-type). Atherosclerosis 2012;220:72–7.10.1016/j.atherosclerosis.2011.10.006Search in Google Scholar PubMed
36. Singh AT, Evens AM, Anderson RJ, Beckstead JA, Sankar N, Sassano A, et al. All trans retinoic acid nanodisks enhance retinoic acid receptor mediated apoptosis and cell cycle arrest in mantle cell lymphoma. Br J Haematol 2010;150:158–69.10.1111/j.1365-2141.2010.08209.xSearch in Google Scholar PubMed PubMed Central
37. Oda MN, Hargreaves PL, Beckstead JA, Redmond KA, van Antwerpen R, Ryan RO. Reconstituted high density lipoprotein enriched with the polyene antibiotic amphotericin B. J Lipid Res 2006;47:260–7.10.1194/jlr.D500033-JLR200Search in Google Scholar PubMed
38. Wolfrum C, Shi S, Jayaprakash KN, Jayaraman M, Wang G, Pandey RK, et al. Mechanisms and optimization of in vivo delivery of lipophilic siRNAs. Nature Biotechnol 2007;25:1149–57.10.1038/nbt1339Search in Google Scholar PubMed
39. Naresh NK, Xu Y, Klibanov AL, Vandsburger MH, Meyer CH, Leor J, et al. Monocyte and/or macrophage infiltration of heart after myocardial infarction: MR imaging by using T1-shortening liposomes. Radiology 2012;264:428–35.10.1148/radiol.12111863Search in Google Scholar PubMed PubMed Central
40. Duivenvoorden R, Tang J, Cormode DP, Mieszawska AJ, Izquierdo-Garcia D, Ozcan C, et al. A statin-loaded reconstituted high-density lipoprotein nanoparticle inhibits atherosclerotic plaque inflammation. Nature Commun 2014;5:3065.10.1038/ncomms4065Search in Google Scholar PubMed PubMed Central
41. Cormode DP, Skajaa T, van Schooneveld MM, Koole R, Jarzyna P, Lobatto ME, et al. Nanocrystal core high-density lipoproteins: a multimodality contrast agent platform. Nano Lett 2008;8:3715–23.10.1021/nl801958bSearch in Google Scholar PubMed PubMed Central
42. Skajaa T, Cormode DP, Jarzyna PA, Delshad A, Blachford C, Barazza A, et al. The biological properties of iron oxide core high-density lipoprotein in experimental atherosclerosis. Biomaterials 2011;32:206–13.10.1016/j.biomaterials.2010.08.078Search in Google Scholar PubMed PubMed Central
43. Barazza A, Blachford C, Even-Or O, Joaquin VA, Briley-Saebo KC, Chen W, et al. The complex fate in plasma of gadolinium incorporated into high-density lipoproteins used for magnetic imaging of atherosclerotic plaques. Bioconjugate Chem 2013;24:1039–48.10.1021/bc400105jSearch in Google Scholar PubMed PubMed Central
44. Packard RR, Libby P. Inflammation in atherosclerosis: from vascular biology to biomarker discovery and risk prediction. Clin Chem 2008;54:24–38.10.1373/clinchem.2007.097360Search in Google Scholar PubMed
45. Smedbakken LM, Halvorsen B, Daissormont I, Ranheim T, Michelsen AE, Skjelland M, et al. Increased levels of the homeostatic chemokine CXCL13 in human atherosclerosis-Potential role in plaque stabilization. Atherosclerosis 2012;224:266–73.10.1016/j.atherosclerosis.2012.06.071Search in Google Scholar PubMed
46. Signorelli SS, Anzaldi M, Fiore V, Simili M, Puccia G, Libra M, et al. Patients with unrecognized peripheral arterial disease (PAD) assessed by ankle-brachial index (ABI) present a defined profile of proinflammatory markers compared to healthy subjects. Cytokine 2012;59:294–8.10.1016/j.cyto.2012.04.038Search in Google Scholar PubMed
47. Di Taranto MD, Morgante A, Bracale UM, D’Armiento FP, Porcellini M, Bracale G, et al. Altered expression of inflammation-related genes in human carotid atherosclerotic plaques. Atherosclerosis 2012;220:93–101.10.1016/j.atherosclerosis.2011.10.022Search in Google Scholar PubMed
48. Schuett H, Oestreich R, Waetzig GH, Annema W, Luchtefeld M, Hillmer A, et al. Transsignaling of interleukin-6 crucially contributes to atherosclerosis in mice. Arterioscler Thromb Vasc Biol 2012;32:281–90.10.1161/ATVBAHA.111.229435Search in Google Scholar PubMed
49. Butcher M, Galkina E. Current views on the functions of interleukin-17A-producing cells in atherosclerosis. Thromb Haemostasis 2011;106:787–95.10.1160/TH11-05-0342Search in Google Scholar PubMed PubMed Central
50. Gonzalez-Gay MA, Gonzalez-Juanatey C, Llorca J, Castaneda S. The influence of inflammation in the development of subclinical atherosclerosis in psoriatic arthritis: comment on ‘Cardiovascular comorbidities in patients with psoriatic arthritis: a systematic review’ by Jamnistki et al. Ann Rheum Dis 2014;73:e27.10.1136/annrheumdis-2013-205154Search in Google Scholar PubMed
51. Papagoras C, Voulgari PV, Drosos AA. Atherosclerosis and cardiovascular disease in the spondyloarthritides, particularly ankylosing spondylitis and psoriatic arthritis. Clin Exp Rheumatol 2013;31:612–20.Search in Google Scholar
52. Ait-Oufella H, Taleb S, Mallat Z, Tedgui A. Recent advances on the role of cytokines in atherosclerosis. Arterioscler Thromb Vasc Biol 2011;31:969–79.10.1161/ATVBAHA.110.207415Search in Google Scholar PubMed
53. Chistiakov DA, Sobenin IA, Orekhov AN. Regulatory T cells in atherosclerosis and strategies to induce the endogenous atheroprotective immune response. Immunol Lett 2013; 151:10–22.10.1016/j.imlet.2013.01.014Search in Google Scholar PubMed
54. Zhu J, Chen T, Yang L, Li Z, Wong MM, Zheng X, et al. Regulation of microRNA-155 in atherosclerotic inflammatory responses by targeting MAP3K10. PloS One 2012;7:e46551.10.1371/journal.pone.0046551Search in Google Scholar PubMed PubMed Central
55. Foks AC, Bot I, Frodermann V, de Jager SC, Ter Borg M, van Santbrink PJ, et al. Interference of the CD30-CD30L pathway reduces atherosclerosis development. Arterioscler Thromb Vasc Biol 2012;32:2862–8.10.1161/ATVBAHA.112.300509Search in Google Scholar PubMed
56. Subramanian M, Thorp E, Hansson GK, Tabas I. Treg-mediated suppression of atherosclerosis requires MYD88 signaling in DCs. The J Clin Invest 2013;123:179–88.10.1172/JCI64617Search in Google Scholar PubMed PubMed Central
57. Pinderski Oslund LJ, Hedrick CC, Olvera T, Hagenbaugh A, Territo M, Berliner JA, et al. Interleukin-10 blocks atherosclerotic events in vitro and in vivo. Arterioscler Thromb Vasc Biol 1999;19:2847–53.10.1161/01.ATV.19.12.2847Search in Google Scholar PubMed
58. Pinderski LJ, Fischbein MP, Subbanagounder G, Fishbein MC, Kubo N, Cheroutre H, et al. Overexpression of interleukin-10 by activated T lymphocytes inhibits atherosclerosis in LDL receptor-deficient Mice by altering lymphocyte and macrophage phenotypes. Circ Res 2002;90:1064–71.10.1161/01.RES.0000018941.10726.FASearch in Google Scholar PubMed
59. Xie G, Myint PK, Zaman MJ, Li Y, Zhao L, Shi P, et al. Relationship of serum interleukin-10 and its genetic variations with ischemic stroke in a Chinese general population. PloS One 2013;8:e74126.10.1371/journal.pone.0074126Search in Google Scholar PubMed PubMed Central
60. Mallat Z, Heymes C, Ohan J, Faggin E, Leseche G, Tedgui A. Expression of interleukin-10 in advanced human atherosclerotic plaques: relation to inducible nitric oxide synthase expression and cell death. Arterioscler Thromb Vasc Biol 1999;19: 611–6.10.1161/01.ATV.19.3.611Search in Google Scholar
61. Mallat Z, Besnard S, Duriez M, Deleuze V, Emmanuel F, Bureau MF, et al. Protective role of interleukin-10 in atherosclerosis. Circ Res 1999;85:e17–24.10.1161/01.RES.85.8.e17Search in Google Scholar
62. Nishihira K, Imamura T, Yamashita A, Hatakeyama K, Shibata Y, Nagatomo Y, et al. Increased expression of interleukin-10 in unstable plaque obtained by directional coronary atherectomy. Eur Heart J 2006;27:1685–9.10.1093/eurheartj/ehl058Search in Google Scholar PubMed
63. Terkeltaub RA. IL-10: An “immunologic scalpel” for atherosclerosis? Arterioscler Thromb Vasc Biol 1999;19:2823–5.10.1161/01.ATV.19.12.2823Search in Google Scholar PubMed
64. McCarthy C, Duffy MM, Mooney D, James WG, Griffin MD, Fitzgerald DJ, et al. IL-10 mediates the immunoregulatory response in conjugated linoleic acid-induced regression of atherosclerosis. FASEB journal: official publication of the Fed Am Soc Exp Biol 2013;27:499–510.10.1096/fj.12-215442Search in Google Scholar PubMed
65. de Waal Malefyt R, Abrams J, Bennett B, Figdor CG, de Vries JE. Interleukin 10(IL-10) inhibits cytokine synthesis by human monocytes: an autoregulatory role of IL-10 produced by monocytes. J Exp Med 1991;174:1209–20.10.1084/jem.174.5.1209Search in Google Scholar PubMed PubMed Central
66. Peguet-Navarro J, Moulon C, Caux C, Dalbiez-Gauthier C, Banchereau J, Schmitt D. Interleukin-10 inhibits the primary allogeneic T cell response to human epidermal Langerhans cells. Eur J Immunol 1994;24:884–91.10.1002/eji.1830240416Search in Google Scholar
67. Fiorentino DF, Zlotnik A, Mosmann TR, Howard M, O’Garra A. IL-10 inhibits cytokine production by activated macrophages. J Immunol 1991;147:3815–22.10.4049/jimmunol.147.11.3815Search in Google Scholar
68. O’Garra A, Vieira P. T(H)1 cells control themselves by producing interleukin-10. Nat Rev Immunol 2007;7:425–8.10.1038/nri2097Search in Google Scholar
69. Le Gros G, Erard F. Non-cytotoxic, IL-4, IL-5, IL-10 producing CD8+ T cells: their activation and effector functions. Curr Opin Immunol 1994;6:453–7.10.1016/0952-7915(94)90127-9Search in Google Scholar
70. Fillatreau S, Gray D, Anderton SM. Not always the bad guys: B cells as regulators of autoimmune pathology. Nat Rev Immunol 2008;8:391–7.10.1038/nri2315Search in Google Scholar PubMed
71. Ryan JJ, Kashyap M, Bailey D, Kennedy S, Speiran K, Brenzovich J, et al. Mast cell homeostasis: a fundamental aspect of allergic disease. Crit Rev Immunol 2007;27:15–32.10.1615/CritRevImmunol.v27.i1.20Search in Google Scholar
72. Moore KW, de Waal Malefyt R, Coffman RL, O’Garra A. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol 2001;19:683–765.10.1146/annurev.immunol.19.1.683Search in Google Scholar PubMed
73. Williams LM, Ricchetti G, Sarma U, Smallie T, Foxwell BM. Interleukin-10 suppression of myeloid cell activation – a continuing puzzle. Immunology 2004;113:281–92.10.1111/j.1365-2567.2004.01988.xSearch in Google Scholar PubMed PubMed Central
74. Donnelly RP, Dickensheets H, Finbloom DS. The interleukin-10 signal transduction pathway and regulation of gene expression in mononuclear phagocytes. J Interferon Cytokine Res 1999;19:563–73.10.1089/107999099313695Search in Google Scholar PubMed
75. Kershaw EE, Flier JS. Adipose tissue as an endocrine organ. J Clin Endocrinol Metab 2004;89:2548–56.10.1210/jc.2004-0395Search in Google Scholar PubMed
76. Romacho T, Elsen M, Rohrborn D, Eckel J. Adipose Tissue and its Role in Organ Crosstalk. Acta physiologica 2014;210:733–53.10.1111/apha.12246Search in Google Scholar PubMed
77. Alexopoulos N, Katritsis D, Raggi P. Visceral adipose tissue as a source of inflammation and promoter of atherosclerosis. Atherosclerosis 2014;233:104–12.10.1016/j.atherosclerosis.2013.12.023Search in Google Scholar PubMed
78. Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun 1996;221:286–9.10.1006/bbrc.1996.0587Search in Google Scholar PubMed
79. Scherer PE, Williams S, Fogliano M, Baldini G, Lodish HF. A novel serum protein similar to C1q, produced exclusively in adipocytes. J Biol Chem 1995;270:26746–9.10.1074/jbc.270.45.26746Search in Google Scholar PubMed
80. Chandran M, Phillips SA, Ciaraldi T, Henry RR. Adiponectin: more than just another fat cell hormone? Diabetes Care 2003;26:2442–50.10.2337/diacare.26.8.2442Search in Google Scholar PubMed
81. Arita Y, Kihara S, Ouchi N, Takahashi M, Maeda K, Miyagawa J, et al. Paradoxical decrease of an adipose-specific protein, adiponectin, in obesity. Biochem Biophys Res Commun 1999;257:79–83.10.1006/bbrc.1999.0255Search in Google Scholar PubMed
82. Mangge H, Almer G, Haj-Yahya S, Grandits N, Gasser R, Pilz S, et al. Nuchal thickness of subcutaneous adipose tissue is tightly associated with an increased LMW/total adiponectin ratio in obese juveniles. Atherosclerosis 2009;203:277–83.10.1016/j.atherosclerosis.2008.06.013Search in Google Scholar PubMed
83. Mangge H, Almer G, Haj-Yahya S, Pilz S, Gasser R, Moller R, et al. Preatherosclerosis and adiponectin subfractions in obese adolescents. Obesity 2008;16:2578–84.10.1038/oby.2008.439Search in Google Scholar PubMed
84. Bobbert T, Rochlitz H, Wegewitz U, Akpulat S, Mai K, Weickert MO, et al. Changes of adiponectin oligomer composition by moderate weight reduction. Diabetes 2005;54:2712–9.10.2337/diabetes.54.9.2712Search in Google Scholar PubMed
85. van Stijn CM, Kim J, Barish GD, Tietge UJ, Tangirala RK. Adiponectin expression protects against angiotensin ii-mediated inflammation and accelerated atherosclerosis. PloS One 2014;9:e86404.10.1371/journal.pone.0086404Search in Google Scholar PubMed PubMed Central
86. Luo N, Liu J, Chung BH, Yang Q, Klein RL, Garvey WT, et al. Macrophage adiponectin expression improves insulin sensitivity and protects against inflammation and atherosclerosis. Diabetes 2010;59:791–9.10.2337/db09-1338Search in Google Scholar PubMed PubMed Central
87. Pilz S, Horejsi R, Moller R, Almer G, Scharnagl H, Stojakovic T, et al. Early atherosclerosis in obese juveniles is associated with low serum levels of adiponectin. J Clin Endocrinol Metabol 2005;90:4792–6.10.1210/jc.2005-0167Search in Google Scholar PubMed
88. Pilz S, Maerz W, Weihrauch G, Sargsyan K, Almer G, Nauck M, et al. Adiponectin serum concentrations in men with coronary artery disease: the LUdwigshafen RIsk and Cardiovascular Health (LURIC) study. Clin Chim Ata; Int J Clin Chem 2006;364:251–5.10.1016/j.cccn.2005.07.011Search in Google Scholar PubMed
89. Li CJ, Sun HW, Zhu FL, Chen L, Rong YY, Zhang Y, et al. Local adiponectin treatment reduces atherosclerotic plaque size in rabbits. J Endocrinol 2007;193:137–45.10.1677/JOE-06-0173Search in Google Scholar PubMed
90. Okamoto Y, Ishii S, Croce K, Katsumata H, Fukushima M, Kihara S, et al. Adiponectin inhibits macrophage tissue factor, a key trigger of thrombosis in disrupted atherosclerotic plaques. Atherosclerosis 2013;226:373–7.10.1016/j.atherosclerosis.2012.12.012Search in Google Scholar PubMed PubMed Central
91. Ouchi N, Ohishi M, Kihara S, Funahashi T, Nakamura T, Nagaretani H, et al. Association of hypoadiponectinemia with impaired vasoreactivity. Hypertension 2003;42:231–4.10.1161/01.HYP.0000083488.67550.B8Search in Google Scholar PubMed
92. Shimabukuro M, Higa N, Asahi T, Oshiro Y, Takasu N, Tagawa T, et al. Hypoadiponectinemia is closely linked to endothelial dysfunction in man. J Clin Endocrinol Metab 2003;88:3236–40.10.1210/jc.2002-021883Search in Google Scholar PubMed
93. Lavoie V, Kernaleguen AE, Charron G, Farhat N, Cossette M, Mamarbachi AM, et al. Functional effects of adiponectin on endothelial progenitor cells. Obesity (Silver Spring) 2011;19:722–8.10.1038/oby.2010.187Search in Google Scholar PubMed
94. Hitsumoto T, Takahashi M, Iizuka T, Shirai K. Relationship between metabolic syndrome and early stage coronary atherosclerosis. J Atheroscler Thromb 2007;14:294–302.10.5551/jat.E506Search in Google Scholar PubMed
95. Waki H, Yamauchi T, Kamon J, Kita S, Ito Y, Hada Y, et al. Generation of globular fragment of adiponectin by leukocyte elastase secreted by monocytic cell line THP-1. Endocrinology 2005;146:790–6.10.1210/en.2004-1096Search in Google Scholar PubMed
96. Fruebis J, Tsao TS, Javorschi S, Ebbets-Reed D, Erickson MR, Yen FT, et al. Proteolytic cleavage product of 30-kDa adipocyte complement-related protein increases fatty acid oxidation in muscle and causes weight loss in mice. Proc Natl Acad Sci USA 2001;98:2005–10.10.1073/pnas.98.4.2005Search in Google Scholar PubMed PubMed Central
97. Bruce CR, Mertz VA, Heigenhauser GJ, Dyck DJ. The stimulatory effect of globular adiponectin on insulin-stimulated glucose uptake and fatty acid oxidation is impaired in skeletal muscle from obese subjects. Diabetes 2005;54:3154–60.10.2337/diabetes.54.11.3154Search in Google Scholar PubMed
98. Palanivel R, Fang X, Park M, Eguchi M, Pallan S, De Girolamo S, et al. Globular and full-length forms of adiponectin mediate specific changes in glucose and fatty acid uptake and metabolism in cardiomyocytes. Cardiovasc Res 2007;75:148–57.10.1016/j.cardiores.2007.04.011Search in Google Scholar PubMed
99. Sun Y, Xun K, Wang C, Zhao H, Bi H, Chen X, et al. Adiponectin, an unlocking adipocytokine. Cardiovasc Ther 2009;27:59–75.10.1111/j.1755-5922.2008.00069.xSearch in Google Scholar PubMed
100. Sun Y, Chen X. Effect of adiponectin on apoptosis: proapoptosis or antiapoptosis? Biofactors 2010;36:179–86.10.1002/biof.83Search in Google Scholar PubMed
101. Fang X, Sweeney G. Mechanisms regulating energy metabolism by adiponectin in obesity and diabetes. Biochem Soc Trans 2006;34:798–801.10.1042/BST0340798Search in Google Scholar PubMed
102. Belo VA, Souza-Costa DC, Lacchini R, Sertorio JT, Lanna CM, Carmo VP, et al. Adiponectin associates positively with nitrite levels in children and adolescents. Int J Obes (Lond) 2013;37:740–3.10.1038/ijo.2012.104Search in Google Scholar PubMed
103. Chen H, Montagnani M, Funahashi T, Shimomura I, Quon MJ. Adiponectin stimulates production of nitric oxide in vascular endothelial cells. J Biol Chem 2003;278:45021–6.10.1074/jbc.M307878200Search in Google Scholar PubMed
104. Bi CW, Xu L, Tian XY, Liu J, Zheng KY, Lau CW, et al. Fo Shou San, an ancient Chinese herbal decoction, protects endothelial function through increasing endothelial nitric oxide synthase activity. PloS One 2012;7:e51670.10.1371/journal.pone.0051670Search in Google Scholar PubMed PubMed Central
105. Shimada K, Miyazaki T, Daida H. Adiponectin and atherosclerotic disease. Clin Chim Acta 2004;344:1–12.10.1016/j.cccn.2004.02.020Search in Google Scholar PubMed
106. Adya R, Tan BK, Chen J, Randeva HS. Protective actions of globular and full-length adiponectin on human endothelial cells: novel insights into adiponectin-induced angiogenesis. J Vasc Res 2012;49:534–43.10.1159/000338279Search in Google Scholar PubMed
107. Araki S, Dobashi K, Kubo K, Asayama K, Shirahata A. High molecular weight, rather than total, adiponectin levels better reflect metabolic abnormalities associated with childhood obesity. J Clin Endocrinol Metab 2006;91:5113–6.10.1210/jc.2006-1051Search in Google Scholar PubMed
108. Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003;423:762–9.10.1038/nature01705Search in Google Scholar PubMed
109. Hug C, Wang J, Ahmad NS, Bogan JS, Tsao TS, Lodish HF. T-cadherin is a receptor for hexameric and high-molecular-weight forms of Acrp30/adiponectin. Proc Natl Acad Sci USA 2004;101:10308–13.10.1073/pnas.0403382101Search in Google Scholar PubMed PubMed Central
110. Takemura Y, Ouchi N, Shibata R, Aprahamian T, Kirber MT, Summer RS, et al. Adiponectin modulates inflammatory reactions via calreticulin receptor-dependent clearance of early apoptotic bodies. J Clin Invest 2007;117:375–86.10.1172/JCI29709Search in Google Scholar PubMed PubMed Central
111. Okamoto Y, Arita Y, Nishida M, Muraguchi M, Ouchi N, Takahashi M, et al. An adipocyte-derived plasma protein, adiponectin, adheres to injured vascular walls. Horm Metab Res 2000;32:47–50.10.1055/s-2007-978586Search in Google Scholar PubMed
112. Ouchi N, Kihara S, Arita Y, Nishida M, Matsuyama A, Okamoto Y, et al. Adipocyte-derived plasma protein, adiponectin, suppresses lipid accumulation and class A scavenger receptor expression in human monocyte-derived macrophages. Circulation 2001;103:1057–63.10.1161/01.CIR.103.8.1057Search in Google Scholar PubMed
113. Pilz S, Mangge H, Wellnitz B, Seelhorst U, Winkelmann BR, Tiran B, et al. Adiponectin and mortality in patients undergoing coronary angiography. J Clin Endocrinol Metabol 2006;91:4277–86.10.1210/jc.2006-0836Search in Google Scholar PubMed
114. Junghans M, Kreuter J, Zimmer A. Antisense delivery using protamine-oligonucleotide particles. Nucleic Acids Res 2000;28:E45.10.1093/nar/28.10.e45Search in Google Scholar PubMed PubMed Central
115. Lochmann D, Weyermann J, Georgens C, Prassl R, Zimmer A. Albumin-protamine-oligonucleotide nanoparticles as a new antisense delivery system. Part 1: physicochemical characterization. Eur J Pharm Biopharm 2005;59:419–29.10.1016/j.ejpb.2004.04.001Search in Google Scholar PubMed
116. Weyermann J, Lochmann D, Georgens C, Zimmer A. Albumin-protamine-oligonucleotide-nanoparticles as a new antisense delivery system. Part 2: cellular uptake and effect. Eur J Pharm Biopharm 2005;59:431–8.10.1016/j.ejpb.2004.07.014Search in Google Scholar PubMed
117. Wernig K, Griesbacher M, Andreae F, Hajos F, Wagner J, Mosgoeller W, et al. Depot formulation of vasoactive intestinal peptide by protamine-based biodegradable nanoparticles. J Control Release 2008;130:192–8.10.1016/j.jconrel.2008.06.005Search in Google Scholar PubMed
118. Rader DJ, Parmacek MS. Secreted miRNAs suppress atherogenesis. Nature Cell Biol 2012;14:233–5.10.1038/ncb2452Search in Google Scholar PubMed PubMed Central
119. Mayr M, Zampetaki A, Willeit P, Willeit J, Kiechl S. MicroRNAs within the continuum of postgenomics biomarker discovery. Arteriosclerosis, Thromb Vasc Biol 2013;33:206–14.10.1161/ATVBAHA.112.300141Search in Google Scholar PubMed
120. van Dijk RA, Kolodgie F, Ravandi A, Leibundgut G, Hu PP, Prasad A, et al. Differential expression of oxidation-specific epitopes and apolipoprotein(a) in progressing and ruptured human coronary and carotid atherosclerotic lesions. J Lipid Res 2012;53:2773–90.10.1194/jlr.P030890Search in Google Scholar PubMed PubMed Central
©2014 by Walter de Gruyter Berlin/Boston
Articles in the same Issue
- Frontmatter
- Guest Editorial and News
- Cardiovascular nanomedicine
- News from the European Foundation for Nanomedicine (CLINAM): The 2014 CLINAM Summit
- What’s up in nanomedicine for cardiovascular diseases?
- Review Articles
- Cardiovascular therapy through nanotechnology – how far are we still from bedside?
- The simultaneous systematic analysis approach for personalized management of cardiovascular diseases
- Anti-inflammatory mediators for molecular imaging of atherosclerosis
- Perspectives
- Atherosclerotic disease and management challenges with nanomedicine: EU FP7 NMP funded “NanoAthero” and “CosmoPHOS-nano” large-scale projects
Articles in the same Issue
- Frontmatter
- Guest Editorial and News
- Cardiovascular nanomedicine
- News from the European Foundation for Nanomedicine (CLINAM): The 2014 CLINAM Summit
- What’s up in nanomedicine for cardiovascular diseases?
- Review Articles
- Cardiovascular therapy through nanotechnology – how far are we still from bedside?
- The simultaneous systematic analysis approach for personalized management of cardiovascular diseases
- Anti-inflammatory mediators for molecular imaging of atherosclerosis
- Perspectives
- Atherosclerotic disease and management challenges with nanomedicine: EU FP7 NMP funded “NanoAthero” and “CosmoPHOS-nano” large-scale projects