Beating the bugs: roles of microbial biofilms in corrosion
-
Kwan Li is a Master’s candidate in Materials Science and Engineering at MIT, working with Van Vliet on microbiologically influenced corrosion via a focus on electrochemistry. Li earned his BS (Duke University, 2011) in Mechanical Engineering and Biomedical Engineering, with a minor in chemistry.
Matthew Whitfield is a postdoctoral research associate in Materials Science and Engineering at MIT, working with Van Vliet. He has developed experimental assays and computational models of bacterial adhesion, including biofilm analysis and corrosion onset in ferrous alloys. Whitfield earned his BE (Vanderbilt University, 2006) in Biomedical Engineering and PhD (University of Washington, 2011) in Bioengineering.
Krystyn J. Van Vliet is the SMART Scholar Associate Professor of Materials Science and Engineering and Biological Engineering at MIT. She and her group study coupling between chemistry and mechanics at material interfaces, including the cell-material interface. As one example, her group studies chemomechanics of abiotic corrosion and biotic corrosion of ferrous alloys in sour environments, via both experiments and simulations. Van Vliet earned her ScB (Brown University, 1998) and PhD (MIT, 2002) in Materials Science and Engineering.


Abstract
Microbiologically influenced corrosion (MIC) is a complex type of environmentally assisted corrosion. Although poorly understood and challenging to ameliorate, it is increasingly appreciated that MIC accelerates failure of metal alloys, including steel pipeline. Historically, this type of material degradation process has been treated from either an electrochemical materials perspective or a microbiological perspective. Here, we review the current understanding of MIC mechanisms for steel – particularly those in sour environments relevant to fossil fuel recovery and processing – and outline the role of the bacterial biofilm in both corrosion processes and mitigation responses.
1 Introduction
Microbiologically influenced corrosion (MIC) can accelerate mechanical failure of metals in a wide range of environments ranging from oil and water pipelines and machinery to biomedical devices. Although this mode of environmentally assisted corrosion has no single definition, MIC invariably includes corrosion in the presence of microbial species including bacteria and the associated biofilms produced by those bacteria. MIC has been noted to occur for a variety of metals and alloys such as iron and steel, copper, and titanium (Rao, 2012), as well as on nonmetallic materials such as concrete (George, 2012). For instance, MIC and associated biofouling are estimated to account for 20% of direct fuel pipeline integrity and reliability costs exceeding $2 billion USD per year (Flemming, 1996; Koch, Brongers, Thompson, Virmani, & Payer, 2002). Recently, this corrosion mechanism has been implicated in several rapid, high-profile failures of pipeline including a light grade crude oil pipeline failure (Al-Jaroudi, Ul-Hamid, & Al-Gahtani, 2011), a seawater pipeline failure within a few years of installation (Abraham, Kain, & Dey, 2009), leaks in a stainless steel cooling water system (Yu, Dillon, & Henry, 2010), and leaks in the Trans-Alaskan Pipeline (Egan, 2011).
Despite decades of research, the exact mechanisms by which and environments in which MIC enhances corrosion are poorly understood. Indeed, even within the same oilfield or pipeline, some locations may exhibit MIC whereas others do not (Maruthamuthu et al., 2011; Voordouw et al., 2009; Yu et al., 2010). Conventionally, this phenomenon has been studied either from a biological perspective – considering the complex nature of bacteria consortia and biofilm formation – or from a materials corrosion perspective – considering the evidence and mitigation of metal pitting under controlled conditions. Therefore, there is no consensus on how to mitigate corrosive losses due to MIC. Here, we review the most current understanding of MIC mechanisms on steel (i.e., iron-carbon or ferrous alloys), MIC mitigation strategies, and the role of the bacterial biofilm in both corrosion and mitigation response.
2 Characteristics of MIC
MIC can occur in both aerobic and anaerobic conditions, and is defined by no single characteristic trait other than corrosion in the presence of microbial species (bacteria) and their associated biofilms. Thus, MIC is considered biotic corrosion, and the terms “microbes” and “bacteria” are used interchangeably. As bacteria are ubiquitous in almost all environments (air, water, and organic fluids), the mere presence of bacteria found at sites of corrosion does not necessarily mean that the bacteria were a significant contributing factor in corrosion initiation or acceleration. MIC can occur in conjunction with other types of abiotic corrosion, further obfuscating attempts to identify the root cause of corrosion failure. Often, MIC is associated with surface pitting, which leads to more rapid corrosive failure than uniform corrosion (Bryant, Jansen, Boivin, Laishley, & Costerton, 1991; Chamritski, Burns, Laycock, & Webster, 2004; Jan-Roblero, Romero, Amaya, & Le Borgne, 2004; Miranda et al., 2006; Yuan & Pehkonen, 2009) (Figure 1). Thus, even nonferrous metals that exhibit a naturally passivating layer at the surface (e.g., copper oxides and titanium oxides) exhibit MIC when metabolic byproducts of the microbes serve to chemically reduce and thus disrupt the passivating film under anaerobic conditions; this results in pitting corrosion at the metal-biofilm interface (Rao, 2012). As many surface characteristics including roughness, charge, and hydrophilicity can modulate bacterial adhesion, it is not yet clear whether bacteria initiate the pits or adhere preferentially to pre-existing pits. Although several studies have found evidence that microbes adhere preferentially to anodic metal regions such as welded joints, scratched edges, grain boundaries (Dexter & Eashwar, 1999; Sreekumari, Nandakumar, & Kikuchi, 2001), or to previously corroded locations (Little et al., 1999), a more recent study was not able to confirm these site-specific claims (Sherar et al., 2011). When many species of bacteria adhere and proliferate on a surface, they secrete a polysaccharide-rich matrix; a biofilm includes that extracellular polysaccharide matrix and the bacterial cells within. Attempts to spatially correlate localized corrosion with the overlying biofilm, for example, using electrochemical mapping techniques, have failed due to the conductive nature of the biofilm itself (Dong, Shi, Ruan, & Zhang, 2011). Further, although pitting is a known signature of MIC, others have found no unique characteristics that distinguish the morphology of MIC pits from abiotic pits (Thomas & Chung, 1999).
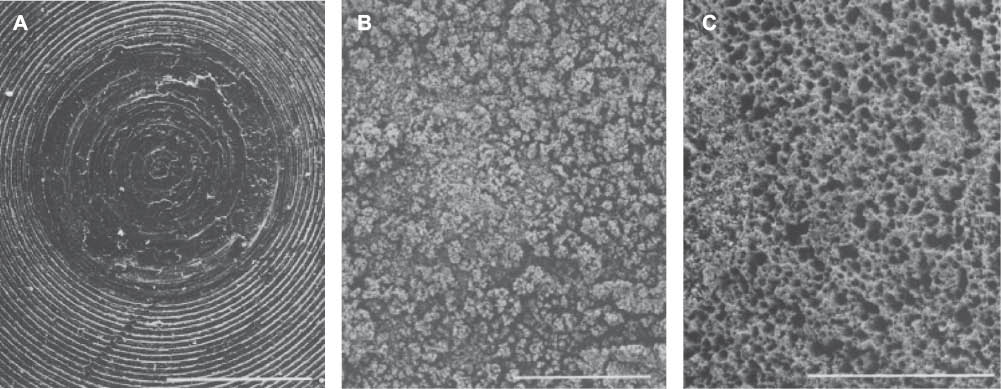
Scanning electron microscopy images which demonstrate the growth and pitting damage caused by sulfate-reducing bacteria growth on SAE 1020 steel studs. (A) Control stud showing concentric rings from lathe cutting; scale bar, 500 μm. (B) sulfate-reducing bacterial biofilm deposit after 64 days; scale bar, 500 μm. (C) Corroded stud after biofilm removal revealing severe pitting damage; scale bar, 500 μm (Bryant et al., 1991). Reprinted with permission from the American Society for Microbiology.
At sites of suspected MIC, a so-called consortium of several bacterial species is always found – rather than a single species – although the number and types of bacteria in that consortium varies widely from location to location. Because the culture conditions of the majority of bacterial species are not well established, determining the phylogenetic makeup is difficult. However, the application of molecular techniques now allow much more detailed analysis of extracts, quantifying the types and relative numbers of species (Amann, Ludwig, & Schleifer, 1995). Several studies have performed detailed phylogenetic characterization of fluids from gas and oil pipelines (Jan-Roblero et al., 2004; Maruthamuthu et al., 2011; Rajasekar, Anandkumar, Maruthamuthu, Ting, & Rahman, 2010) and water outlet pipes at oil production facilities (Larsen et al., 2010). The categories of bacteria typically directly linked to corrosion are sulfate-reducing bacteria (SRB), iron-oxidizing bacteria, sulfur-oxidizing bacteria, nitrate-reducing bacteria (NRB), and methanogens. These bacteria may reduce the metal directly, produce corrosive metabolic byproducts, and/or produce biofilms that indirectly alter the local environment to promote corrosion (Rao, 2012).
SRB are considered one of the main culprits of biotic corrosion in anaerobic conditions. SRB are a grouping of bacteria that includes at least 220 species that produce H2S, and use sulfates as the terminal electron acceptor (Barton & Fauque, 2009). Most SRB are considered obligate anaerobes, meaning that the cells cannot metabolize and/or replicate in the presence of oxygen, although many species can temporarily tolerate low levels of oxygen (Johnson, Zhulin, Gapuzan, & Taylor, 1997). Furthermore, anaerobic conditions capable of supporting SRB growth can be created in overall aerobic environments, due to the microniches created within the bacterial biofilm/corrosion product layer. Although SRB are the most studied and well understood of the anaerobic corrosion inducing bacteria, we note that MIC can occur in anaerobic conditions in the absence of SRB (Rajasekar et al., 2010). However, given the wide acknowledgment of SRB presence in industrial MIC contexts, and the substantial study of this bacteria grouping at the genetic and structural levels (Wall, Arkin, Balci, & Rapp-Giles, 2008), we will focus primarily on the role of this microbe type within the consortium and biofilms relevant to MIC of ferrous alloys.
3 Characteristics of MIC-associated biofilms
MIC is attributed not to planktonic bacteria but instead to adherent bacteria in the form of biofilms on surfaces. Many bacteria types adhere to and form polysaccharide-rich biofilms on material surfaces, with interactions governed strongly by the material surface properties, fluid properties, and bacteria adhesion mechanisms (Lichter et al., 2008; Lichter, Van Vliet, & Rubner, 2009). Biofilm formation follows a series of critical steps: an initial rapid and reversible bacterial attachment, a more stable, longer term attachment, bacterial replication and matrix secretion, biofilm maturation, and finally bacterial dispersal (Lichter et al., 2009; Monds & O’Toole, 2009). The main structural components of the biofilms are produced by the bacteria themselves through the secretion of extracellular polymeric substances (EPSs). EPSs are composed chiefly of polysaccharides, proteins, nucleic acids, and lipids (Vu, Chen, Crawford, & Ivanova, 2009). Biofilms can be anywhere from μm to mm in thickness. These structures can develop and mature over the course of hours, days, or months, depending on the bacterial species and environment (Figure 2). Repeated biofilm formation has the potential to occur due to pigging, an intentional mechanical delamination of corrosion products including iron sulfides within internal pipeline surfaces, or due to general biofilm rupture. In the context of fuel transport within pipelines, the sporadic flow rates and fluid composition can also disrupt and promote biofilm formation, and likewise the presence of the biofilm can affect flow trajectories. Essentially, through these biofilms the bacteria create a unique and sustaining microenvironment that can differ significantly – in terms of composition and distribution of solids, fluids, and gases – from the overall macroenvironment.

(A) Scanning electron microscopy images demonstrating the growth of a biofilm over the course of 60 days. All images are at 1000× except for 60 days which is 200×. Inner boxes are at 4000× (Fernandez, Diaz, Amils, & Sanz, 2008). Reprinted with permission from Springer. (B) Original image demonstrating a cross-section of the combined biofilm/corrosion product layer after 2 days of growth of Desulfovibrio vulgaris on music spring quality steel (ASTM A228).
The formation of a biofilm itself does not necessitate an increased corrosion rate and can actually lead to corrosion resistance in some circumstances. Certain bacteria within biofilms can remove corrosion-promoting agents, such as O2, (Dubiel, Hsu, Chien, Mansfeld, & Newman, 2002) as part of the bacteria metabolism, can secrete antimicrobial agents that inhibit growth of corrosion-causing bacteria, and can generate physical barriers that protect surfaces from corrosion (Avidan, Satanower, & Banin 2010; Zuo, 2007). The particular species makeup and environmental conditions greatly influence the corrosive potential of a biofilm. This variation has obfuscated the identification of a general, causal link between bacteria presence and corrosion. Such complexity prohibits the community from drawing broad conclusions on corrosion potential, mechanisms, and rates beyond the specific conditions in which observations were made. This difficulty has prompted the development of models to better predict when MIC will be a problem (Maxwell, 2006; Skovhus, Holmkvist, Andersen, Larsen, & Pedersen, 2012), but the applicability of such models is still currently limited to specific species and environmental conditions.
Although single species studies are naturally better controlled and simpler to interpret, recognition of the significance of the interplay among the different bacterial species within a biofilm has prompted multiple-species studies. For instance, Okabe, Itoh, Satoh, and Watanabe (1999) studied the spatial distribution of SRB in an aerobic wastewater biofilm and found that although SRB were located throughout the biofilm, sulfate reduction was focused in a narrow anoxic band. Andersson, Kuttuva Rajarao, Land, and Dalhammar (2008) systematically studied initial adherence and biofilm formation with different combinations of 13 bacterial species often found in wastewater systems, and found complex protagonistic and antagonistic interactions that depended on species composition. Interestingly, Lee, Buehler, and Newman (2006) found that a dual species biofilm with an SRB and iron-reducing bacteria resulted in reduced corrosion, as compared with a biofilm comprising only SRB. Videla, Borgne, Panter, and Singh Raman (2008) also found differences in corrosion with iron-reducing bacteria and SRB in mixed culture in different environments. Overall, however, these studies illustrate the complexity of the process rather than providing concrete conclusions about MIC mechanisms.
Environmental factors such as fluid shear flow found within pipelines add another layer of complexity to biofilm development and thus to MIC in industrial settings. It is known that shearing forces can greatly influence bacterial adhesion and biofilm development in dramatic ways, regardless of the context (Rochex, Godon, Bernet, & Escudie, 2008; Whitfield, Ghose, & Thomas, 2010). For example, in other bacterial species (e.g., aerobic species such as Escherichia coli), it has been shown that adhesion is strengthened in the presence of moderate flow rates. At lower rates, the so-called catch bond mechanism is not activated, and at higher rates the binding is again disrupted; only at intermediate rates are the adhesive structures at the bacterial surface configured to bind to the surface with highest affinity and longest lifetime (Thomas, Trintchina, Forero, Vogel, & Sokurenko, 2002; Whitfield et al., 2010). For SRB, it is unknown whether similar adhesive structures and mechanisms exist. However, moderate flow rates (0.1–0.5 m/s) have been shown to change SRB biofilm morphology, and to increase biofilm rigidity and yield shear stress (Dunsmore et al., 2002; Stoodley et al., 2001). At sufficiently high flow rates (3.5 m/s), SRB adhesion and biofilm growth are inhibited (Wen, Gu, & Nesic, 2007). Qualitatively, these results are similar to more widely studied aerobic biofilms, making it possible that mechanistic insights can be shared. In a study not directly related to corrosion research, Rochex et al. (2008) found that increased shear stresses lowered the species diversity within biofilms. A more robust understanding of how flow conditions affect MIC-associated biofilm structure, diversity, and corrosive potential would allow for improved risk assessment and pipeline flow rate and flow pattern selection.
4 Mitigation techniques
Several techniques have been investigated to mitigate the effects of MIC. These include using corrosion resistant metals, protective coatings, pigging, anodic and cathodic protection, biocides, introduction of competing bacteria, and nutrient addition or removal. Some techniques such as pigging (i.e., mechanical delamination of corrosion products) and biocide addition are widely used in field conditions that include sour (anaerobic, sulfur-containing) environments such as buried pipelines. Other approaches such as sol-gel coating barriers (Akid, Wang, Smith, Greenfield, & Earthman, 2008) or precoating with purified EPSs (Stadler, Wei, Furbeth, Grooters, & Kuklinski, 2010) are still in experimental stages. One of the more recently field tested techniques – and one that relies on a robust understanding of the biofilm – is to alter the corrosive ability of biofilms by manipulating the available nutrients via addition of nitrates or removal of sulfates.
In field conditions, nitrate addition (Voordouw et al., 2009) and sulfate removal (McElhiney, Burger, Maxwell, Davis, & Walsh, 2008) have been shown to affect SRB activity and be somewhat effective in reducing or altering corrosion. Nitrate addition can allow other bacteria such as NRB to outcompete the more corrosive SRB (Figure 3) (Hubert & Voordouw, 2007; Thorstenson et al., 2002). Additionally, there is evidence that nitrate can directly inhibit SRB metabolism and proliferation (Haveman, Greene, Stilwell, Voordouw, & Voordouw, 2004; He et al., 2010). However, nitrate addition is not a definitive solution to MIC. Although this approach typically reduces sulfide production, the effect is only temporary and SRB are not killed. Thus, sulfide production rapidly returns if active nitrate treatment is terminated (Mohanakrishnan et al., 2011). Another study showed that nitrate addition could reduce uniform surface corrosion but not pitting corrosion, the latter of which is often the cited cause of pipeline failures (Schwermer et al., 2008). Further, the same nutrient manipulation protocols work in some pipelines but not others, necessitating trial and error at each location to achieve the best results (Voordouw et al., 2009). One study even found that nitrates can increase corrosion of steel (Marques, Almeida, Lins, Seldin, & Korenblum, 2012). Finally, even when corrosion inhibition is achieved, for applications such as fuel recovery and processing, the addition of such chemicals then requires subsequent removal processes for the fluid of interest; this is not a treatment localized to the metal surface, and thus the high volume addition and removal of chemical agents affects product cost and thus the decisions to employ such anti-MIC strategies. Thus, although nutrient alteration has produced intriguing results on MIC mitigation, it has yet to produce a definitive solution.
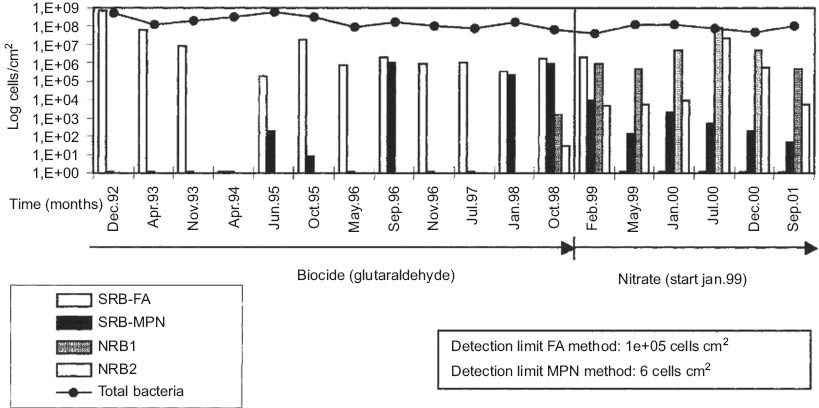
Measured data on the bacteria content of a floating unit at Veslefrikk, an oil production facility in the North Sea. Production began in 1989, utilizing a biocide treatment until January 1999 when it was stopped in favor of nitrate injection. A subsequent decrease in sulfate-reducing bacteria (SRB) and increase in nitrate-reducing bacteria (NRB) numbers was observed. The population of SRB was characterized using a fluorescent antibody (FA) technique as well as the “most probable number” (MPN) method. Two types of NRB were measured using the MPN method: NRB1 which are facultative anaerobes, and NRB2 which are obligate anaerobes (Thorstenson et al., 2002). Reprinted with permission from NACE International.
5 Mechanisms of MIC
Owing to the complexity of the microbe-metal interaction, the underlying mechanism of MIC remains an open question and subject to much debate. Many possible mechanisms for MIC have been proposed, and it is possible that no predominant mechanism exists. In fact, this seems likely given the incredible diversity of species and conditions in which MIC is observed. It is known that chemical, mechanical, and structural factors all play a role, both biotic and abiotic in nature, and these may all work in concert to cause and sustain MIC (Beech & Gaylarde, 1999). Thus, it must be emphasized that evidence for one hypothesis is not necessarily contradictory to another. The given environmental conditions and bacterial species present may dictate the predominant mechanism.
Among the first proposed mechanisms for MIC, the cathodic depolarization theory – also known as the classical theory – was put forth in 1934 by Kuhr and Vlugt in order to explain the unexpectedly high rate of corrosion failures encountered on buried cast iron pipelines in the Dutch countryside (von Wolzogen Kuehr & Van der Vlugt, 1964). SRB were identified as the culprit due to the ferrous sulfide corrosion products found in conjunction with these failures. In anaerobic conditions where SRB thrive, hydrogen ions typically serve as the terminal electron acceptor at the cathode in a corrosion reaction. This reduced hydrogen would adsorb onto the metal surface, polarizing it. According to the classical theory, the role of the SRB was to consume this cathodic hydrogen by means of the enzyme hydrogenase, catalyzing the recombination of the adsorbed atomic hydrogen into hydrogen gas, and thus depolarizing the cathode.
Although the classical theory was grounded in electrochemistry and found general acceptance for nearly half a century, it was ultimately overturned when new techniques showed that hydrogenase can act only on molecular hydrogen and not atomic hydrogen (Stott, 1993). In support of this finding, Cord-Ruwisch (1996) showed that NRB with an enhanced ability to consume hydrogen failed to cause noticeable corrosion. Despite these findings, however, research continues into the role of hydrogenase in MIC, often with contradictory results. Studies by Bryant et al. (1991) and Da Silva, Basséguy, and Bergel (2002) showed a positive correlation between the presence of hydrogenase and MIC, whereas King, Miller, and Wakerley (1973b) found no such link.
Following the fall of the classical theory, myriad “alternative theories” have been proposed, often attempting to explain MIC while minimizing the role of the bacteria themselves (Javaherdashti, 2011). Such theories include anodic depolarization (Crolet, 1992), the presence of a volatile phosphorous compound (Iverson & Olson, 1983), metal-binding exopolymers (Beech & Cheung, 1995), sulfide-induced stress corrosion cracking, and hydrogen-induced blistering (Edyvean, Benson, Thomas, Beech, & Videla, 1998). Here, however, we restrict our focus to the biofilm and its role in MIC. Figure 4 illustrates the mechanisms by which a biofilm can contribute to corrosion initiation and accelerated corrosion rates.
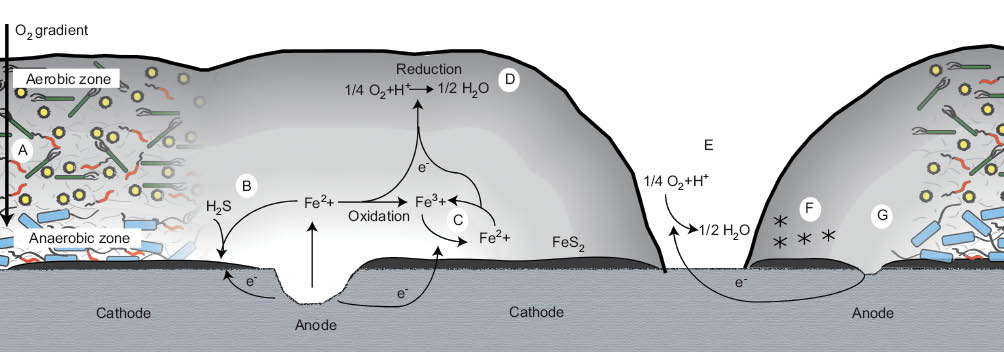
Mechanisms by which a biofilm can contribute to corrosion. (A) Creation of anaerobic zones; (B) concentration of corrosive chemicals; (C) concentration of ferrous ions; (D) electron conduction away from surface; (E) creation of differential aeration zones; (F) binding of corrosion promoters; and (G) disruption of passivating film.
5.1 The biofilm as a chemical barrier
The existence of a biofilm can impede the diffusion of chemical species – both into and out of the biofilm – producing localized chemical environments that are significantly different from the bulk (Beech & Sunner, 2004). These localized environments can, in turn, greatly affect the underlying corrosion in a number of ways (Figure 4). In aerobic conditions, the oxygen concentration may differ greatly across the biofilm due both to O2 diffusion limitations as well as O2 consumption by aerobic bacteria. Thus, even in an aerated environment, anaerobic regions can develop beneath a biofilm, allowing for the proliferation of SRB and other corrosion-enhancing anaerobes (Figure 4A). By inhibiting the diffusion of corrosive chemicals away from the metal surface, the biofilm can effectively act as a concentration multiplier. The H2S generated by SRB in anaerobic regions is potentially much more potent when encapsulated beneath a biofilm than otherwise (Figure 4B). King et al. (1973b) demonstrated a significant difference in the rate of corrosion between an SRB colony and a sterile medium to which an equivalent quantity of sulfide was added periodically. In the former case, the biogenic sulfides were produced beneath a biofilm and near the metal surface, whereas in the latter case the abiotic sulfide was added directly to the bulk solution.
In addition to concentrating the H2S, the biofilm can also concentrate the corroded ferrous ions, a key factor in MIC (Figure 4C). Past studies (King et al., 1973b; Lee & Characklis, 1993) have shown that the presence of ferrous ions is a major factor for MIC to occur on iron or steel in an aqueous environment. Lee & Characklis (1993) grew a pure culture of SRB in the absence of ferrous ions and little to no corrosion was observed despite a high level of microbial activity. Table 1 demonstrates the positive correlation between ferrous ion concentration and corrosion rates. This strong correlation implies that ferrous ions play an integral part in the corrosion mechanism of MIC, at least in the presence of SRB. Two possible roles may be suggested: that the ferrous/ferric ion (Fe2+/Fe3+) redox couple plays a significant role, and/or the ferrous corrosion products do. It should be mentioned that ferrous ions are not known to affect the metabolism of SRB in any manner, although other bacteria are known to use them to cause corrosion on metals other than steel, such as Acidithiobacillus ferrooxidans in the industrial process of bioleaching (Rohwerder, Gehrke, Kinzler, & Sand, 2003).
Relationship between ferrous ion concentration and corrosion rate, adapted from King et al. (1973b).
| Source | Ferrous ion concentration, mm | Average corrosion rate, mg/dm2 day | Standard deviation |
|---|---|---|---|
| Booth & Wormwell (1961) | 0.025 | 4.04 | 1.75 |
| Booth, Shinn, and Wakerley (1964) | 0.125 | 3.0 | – |
| Booth & Wormwell (1961) | 1.25 | 9.5 | 0.3 |
| King, R.A. PhD thesis, University of Manchester (1971) | 5.0 | 11.0 | – |
| Bunker (1939) | 10.7 | 17.5 | 1.5 |
| Booth, Cooper, and Wakerley (1966) | 12.75 | 20.87 | 3.39 |
| King et al. (1973b) | 20 | 21.2 | 2.47 |
| King et al. (1973b) | 38.2 | 24 | – |
| Booth, Cooper, and Cooper (1967) | 510 | 95 | – |
In a metal/solution interface between iron and aqueous ferrous and ferric ion, a galvanic cell is established and the following two half-cell reactions take place:
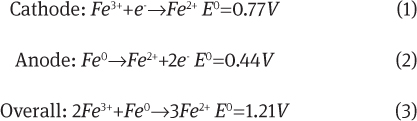
where the standard electrode potentials are referenced to the standard hydrogen electrode. When bound to functional groups in the biofilm, these potentials may become even more conducive towards corrosion, as previously discussed. Nevertheless, as an oxidant, ferric ions (E0=0.77V) are more reducing than hydrogen (E0=0V) but less than oxygen (E0=1.23V), meaning that at the same ion concentrations, a cathodic reaction involving oxygen would be preferred. However, the strength of ferric ions as an oxidant in the corrosion of iron or steel is a positive feedback loop, as illustrated in Figure 4C. Unlike in the oxygen reduction reaction wherein the reactant oxygen molecules are continually consumed – potentially leading to diffusion limiting kinetics – ferrous ions are continually regenerated as this corrosion reaction proceeds. Moreover, each new ferrous ion is generated at the interface, preventing the development of diffusion limited kinetics. These ferrous ions can then be oxidized by oxygen to restore the ferric ion concentration. For pH>4.5, the rate of this reaction is known to be very fast and thus does not limit the rate of corrosion (Singer & Stumm, 1970).
The presence of the biofilm aids this mechanism in two ways: first, it further concentrates the ferric ions near the metal surface; and second, it spatially separates the two oxidation reactions (Beech & Sunner, 2004). Figure 4D illustrates the second point: electrons lost by ferrous ions in the anaerobic zone can travel through the conductive biofilm to be accepted by oxygen in the aerobic zone. By removing the need for either species to diffuse through the biofilm, the rate of the corrosion reaction can thus be greatly increased. This suggests that the presence of the biofilm, rather than the presence of the bacteria that made and stabilized that biofilm, directly increases corrosion rate. Separately, we note that species of iron-oxidizing bacteria, which as part of their metabolism oxidizes Fe2+ to Fe3+, have been identified at MIC events, and these have also been implicated as a potential cause for MIC (Rajasekar et al., 2010).
5.2 Physical structure of the biofilm
The physical structure and heterogeneous nature of the biofilm also contribute to the corrosion process. Biofilms are often patchy and of varying thickness at the microscale, creating isolated pockets of oxygen depletion which can instigate localized corrosion through the establishment of differential aeration cells (Figure 4E) (Picioreanu & van Loosdrecht, 2002). Roe, Lewandowski, and Funk (1996) have demonstrated this effect. They placed a μm-thick patch of agarose – a biogel with an oxygen diffusivity that is slightly lower than that of water – on top of a low-carbon steel coupon, and observed localized corrosion beneath the abiotic film. Additionally, it has been proposed that enzymes embedded within the biofilm – such as hydrogenase – or other chemical properties of the biofilm may also enhance corrosion (Figure 4F) (Beech & Sunner, 2004). The complexation of metal ions by functional groups, for example, is thought to enhance corrosion by altering the redox potential of ferrous ions within the biofilm (Beech & Sunner, 2004), although Roe et al. (1996) found no evidence to support this hypothesis.
In the presence of SRB, ferrous ions will also form iron sulfide corrosion products, the exact form and composition of which are difficult to predict. It is known that iron sulfides can transition from unstable, oxygen-reactive phases such as mackinawite to pyrrhotite and to stable pyrite FeS2 (Rickard & Luther, 2007), but in practice the conversion is spatially and temporally heterogeneous. Cwalina & Dzierzewicz (1999) have noted a difference in the structure of iron sulfides formed biotically versus abiotically, the former referred to as “amorphous precipitates” and the latter as “crystalline sediments.” Regardless, iron sulfides are cathodic towards iron, but this effect may be both protective and destructive (Ma et al., 2000; Stott, 1993; Stumper, 1923). Much like the passive chromium oxide film found on stainless steel, a stable film of iron sulfide may protect the metal surface from further corrosion, but even a small disruption in this film can lead to a high rate of crevice or pitting corrosion due to the massive size disparity between the iron sulfide cathode and the exposed ferrous (iron or steel) anode. Of the various forms of iron sulfide, pyrite (cubic FeS2) is known to be the most stable and also the most corrosive at any sites of film disruption (King, Miller, & Smith, 1973a); FeS2 is also known to be susceptible to dissolution in the presence of certain bacteria such as A. ferrooxidans (Rohwerder et al., 2003).
The effect of a biofilm upon this passive film, however, is subject to much debate. Its presence may stabilize this passive film by acting as a physical binder and a protective coating. By contrast, biofilm presence could potentially inhibit the formation of such a film, or facilitate localized destabilization of such a coating (Figure 4G). Intriguingly, experiments by Lee & Characklis (1993) demonstrate both possibilities, differing only in the order of arrival of the biofilm versus the iron sulfide film. Less well-controlled, industrial environments such as oil pipelines include fluid flow and the continual formation and disruption of both iron sulfides and biofilms. In such contexts, the order of arrival for either film on the ferrous alloy, and thus the effect of the biofilm, remains ambiguous.
6 Summary and outlook
Over decades of research from the perspectives of both corrosion and microbiology, significant progress has been made in understanding MIC. However, this mode of environmentally assisted corrosion remains a pervasive failure mechanism in a wide range of industrial contexts. We know more about the bacterial species and biofilms than ever before, but the complex interplay among species in extreme environments such as anaerobic fluid-metal interfaces has defied effective mitigation strategies. Renewed attention to the biofilm as a biotic coating that can potentially promote and mitigate corrosion of ferrous alloys in sour environments prompts several outstanding questions. In which ways do biotic and abiotic corrosion pits and rates differ? What combination of bacterial species and environmental factors promote MIC? How does fluid flow affect SRB biofilm stability, and how does this depend on the genetic diversity of that species? Answers to questions such as these will require new in situ assays of biotic corrosion, and will allow for improved predictions of environmental conditions susceptible to MIC. Together, these will facilitate cost-effective corrosion-risk monitoring. By approaching this challenging problem of MIC from both a materials engineering and a biological perspective, quantitative insights at the cell-material interface are poised to offer new understanding and mitigation strategies.
About the authors

Kwan Li is a Master’s candidate in Materials Science and Engineering at MIT, working with Van Vliet on microbiologically influenced corrosion via a focus on electrochemistry. Li earned his BS (Duke University, 2011) in Mechanical Engineering and Biomedical Engineering, with a minor in chemistry.

Matthew Whitfield is a postdoctoral research associate in Materials Science and Engineering at MIT, working with Van Vliet. He has developed experimental assays and computational models of bacterial adhesion, including biofilm analysis and corrosion onset in ferrous alloys. Whitfield earned his BE (Vanderbilt University, 2006) in Biomedical Engineering and PhD (University of Washington, 2011) in Bioengineering.

Krystyn J. Van Vliet is the SMART Scholar Associate Professor of Materials Science and Engineering and Biological Engineering at MIT. She and her group study coupling between chemistry and mechanics at material interfaces, including the cell-material interface. As one example, her group studies chemomechanics of abiotic corrosion and biotic corrosion of ferrous alloys in sour environments, via both experiments and simulations. Van Vliet earned her ScB (Brown University, 1998) and PhD (MIT, 2002) in Materials Science and Engineering.
We gratefully acknowledge support from the MIT Energy Initiative (MITEI)-BP Energy Research Fund.
References
Abraham G, Kain V, Dey G. MIC failure of type 316l seawater pipeline. Mater Perform 2009; 48: 64–69.10.5006/MP2009_48_1-64Search in Google Scholar
Akid R, Wang H, Smith T, Greenfield D, Earthman J. Biological functionalization of a sol-gel coating for the mitigation of microbial-induced corrosion. Adv Funct Mater 2008; 18: 203–211.10.1002/adfm.200600493Search in Google Scholar
Al-Jaroudi S, Ul-Hamid A, Al-Gahtani M. Failure of crude oil pipeline due to microbiologically induced corrosion. Corros Eng Sci Technol 2011; 46: 568–579.10.1179/147842210X12695149033819Search in Google Scholar
Amann RI, Ludwig W, Schleifer KH. Phylogenetic identification and in situ detection of individual microbial cells without cultivation. Microbiol Rev 1995; 59: 143–169.10.1128/mr.59.1.143-169.1995Search in Google Scholar PubMed PubMed Central
Andersson S, Kuttuva Rajarao G, Land C, Dalhammar G. Biofilm formation and interactions of bacterial strains found in wastewater treatment systems. FEMS Microbiol Lett 2008; 283: 83–90.10.1111/j.1574-6968.2008.01149.xSearch in Google Scholar PubMed
Avidan O, Satanower S, Banin E. Iron and bacterial biofilm development. Microbial Mats 2010; 14: 359–383.10.1007/978-90-481-3799-2_19Search in Google Scholar
Barton LL, Fauque GD. Biochemistry, physiology and biotechnology of sulfate-reducing bacteria. Adv Appl Microbiol 2009; 68: 41–98.10.1016/S0065-2164(09)01202-7Search in Google Scholar PubMed
Beech I, Cheung CS. Interactions of exopolymers produced by sulphate-reducing bacteria with metal ions. Int Biodeterior Biodegrad 1995; 35: 59–72.10.1016/0964-8305(95)00082-GSearch in Google Scholar
Beech IB, Gaylarde CC. Recent advances in the study of biocorrosion: an overview. Rev Microbiol 1999; 30: 117–190.10.1590/S0001-37141999000300001Search in Google Scholar
Beech IB, Sunner J. Biocorrosion: towards understanding interactions between biofilms and metals. Curr Opin Biotechnol 2004; 15: 181–186.10.1016/j.copbio.2004.05.001Search in Google Scholar PubMed
Booth G, Wormwell F. Corrosion of mild steel by sulphate-reducing bacteria. Effect of different strains of organisms. In: First International Congress on Metallic Corrosion, London, 1961: 341–344.Search in Google Scholar
Booth G, Shinn P, Wakerley D. The influence of various strains of actively growing sulphate-reducing bacteria on the anaerobic corrosion of mild steel. In: Congres Int de la Corrosion Marine et des Salissures, Cannes, volume 363, 1964.Search in Google Scholar
Booth G, Cooper PM, Wakerley D. Corrosion of mild steel by actively growing cultures of sulphate-reducing bacteria. The influence of ferrous iron. Br Corros J 1966; 1: 345–349.10.1179/000705966798327254Search in Google Scholar
Booth G, Cooper A, Cooper P. Rates of microbial corrosion in continuous culture. Chem Ind 1967; 9: 2084–2085.Search in Google Scholar
Bryant RD, Jansen W, Boivin J, Laishley EJ, Costerton JW. Effect of hydrogenase and mixed sulfate-reducing bacterial populations on the corrosion of steel. Appl Environ Microbiol 1991; 57: 2804–2809.10.1128/aem.57.10.2804-2809.1991Search in Google Scholar PubMed PubMed Central
Bunker H. Microbiological experiments in anaerobic corrosion. J Soc Chem Ind 1939; 58: 93.Search in Google Scholar
Chamritski I, Burns G, Laycock N, Webster B. Effect of iron-oxidizing bacteria on pitting of stainless steel. Corrosion 2004; 60: 658–669.10.5006/1.3287842Search in Google Scholar
Cord-Ruwisch R. MIC in hydrocarbon transportation systems. Corros Australasia 1996; 21: 8–11.Search in Google Scholar
Crolet J-L. From biology and corrosion to biocorrosion. Oceanol Acta 1992; 15: 87–94.Search in Google Scholar
Cwalina O, Dzierzewicz Z. The influence of sulfur oxy-anions on the rate of biocorrosion of metals. Eur Fed Corros Publ 1999; 29: 47–52.Search in Google Scholar
Da Silva S, Basséguy R, Bergel A. The role of hydrogenases in the anaerobic microbiologically influenced corrosion of steels. Bioelectrochemistry 2002; 56: 77–79.10.1016/S1567-5394(02)00034-8Search in Google Scholar PubMed
Dexter S, Eashwar M. Relation of bacterial settlement patterns to anodic activity on stainless steel weldments. Corrosion 1999; 99: 1–19.10.5006/C1999-99174Search in Google Scholar
Dong Z, Shi W, Ruan H, Zhang G. Heterogeneous corrosion of mild steel under SRB-biofilm characterised by electrochemical mapping technique. Corros Sci 2011; 53: 2978–2987.10.1016/j.corsci.2011.05.041Search in Google Scholar
Dubiel M, Hsu CH, Chien CC, Mansfeld F, Newman DK. Microbial iron respiration can protect steel from corrosion. Appl Environ Microbiol 2002; 68: 1440–1445.10.1128/AEM.68.3.1440-1445.2002Search in Google Scholar PubMed PubMed Central
Dunsmore B, Jacobsen A, Hall-Stoodley L, Bass C, Lappin-Scott H, Stoodley P. The influence of fluid shear on the structure and material properties of sulphate-reducing bacterial biofilms. J Ind Microbiol Biotechnol 2002; 29: 347–353.10.1038/sj/jim/7000302Search in Google Scholar
Edyvean R, Benson I, Thomas C, Beech I, Videla H. Biological influences on hydrogen effects in steel in seawater. Mater Perform 1998; 37: 40–44.Search in Google Scholar
Egan M. Internal corrosion suspected as cause of Alaskan pipeline leak. Mater Perform 2011; 50: 14–23.10.5006/MP2011_50_5-14Search in Google Scholar
Fernandez N, Diaz EE, Amils R, Sanz JL. Analysis of microbial community during biofilm development in an anaerobic wastewater treatment reactor. Microb Ecol 2008; 56: 121–132.10.1007/s00248-007-9330-2Search in Google Scholar PubMed
Flemming HC. Economical and technical overview. Berlin: Springer-Verlag, 1996.10.1007/978-3-642-80017-7_2Search in Google Scholar
George R. Current understanding and future approaches for controlling microbially influenced concrete corrosion: a review. Concr Res Lett 2012; 3: 491–506.Search in Google Scholar
Haveman SA, Greene EA, Stilwell CP, Voordouw JK, Voordouw G. Physiological and gene expression analysis of inhibition of Desulfovibrio vulgaris Hildenborough by nitrite. J Bacteriol 2004; 186: 7944–7950.10.1128/JB.186.23.7944-7950.2004Search in Google Scholar PubMed PubMed Central
He Q, He Z, Joyner D, Joachimiak M, Price M, Yang Z, Yen H, Hemme C, Chen W, Fields M. Impact of elevated nitrate on sulfate-reducing bacteria: a comparative study of Desulfovibrio vulgaris. ISME J 2010; 4: 1386–1397.10.1038/ismej.2010.59Search in Google Scholar PubMed
Hubert C, Voordouw G. Oil field souring control by nitrate-reducing Sulfurospirillum spp. that outcompete sulfate-reducing bacteria for organic electron donors. Appl Environ Microbiol 2007; 73: 2644–2652.10.1128/AEM.02332-06Search in Google Scholar PubMed PubMed Central
Iverson W, Olson G. Anaerobic corrosion by sulfate-reducing bacteria due to a highly-reactive volatile phosphorus compound. Final report. Technical report. Washington, DC: National Bureau of Standards, 1983: 46–53.Search in Google Scholar
Jan-Roblero J, Romero J, Amaya M, Le Borgne S. Phylogenetic characterization of a corrosive consortium isolated from a sour gas pipeline. Appl Microbiol Biotechnol 2004; 64: 862–867.10.1007/s00253-004-1613-7Search in Google Scholar PubMed
Javaherdashti R. Impact of sulphate-reducing bacteria on the performance of engineering materials. Appl Microbiol Biotechnol 2011; 91: 1507–1517.10.1007/s00253-011-3455-4Search in Google Scholar PubMed
Johnson MS, Zhulin IB, Gapuzan M, Taylor BL. Oxygen-dependent growth of the obligate anaerobe Desulfovibrio vulgaris Hildenborough. J Bacteriol 1997; 179: 5598–5601.10.1128/jb.179.17.5598-5601.1997Search in Google Scholar PubMed PubMed Central
King R, Miller J, Smith J. Corrosion of mild steel by iron sulphides. Br Corros J 1973a; 8: 137–141.10.1179/000705973798322251Search in Google Scholar
King R, Miller J, Wakerley D. Corrosion of mild steel in cultures of sulphate-reducing bacteria: effect of changing the soluble iron concentration during growth. Br Corros J 1973b; 8: 89–93.10.1179/000705973798322396Search in Google Scholar
Koch G, Brongers M, Thompson N, Virmani Y, Payer J. Corrosion cost and preventive strategies in the United States. Technical report. McLean, VA: Turner-Fairbank Highway Research Center, 2002: E5–E6.Search in Google Scholar
Larsen J, Rasmussen K, Pedersen H, Sarensen K, Lundgaard T, Skovhus TL. Consortia of MIC bacteria and archaea causing pitting corrosion in top side oil production facilities. In: Corrosion 2010, March 14–18, 2010, San Antonio, TX, USA. Houston, TX: NACE International, 2010: 10252.10.5006/C2010-10252Search in Google Scholar
Lee W, Characklis W. Corrosion of mild steel under anaerobic biofilm. Corrosion 1993; 49: 186–199.10.5006/1.3316040Search in Google Scholar
Lee A, Buehler M, Newman D. Influence of a dual-species biofilm on the corrosion of mild steel. Corros Sci 2006; 48: 165–178.10.1016/j.corsci.2004.11.013Search in Google Scholar
Lichter JA, Thompson MT, Delgadillo M, Nishikawa T, Rubner MF, Van Vliet KJ. Substrata mechanical stiffness can regulate adhesion of viable bacteria. Biomacromolecules 2008; 9: 1571–1578.10.1021/bm701430ySearch in Google Scholar PubMed
Lichter JA, Van Vliet KJ, Rubner MF. Design of antibacterial surfaces and interfaces: polyelectrolyte multilayers as a multifunctional platform. Macromolecules 2009; 42: 8573.10.1021/ma901356sSearch in Google Scholar
Little B, Ray R, Wagner P, Jones-Meehan J, Lee C, Mansfeld F. Spatial relationships between marine bacteria and localized corrosion on polymer coated steel. Biofouling 1999; 13: 301–321.10.1080/08927019909378387Search in Google Scholar
Ma H, Cheng X, Li G, Chen S, Quan Z, Zhao S, Niu L. The influence of hydrogen sulfide on corrosion of iron under different conditions. Corros Sci 2000; 42: 1669–1683.10.1016/S0010-938X(00)00003-2Search in Google Scholar
Marques J, Almeida F, Lins U, Seldin L, Korenblum E. Nitrate treatment effects on bacterial community biofilm formed on carbon steel in produced water stirred tank bioreactor. World J Microbiol Biotechnol 2012; 28: 2355–2363.10.1007/s11274-012-1043-0Search in Google Scholar PubMed
Maruthamuthu S, Kumar BD, Ramachandran S, Anandkumar B, Palanichamy S, Chandrasekaran M, Subramanian P, Palaniswamy N. Microbial corrosion in petroleum product transporting pipelines. Ind Eng Chem Res 2011; 50: 8006–8015.10.1021/ie1023707Search in Google Scholar
Maxwell S. Predicting microbially influenced corrosion in seawater injection systems. In: SPE International Oilfield Corrosion Symposium, 30 May 2006, Aberdeen, UK. Richardson, TX: Society of Petroleum Engineers, 2006: 100519-MS.10.2118/100519-MSSearch in Google Scholar
McElhiney J, Burger E, Maxwell S, Davis R, Walsh J. Adoption of a process to prevent souring associated with seawater waterflooding in the Ursa-Princess fields, deepwater Gulf of Mexico. In: SPE International Oilfield Corrosion Conference, 27 May 2008, Aberdeen, UK. Richardson, TX: Society of Petroleum Engineers, 2008: 113317-MS.Search in Google Scholar
Miranda E, Bethencourt M, Botana F, Cano M, Sanchez-Amaya J, Corzo A, De Lomas J, Fardeau M, Ollivier B. Biocorrosion of carbon steel alloys by an hydrogenotrophic sulfate-reducing bacterium Desulfovibrio capillatus isolated from a Mexican oil field separator. Corros Sci 2006; 48: 2417–2431.10.1016/j.corsci.2005.09.005Search in Google Scholar
Mohanakrishnan J, Kofoed M, Barr J, Yuan Z, Schramm A, Meyer R. Dynamic microbial response of sulfidogenic wastewater biofilm to nitrate. Appl Microbiol Biotechnol 2011; 91: 1647–1657.10.1007/s00253-011-3330-3Search in Google Scholar PubMed
Monds RD, O’Toole GA. The developmental model of microbial biofilms: ten years of a paradigm up for review. Trends Microbiol 2009; 17: 73–87.10.1016/j.tim.2008.11.001Search in Google Scholar PubMed
Okabe S, Itoh T, Satoh H, Watanabe Y. Analyses of spatial distributions of sulfate-reducing bacteria and their activity in aerobic wastewater biofilms. Appl Environ Microbiol 1999; 65: 5107–5116.10.1128/AEM.65.11.5107-5116.1999Search in Google Scholar PubMed PubMed Central
Picioreanu C, van Loosdrecht MC. A mathematical model for initiation of microbiologically influenced corrosion by differential aeration. J Electrochem Soc 2002; 149: B211–B223.10.1149/1.1470657Search in Google Scholar
Rajasekar A, Anandkumar B, Maruthamuthu S, Ting Y-P, Rahman PK. Characterization of corrosive bacterial consortia isolated from petroleum-product-transporting pipelines. Appl Microbiol Biotechnol 2010; 85: 1175–1188.10.1007/s00253-009-2289-9Search in Google Scholar PubMed
Rao T. Microbial fouling and corrosion: fundamentals and mechanisms. In: Operational and environmental consequences of large industrial cooling water systems. New York: Springer US, 2012: 95–126.10.1007/978-1-4614-1698-2_6Search in Google Scholar
Rickard D, Luther III GW. Chemistry of iron sulfides. Chem Rev 2007; 107: 514–562.10.1021/cr0503658Search in Google Scholar PubMed
Rochex A, Godon J, Bernet N, Escudie R. Role of shear stress on composition, diversity and dynamics of biofilm bacterial communities. Water Res 2008; 42: 4915–4922.10.1016/j.watres.2008.09.015Search in Google Scholar PubMed
Roe F, Lewandowski Z, Funk T. Simulating microbiologically influenced corrosion by depositing extracellular biopolymers on mild steel surfaces. Corrosion 1996; 52: 744–752.10.5006/1.3292067Search in Google Scholar
Rohwerder T, Gehrke T, Kinzler K, Sand W. Bioleaching review. Part A. Appl Microbiol Biotechnol 2003; 63: 239–248.10.1007/s00253-003-1448-7Search in Google Scholar PubMed
Schwermer C, Lavik G, Abed R, Dunsmore B, Ferdelman T, Stoodley P, Gieseke A, De Beer D. Impact of nitrate on the structure and function of bacterial biofilm communities in pipelines used for injection of seawater into oil fields. Appl Environ Microbiol 2008; 74: 2841–2851.10.1128/AEM.02027-07Search in Google Scholar PubMed PubMed Central
Sherar B, Power I, Keech P, Mitlin S, Southam G, Shoesmith D. Characterizing the effect of carbon steel exposure in sulfide containing solutions to microbially induced corrosion. Corros Sci 2011; 53: 955–960.10.1016/j.corsci.2010.11.027Search in Google Scholar
Singer PC, Stumm W. Acidic mine drainage: the rate-determining step. Science 1970; 167: 1121.10.1126/science.167.3921.1121Search in Google Scholar PubMed
Skovhus T, Holmkvist L, Andersen K, Larsen J, Pedersen H. MIC risk assessment of the Halfdan Oil Export Spool. In: SPE International Conference and Exhibition on Oilfield Corrosion, 28–29 May 2012, Aberdeen, UK. Richardson, TX: Society of Petroleum Engineers, 2012: 155080-MS.10.2118/155080-MSSearch in Google Scholar
Sreekumari K, Nandakumar K, Kikuchi Y. Bacterial attachment to stainless steel welds: significance of substratum microstructure. Biofouling 2001; 17: 303–316.10.1080/08927010109378490Search in Google Scholar
Stadler R, Wei L, Furbeth W, Grooters M, Kuklinski A. Influence of bacterial exopolymers on cell adhesion of Desulfovibrio vulgaris on high alloyed steel: corrosion inhibition by extracellular polymeric substances (EPS). Mater Corros 2010; 61: 1008–1016.10.1002/maco.201005819Search in Google Scholar
Stoodley P, Jacobsen A, Dunsmore BC, Purevdorj B, Wilson S, Lappin-Scott HM, Costerton JW. The influence of fluid shear and AICI3 on the material properties of Pseudomonas aeruginosa PAO1 and Desulfovibrio sp. EX265 biofilms. Water Sci Technol 2001; 43: 113–120.10.2166/wst.2001.0353Search in Google Scholar
Stott J. What progress in the understanding of microbially induced corrosion has been made in the last 25 years? A personal viewpoint. Corros Sci 1993; 35: 667–673.10.1016/0010-938X(93)90202-RSearch in Google Scholar
Stumper R. La corrosion du fer en presence du sulfure de fer. Compt Rend Acad Sci (Paris) 1923; 176: 1316.Search in Google Scholar
Thomas L, Chung Y. Comparison of MIC pit morphology with chloride induced pits in types 304/304l/E308 stainless steel base metal/welds. In: Corrosion 1999, April 25–30, 1999, San Antonio, TX, USA. Houston, TX: NACE International, 1999: 99159.10.5006/C1999-99159Search in Google Scholar
Thomas WE, Trintchina E, Forero M, Vogel V, Sokurenko EV. Bacterial adhesion to target cells enhanced by shear force. Cell 2002; 109: 913–923.10.1016/S0092-8674(02)00796-1Search in Google Scholar PubMed
Thorstenson T, Torsvik T, Beeder J, Lillebo B-L, Sunde E, Bodtker G. Biocide replacement by nitrate in sea water injection systems. In: Corrosion 2002, April 7–11, 2002, Denver, CO, USA. Houston, TX: NACE International, 2002: 02033.10.5006/C2002-02033Search in Google Scholar
Videla H, Borgne S, Panter C, Singh Raman R. MIC of steels by iron reducing bacteria. Corrosion 2008, March 16–20, 2008, New Orleans, LA, USA. Houston, TX: NACE International, 2008: 08505.10.5006/C2008-08505Search in Google Scholar
von Wolzogen Kuehr C, Van der Vlugt L. Graphitization of cast iron as an electrobiochemical process in anaerobic soils. Technical report. DTIC document. Fredrick, MD: US Army Biological Laboratories, 1964.Search in Google Scholar
Voordouw G, Grigoryan AA, Lambo A, Lin S, Park HS, Jack TR, Coombe D, Clay B, Zhang F, Ertmoed R, Miner K, Arensdorf JJ. Sulfide remediation by pulsed injection of nitrate into a low temperature Canadian heavy oil reservoir. Environ Sci Technol 2009; 43: 9512–9518.10.1021/es902211jSearch in Google Scholar PubMed
Vu B, Chen M, Crawford R, Ivanova E. Bacterial extracellular polysaccharides involved in biofilm formation. Molecules 2009; 14: 2535–2554.10.3390/molecules14072535Search in Google Scholar PubMed PubMed Central
Wall J, Arkin A, Balci N, Rapp-Giles B. Genetics and genomics of sulfate respiration in desulfovibrio. In: Dahl C, Friedrich C, editors. Microbial sulfur metabolism. Berlin: Springer, 2008: 1–12.10.1007/978-3-540-72682-1_1Search in Google Scholar
Wen J, Gu T, Nesic S. Investigation of the effects of fluid flow on SRB biofilm. In: Corrosion 2007, March 11–15, 2007, Nashville, TN, USA. Houston, TX: NACE International Corrosion 2007: 07516.10.5006/C2007-07516Search in Google Scholar
Whitfield M, Ghose T, Thomas W. Shear-stabilized rolling behavior of E. coli examined with simulations. Biophys J 2010; 99: 2470–2478.10.1016/j.bpj.2010.08.045Search in Google Scholar PubMed PubMed Central
Yuan S, Pehkonen S. AFM study of microbial colonization and its deleterious effect on 304 stainless steel by Pseudomonas NCIMB 2021 and Desulfovibrio desulfuricans in simulated seawater. Corros Sci 2009; 51: 1372–1385.10.1016/j.corsci.2009.03.037Search in Google Scholar
Yu F, Dillon J, Henry T. Identification and control of microbiologically influenced corrosion in a power plant. In: Corrosion 2010, March 14–18, 2010, San Antonio, TX, USA. Houston, TX: NACE International, 2010: 10211.10.5006/C2010-10211Search in Google Scholar
Zuo R. Biofilms: strategies for metal corrosion inhibition employing microorganisms. Appl Microbiol Biotechnol 2007; 76: 1245–1253.10.1007/s00253-007-1130-6Search in Google Scholar PubMed
©2013 by Walter de Gruyter Berlin Boston
Articles in the same Issue
- Masthead
- Masthead
- Graphical abstract
- Graphical abstract
- Reviews
- Beating the bugs: roles of microbial biofilms in corrosion
- A critical review of the influence of hydrogen on the mechanical properties of medium-strength steels
- In situ remediation of leaks in potable water supply systems
- A state-of-the-art review on the durability of silica fume-blended concrete – a boon to the construction industry
- Original article
- Corrosion rate of carbon steel under synergistic effect of seawater parameters including pH, temperature, and salinity in turbulent condition
Articles in the same Issue
- Masthead
- Masthead
- Graphical abstract
- Graphical abstract
- Reviews
- Beating the bugs: roles of microbial biofilms in corrosion
- A critical review of the influence of hydrogen on the mechanical properties of medium-strength steels
- In situ remediation of leaks in potable water supply systems
- A state-of-the-art review on the durability of silica fume-blended concrete – a boon to the construction industry
- Original article
- Corrosion rate of carbon steel under synergistic effect of seawater parameters including pH, temperature, and salinity in turbulent condition