Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
-
Giuliana Marletta
Giuliana Marletta has a PhD in Earth and Environmental Sciences at the University of Catania. Her research expertise is the field of marine ecology focusing particularly on conservation and ecological restoration of Mediterranean underwater forests. Furthermore, her research covers studies on the dynamics of benthic vegetation.
Andrea Lombardo is a PhD student at the Department of Biological, Geological and Environmental Sciences, University of Catania. His research concerns the study of Sicilian marine heterobranch (sea slugs) faunas. He is also interested and actively collaborates in studies on brown algae belonging to the order Fucales.
Donatella Serio is a researcher at the University of Catania, Italy. Her current research interests include marine benthic flora and vegetation. The floristic and vegetational searches deal with areas previously studied to estimate changes of biodiversity at both short and long term. In parallel with the above-mentioned research, she is carrying out studies aiming to clarify the taxonomy of some genera of marine macroalgae.



Abstract
The Egadi Islands Marine Protected Area (MPA) (west of Sicily) is one of the largest MPAs in the Mediterranean Sea. Nevertheless, studies of benthic macroalgae in this MPA are few and quite outdated. The latest study, performed about 20 years ago, documented the disappearance of cold affinity species (mainly Cystoseira s.l. and Sargassum species) in favour of warm affinity ones. In this study, we conducted a visual census activity through snorkelling and scuba dives on the island of Marettimo, and we found a total of 17 Fucales: four belonging to Cystoseira, eight to Ericaria, two to Gongolaria, and three to Sargassum. The obtained data were compared with previous studies conducted in the 1970s and in the early 2000s in the E-MPA reporting data on the Fucales stands to understand the dynamics that have occurred during this period. Five species, not previously reported from the island of Marettimo, were found during this study: Cystoseira cf micheleae, Ericaria barbatula, Ericaria brachycarpa var. claudiae, Gongolaria montagnei var. compressa and Sargassum cf furcatum.
1 Introduction
Marine Protected Areas (MPAs) are considered important tools to protect the natural capital from different anthropogenic threats on marine ecosystems and ensure the proper management of human uses in the maritime domain (D’Anna et al. 2016; Rasheed 2020). MPAs are characterized by the interconnection between the social, economic, and ecological dynamics, representing complex conservation tools to achieve sustainability goals. Many studies demonstrated that MPAs can contribute to biodiversity protection, ensuring the sustainable exploitation of marine resources (Halpern 2003; OECD 2017).
The Egadi Islands Marine Protected Area (Central Mediterranean, Italy; hereinafter referred to as “E-MPA”) is one of the largest MPAs in the Mediterranean Sea, covering 53,992 ha and 74 km of coastline (D’Anna et al. 2016). This MPA is located at about 7–9 km from the western coast of Sicily (Figure 1A and B), across the Sicily Channel and the Tyrrhenian Sea, and comprises three main islands (Favignana, Marettimo, and Levanzo) and a few small rocky outcrops (Galeotta, Galera, Preveto, Formica, and Maraone) (Mannino et al. 2017). The E-MPA is partitioned into four management zones (Figure 1C) (D’Anna et al. 2016; Guidetti et al. 2008): Zone A – fully protected zone, where only bathing and scientific research are allowed; Zone B – buffer zone, where bathing, free sailing, rowing, pedal boats or electric propulsion, guided diving tours conducted by resident diving centres and small-scale fishing gears are allowed; Zone C – peripheral zone, in which bathing, free navigation, free anchorage outside the environmentally relevant seabed, guided diving tours conducted by resident diving centres and stipulated selective fishing gears, sport fishing and small-scale professional fishing. are permitted; Zone D – regulated trawling zone, where all legal fishery activities are permitted, including trawl fishing and large-scale purse seine fishing.

Map of the Egadi Islands Marine Protected Area (E-MPA) and investigated sites in the Island of Marettimo. (A, B) Location of the E-MPA in the Mediterranean Sea. (C) Zoning scheme of the E-MPA. (D) Marettimo Island with investigated sites (red circles).
Very few studies have been undertaken on the benthic macroalgae of the E-MPA. The first data on the flora and vegetation of this area were provided by Giaccone (1970), Giaccone and Sortino (1974), Sortino et al. (1976) and Giaccone et al. (1993). The latest study on the benthic algal flora of this MPA was performed about 20 years ago by Catra et al. (2006), who reported an increase of warm affinity species (mainly Rhodophyta) and a decrease of cold affinity species (mainly Phaeophyceae) compared to previous data. In recent years, Mannino et al. (2016, 2017) and Mannino and Balistreri (2017, 2018) analysed the invasive alien species (IAS) of the E-MPA and promoted citizen science (e.g., project Caulerpa cylindracea Sonder) as a useful tool to monitor the spread dynamics of these species in the E-MPA. The finding of invasive alien taxa and observation of an increasing number of warm water affinity species was associated with the process of tropicalization and seawater warming (Furnari and Cormaci 2009). Furthermore, although most of the communities present in the 1970s were still well structured, Catra et al. (2006) had already reported the loss of some canopy-forming species of Fucales, in particular those belonging to Cystoseira sensu lato (s.l.) (comprising the genera Cystoseira C. Agardh, Ericaria Stackhouse and Gongolaria Boehmer) and Sargassum C. Agardh).
In recent years, most members of Fucales have suffered a massive decline in many Mediterranean areas due to anthropogenic activities, exacerbated by the ongoing climate change (Thibaut et al. 2015). Notwithstanding their fundamental role as habitat-forming species and ecosystem engineers in Mediterranean waters (Sales et al. 2011; Thibaut et al. 2005), to date the current presence, distribution and conservation status of these Fucales is surprisingly limited and ecological analyses for many areas are still rare (Mancuso et al. 2018; Neiva et al. 2022). Information on these communities in the E-MPA is quite outdated and scarce. Therefore, this study aimed to obtain an updated knowledge of the presence and distribution of Fucales around one of the islands of the E-MPA (Marettimo), about 20 years after the latest study carried out in this area.
The obtained data were compared with three previous studies conducted in the E-MPA reporting data on the Fucales stands: Giaccone (1970), Giaccone and Sortino (1974), and Catra et al. (2006) to understand the dynamics that have occurred during this period.
2 Materials and methods
During the second week of July 2024, a visual census activity was carried out around the island of Marettimo (Figure 1D), the most distant of the islands belonging to the E-MPA (Central Mediterranean; the only island presenting Zone A – fully protected zone). A total of 18 sites (Figure 1D; Table S1), covering almost all the island’s boundaries, were surveyed through snorkelling and scuba diving activities. In particular, 11 scuba dives and seven snorkelling activities were performed, each lasting from 40 min to 1 h, covering an area of approximately 250 m2 per dive. The visual census activity was conducted through the technique of random courses, which allows for the exploration of different environments and depths (Harmelin-Vivien et al. 1985; La Mesa et al. 2017; Marletta and Lombardo 2020). The dives were carried out perpendicular to the coastline, following the geomorphology of the seabed from the surface to a depth of around 45 m, depending on the site, while the snorkelling activities were performed along the coastline following the same depth range (0–1.5 m).
During the visual census activity, Cystoseira s.l. and Sargassum species were photographed using two underwater cameras, Olympus TG-6 and TG-4 (Olympus Corporation, Tokyo, Japan). The identification was made in situ and/or on photographic documentation. This approach has recently proved to be useful for the identification of these endangered species, reducing the sampling effort on them (Marletta and Lombardo 2023; Marletta et al. 2023) especially in marine protected areas. For each species, photographs were taken of the frond, base, apex, and the receptacles and tophules, if present. Species identification was performed according to the identification keys by Gómez-Garreta et al. (2001), Cormaci et al. (2012), Rodríguez-Prieto et al. (2013), Blanfuné et al. (2022) and the most recent taxonomic studies. The canopy density of each species was estimated in situ according to the classes proposed by Thibaut et al. (2016): absence of individuals (1), scattered individuals (2), abundant patches (3), almost continuous to continuous stands (4).
All taxa at specific and infraspecific level identified (hereinafter referred to as species) are listed in alphabetical order in Table S2, which also provides information on the range of depths at which species were documented and canopy density. Finally, the presence of mature individuals was also reported. For the taxonomy and nomenclature of species, we followed AlgaeBase (Guiry and Guiry 2024).
The obtained data were compared with previous studies conducted in the E-MPA (Table 1). Such reports were corrected according to modern taxonomy and nomenclature for comparison.
Fucalean species found in the E-MPA (Egadi Islands Marine Protected Area): comparison of the present study with the previous studies.
| Species found in the E-MPA | Giaccone (1970) | Giaccone and Sortino (1974) | Catra et al. (2006) | Present study |
|---|---|---|---|---|
| Cystoseira compressa | F, L, M (as Cystoseira fimbriata) | F, M (as Cystoseira fimbriata) | F, L, M (as Cystoseira compressa) | X |
| Cystoseira foeniculacea f. latiramosa | F, L, M (as Cystoseira discors subsp. latiramosa) | F, L, M (as Cystoseira discors subsp. latiramosa) | ||
| C. foeniculacea f. tenuiramosa | F, L, M (as Cystoseira ercegovicii f. tenuiramosa) | M (as Cystoseira foeniculacea f. tenuiramosa) | X | |
| Cystoseira cf. micheleae | X | |||
| Cystoseira pustulata | F, L, M (as Cystoseira myriophylloides var. humilis) | F, L (as Cystoseira humilis) | X | |
| Cystoseira platyclada Sauvageau (taxon inquirendum) | M | |||
| Ericaria amentacea | F, L, M (as Cystoseira stricta) | F, M (as Cystoseira stricta) | F, L, M (as Cystoseira amentacea var. stricta) | X |
| Ericaria barbatula | X | |||
| Ericaria brachycarpa v. brachycarpa | F, L, M (as Cystoseira balearica) | F, L, M (as Cystoseira brachycarpa) | X | |
| Ericaria brachycarpa v. claudiae | X | |||
| Ericaria crinita | F, L, M (as Cystoseira crinita) | F, L, M (as Cystoseira stricta) | X | |
| Ericaria dubia | F (as Cystoseira dubia) | X | ||
| Ericaria mediterranea | M (as Cystoseira mediterranea) | |||
| Ericaria zosteroides | L, M (as Cystoseira zosteroides) | F, M (as Cystoseira zosteroides) | F (as Cystoseira zosteroides) | X |
| Ericaria funkii | F, M (as Cystoseira jabukae) | F, L, M (as Cystoseira funkii) | X | |
| Gongolaria barbata | F, M (as Cystoseira barbata) | |||
| Gongolaria elegans | M (as Cystoseira elegans) | |||
| Gongolaria montagnei v. montagnei | F, L, M (as Cystoseira spinosa v. spinosa) | F, L, M (as Cystoseira spinosa v. spinosa) | F, L, M (as Cystoseira spinosa v. spinosa) | |
| Gongolaria montagnei v. compressa | X | |||
| Gongolaria montagnei var. tenuior | F (as Cystoseira spinosa var. tenuior) | |||
| Gongolaria sauvageauana | F, M (as Cystoseira sauvageauana) | X | ||
| Sargassum acinarium | F, M (as Sargassum acinarium) | X | ||
| Sargassum cf furcatum | X | |||
| Sargassum hornschuchii | F, L, M (as S. hornschuchii) | F, L, M (as Sargassum hornschuchii) | ||
| Sargassum vulgare | M (as Sargassum vulgare) | F, M (as Sargassum vulgare) | F, L, M (as Sargassum vulgare) | X |
| Total (E-MPA) | 9 | 18 | 10 | |
| Total (Marettimo Island) | 9 | 17 | 7 | 17 |
-
Letters F, L, M indicate the islands where the species was found in the past (F, Favignana; L, Levanzo; M, Marettimo) with name under which species was cited in brackets. X in final column indicates that species was found in the present study.
3 Results
During the visual census activity, a total of 17 species were found: four belonging to Cystoseira (C. compressa, C. foeniculacea f. tenuiramosa, C. pustulata and C. cf micheleae), eight to Ericaria (E. amentacea, E. barbatula, E. brachycarpa, E. brachycarpa var. claudiae, E. crinita, E. dubia, E. funkii and E. zosteroides), two to Gongolaria (G. montagnei var. compressa and G. sauvageauana) and three to Sargassum (S. acinarium, S. cf furcatum and S. vulgare) (Table 1).
During this study, it was observed that the areas presenting the highest number of species are located along the southern side of the island, in Zone B, while the sites located in the eastern side of the island, corresponding to Zone C, are those presenting the lowest number of species (Table S2). The western and northern sides of the island, including Zone A and another part of Zone B, show intermediate values compared to the southern coast and the eastern one (Table S2).
Comparing the present data with the study of Giaccone (1970), it was observed that, of the nine reported species of Fucales, four are still present today (C. compressa, E. amentacea, E. zosteroides and S. vulgare). Later, Giaccone and Sortino (1974) found a total of 18 species of Fucales in the Egadi Islands (17 at Marettimo Island). Of them, 12 were found in the present study: C. compressa, C. foeniculacea f. tenuiramosa, C. pustulata, E. amentacea, E. brachycarpa, E. crinita, E. dubia, E. zosteroides, E. funkii, G. sauvageauana, S. acinarium and S. vulgare. In 2001, Catra et al. (2006) found 10 fucalean species (7 at Marettimo Island). Of them, eight were also found during the present study (C. compressa, C. foeniculacea f. tenuiramosa, C. pustulata, E. amentacea, E. brachycarpa, E. zosteroides, E. funkii and S. vulgare) (Table 1).
Five species, not previously reported from the island of Marettimo, were found during this study: C. cf micheleae, E. barbatula, E. brachycarpa var. claudiae, G. montagnei var. compressa and S. cf furcatum.
For each species we found at Marettimo, details of taxonomy, morphological description, local habitat and distribution, as well as further remarks are given below.
Cystoseira compressa (Esper) Gerloff et Nizamuddin (Figure 2).
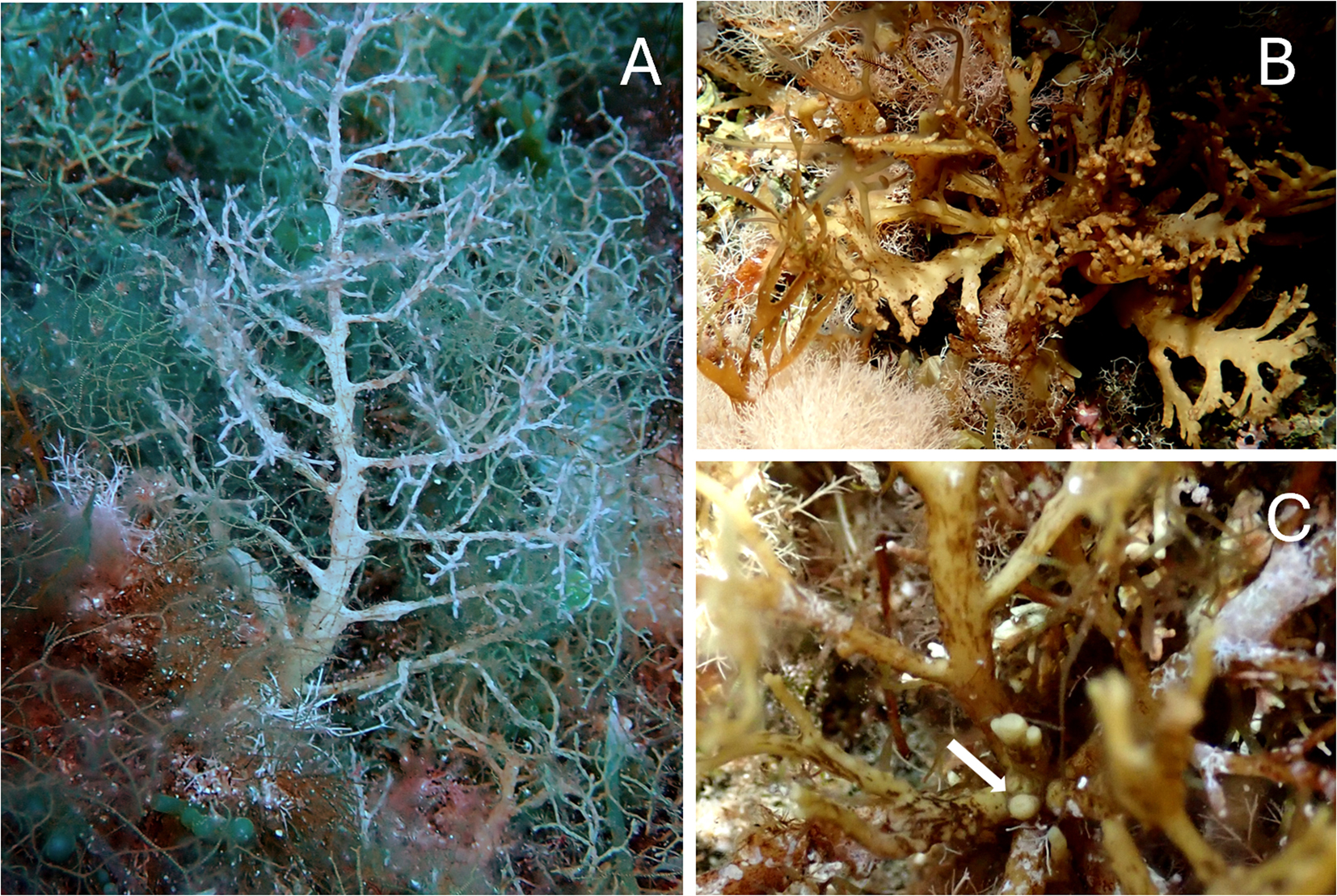
Cystoseira compressa. (A) Habit in deeper waters; (B) habit in exposed and shallow waters; (C) smooth apex (white arrow).
Morphological description of specimens: thalli are caespitose, with several flattened axes originating from a small discoid base. Primary branches are flattened with alternate–distichous branching (Figure 2A). Higher order branches can be flattened or cylindrical but with the same arrangement of primary branches. Thalli can show different habits depending on the environment: in sheltered or deeper waters thalli are taller (Figure 2A), while in more exposed and shallow waters they are small-sized (Figure 2B). The apex is smooth and not prominent (Figure 2C). Specimens with aerocysts were never observed during this study. Receptacles are tiny lanceolate-fusiform, simple or branched, borne on a small pedicel on terminal branches.
Habitat: Cystoseira compressa was found from 0.5 to 17.8 m deep with abundant patches or almost continuous stands in eight sites along all sides of the island of Marettimo.
Remarks: Cystoseira compressa has shown a stable trend over time, having first been reported by Giaccone (1970) and subsequently also by Giaccone and Sortino (1974) and Catra et al. (2006).
Cystoseira foeniculacea (Linnaeus) Greville f. tenuiramosa (Ercegović) A. Gómez Garreta, M.C.Barceló, M.A.Ribera et J. Rull Lluch (Figure S1).
Morphological description of specimens: thalli are caespitose, with several rough and cylindrical axes originating from an irregular discoid holdfast (Figure S1A). The apex is very prominent and covered by spines (Figure S1B). Primary branches are cylindrical, entirely covered by small spines (Figure S1C), knotty, and bear cylindrical higher order branches. The last-order branches are filiform. Receptacles of this form were not observed during this study.
Habitat: Cystoseira foeniculacea f. tenuiramosa was found in the western side of the island (encompassing Zones A and B) and in the southern one (Zone B) from 0.5 to 26.9 m deep with scattered individuals in deeper waters and abundant patches in shallow environments, as well as in infralittoral pools.
Remarks: This form has shown a stable trend over time, as was previously found by Giaccone and Sortino (1974) and Catra et al. (2006).
Cystoseira cf micheleae Verlaque, Blanfuné, Boudouresque, Thibaut et Sellam (Figure S2).
Morphological description of specimens: thalli are not caespitose (Figure S2A), showing an axis originating from a robust discoid holdfast. The apex is not prominent and difficult to detect since it is usually surrounded by tophules. The axis single or branched is often entirely covered by spinose branches, which can trap sediment (Figure S2B) or can be enveloped by other organisms like sponges (Figure S2C). Primary branches are cylindrical at the base of the branching and then become flattened with a midrib. They can be branched in one or several planes and carry a simple or bifid spinose appendage. Tophules are ovoid and spinose in young thalli and rough and tuberculate in older thalli. Receptacles of this species were not observed during this study.
Habitat: Cystoseira micheleae was found along the southern and north-western coasts of the island (both falling within Zone B) from 12.8 m to 34.6 m with scattered individuals, sometimes forming small patches.
Remarks: Cystoseira micheleae is reported for the first time for the E-MPA. To date, this species has only been reported along the coasts of Tunisia (Montagne 1838), Algeria (Sellam et al. 2017), and recently in the Aeolian archipelago (Marletta et al. 2023). This species is morphologically similar to C. montagnei J. Agardh, from which it differs in the tophules, which are spinose and become rugose with age, and the receptacles, which can be basal, intercalary or terminal (Sellam et al. 2017). During this study, we did not find any fertile specimens, but we believe that they belong to C. micheleae because of the tophules and the general habit of the specimens, which match to those described by Sellam et al. (2017).
Cystoseira pustulata (Ercegović) Neiva et Serrão (Figure 3).

Cystoseira pustulata. (A) Habit; (B) detail of frond with receptacles (white arrows).
Morphological description of specimens: thalli are delicate and tangled (Figure 3A), caespitose, attached to the substratum by a small discoid holdfast. The axes are cylindrical, without spinose appendages. The apex is smooth and not prominent. Primary branches are cylindrical at the base and become slightly compressed in the distal portion. Higher order branches are cylindrical. Receptacles are simple, tiny, fusiform, spindle-shaped (Figure 3B).
Habitat: Cystoseira pustulata was found in 12 of the 18 examined sites, covering almost all sides of the island, from 0.5 to 21 m deep, with scattered individuals or forming small patches.
Remarks: Catra et al. (2006), and previously Giaccone and Sortino (1974), reported the presence of Cystoseira humilis in the E-MPA. Through molecular analyses, Neiva et al. (2022) found that C. humilis is distributed in the Atlantic Ocean and most of the previous records of this species are misidentifications that may refer to C. pustulata.
Ericaria amentacea (C. Agardh) Molinari et Guiry (Figure S3).
Morphological description of specimens: thalli are caespitose (Figure S3A, B) with an intricate and encrusting base, which fits deeply into the substratum and strongly adheres to it (Figure S3C). From the base several cylindrical axes arise. Primary branches are cylindrical and with a pyramidal shape. Very often the lower portion of the longer branches has a wavy appearance as a result of the fall of the higher order branches. All branches bear spinose appendages. Receptacles are compact, cylindrical, and with deciduous spines.
Habitat: Ericaria amentacea was found along all the sides of the island, in shallow (0–0.8 m) and very exposed waters, forming continuous belts.
Remarks: Ericaria amentacea has shown a stable trend over the years in the E-MPA. Bermejo et al. (2018) and Neiva et al. (2022) highlighted that E. amentacea together with Ericaria mediterranea and Ericaria selaginoides (Linnaeus) Molinari et Guiry form a single complex, named E. selaginoides complex. These three species do not have distinct boundaries and natural hybridization among them can occur. This implies a low genetic diversity among these entities, and they may correspond to a single taxon with great morphological plasticity (Bermejo et al. 2018; Neiva et al. 2022).
Ericaria barbatula (Kützing) Molinari et Guiry (Figure S4).
Morphological description of specimens: thalli are caespitose (Figure S4A) with several axes originating from an irregular discoid holdfast. Axes are knotty (Figure S4B), bare in the distal part, and bear branches only in the distal portion. The apex is smooth and very prominent in relation to the insertion of branches (Figure S4C). Primary and higher order branches are cylindrical. Receptacles are compact and wrinkly-verrucose, provided with deciduous spines (Figure S3D).
Habitat: Ericaria barbatula was found along the western coasts of the island (covering Zones A and B) in shallow environments (0.5–1 m) with continuous stands.
Remarks: Ericaria barbatula was not previously reporteEricaria in the E-MPA. It may have been overlooked until now since it shares the same ecological niche of Ericaria crinita, or most likely it could be a recent arrival from nearby areas. From a genetic standpoint, according to Neiva et al. (2022), all the samples identified as E. crinita or E. barbatula are part of a single complex defined E. crinita complex. Nevertheless, these species are different at the apex (smooth in E. barbatula, spinose in E. crinita) and in the habit of the thalli (robust in E. barbatula and slender in E. crinita).
Ericaria brachycarpa (J. Agardh) Molinari et Guiry (Figure 4A–C).

Ericaria brachycarpa v. brachycarpa. (A) Habit; (B) detail of smooth apex slightly prominent (white arrow); (C) detail of smooth apex surrounded by spinose appendages (white arrow). Ericaria brachycarpa var. claudiae. (D) Detail of apex (white arrow) and pseudotophules (white arrowheads).
Morphological description of specimens: thalli are characterised by a greenish-brown colouration (Figure 4A) and are caespitose with several axes originating from a wide discoid base. Axes knotty and rugose. Apices are smooth and slightly prominent (Figure 4B), sometimes surrounded by spinose appendages and the apical portion appears to be spinose (Figure 4C). Primary branches are cylindrical and may have small spines near the base. Higher order branches are cylindrical without spines. Receptacles were not observed during this study.
Habitat: Ericaria brachycarpa was found along all the sides of the island from 0.5 to 5 m deep, forming continuous stands.
Remarks: Ericaria brachycarpa has maintained a stable trend on the island, as previously reported in the E-MPA by Giaccone and Sortino (1974) and Catra et al. (2006).
Ericaria brachycarpa var. claudiae Boudouresque, Perret-Boudouresque et Blanfuné (Figure 4D).
Morphological description of specimens: thalli have a greenish colouration and are caespitose and attached to the substratum by either a wide discoid holdfast or haptera. Apices are smooth and slightly prominent (Figure 4D). The axes of this variety are characterised by basal swellings, which are considered as pseudo-tophules (Figure 4D). Primary and higher order branches are cylindrical with a few spinose appendages. Receptacles are compact, rough, and provided with deciduous spines.
Habitat: Ericaria brachycarpa var. claudiae was found along all sides of the island from 9 to 27.8 m deep with scattered individuals or abundant patches.
Remarks: Ericaria brachycarpa var. claudiae was not previously reported in the E-MPA. Furnari and Serio (2022) recently validated the name E. brachycarpa var. claudiae.
Ericaria crinita (Duby) Molinari et Guiry (Figure S5).
Morphological description of specimens: thalli are caespitose (Figure S5A), with several axes originating from a discoid holdfast. Axes are rough, slender and almost bare in the basal portion. The apices are covered by small spines and are prominent (Figure S5B). Primary and higher order branches are cylindrical (Figure S5A) slender and with small spines. Receptacles are cylindrical, compact, rough and without spines (Figure S5C).
Habitat: Ericaria crinita was found along the western coasts of the island (encompassing Zones A and B), forming continuous stands with E. barbatula in shallow waters (0.5–1 m).
Remarks: After the reports by Giaccone (1970) and Giaccone and Sortino (1974), E. crinita may have suffered a regression, since it was not found by Catra et al. (2006). The finding of the species after almost 20 years could suggest that it maintained in a few areas and only recently found better conditions to thrive.
Ericaria dubia (Valiante) Neiva et Serrão (Figure S56).
Morphological description of specimens: thalli with cylindrical stoloniferous primary cauloid, from which erect primary branches arise, cylindrical at the base then ribbon-like in the upper third (Figure S6A). The ribbon-like portions are subdichotomous with entire margins and an evident median midrib (Figure S6B). Receptacles were not observed.
Habitat: Ericaria dubia was found on almost all sides of the island, from 20 to 42.2 m deep with scattered individuals or forming small patches.
Remarks: Ericaria dubia was reported by Giaccone and Sortino (1974) only from the island of Favignana.
Ericaria funkii (Schiffner ex Gerloff et Nizamuddin) Molinari et Guiry (Figure 5).

Ericaria funkii. (A) Habit; (B) detail of apex (arrowhead); (C) detail of a spinose tophule (white arrow).
Morphological description of specimens: thalli have a strong iridescence in situ (Figure 5A), are not caespitose, with a single axis that divides to create a coralloid appearance, and are attached to the substratum by a wide digitiform holdfast. The apex is smooth and not prominent (Figure 5B). Primary and higher order branches are cylindrical and covered by spinose appendages (Figure 5A). Tophules are oblong, rough and spinose (Figure 5C). They are arranged in a corymb in the apical zone of the axis. Receptacles are cylindrical, diffuse and with spinose appendages.
Habitat: Ericaria funkii was observed along all sides of the island, except the east, from 14.6 to 42 m deep forming abundant patches.
Remarks: Ericaria funkii has shown a stable trend on the island, as previously reported in the E-MPA by Giaccone and Sortino (1974) and Catra et al. (2006).
Ericaria zosteroides (C. Agardh) Molinari et Guiry (Figure S7).
Morphological description of specimens: thalli are not caespitose, with a short single axis, which branches out in height or extension (Figure S7A). The holdfast is formed by simple or branched haptera. Apex is smooth and not prominent. Primary branches are cylindrical near the base and flattened in the distal portion. Higher order branches are ribbon-shaped, with an alternate–distichous branching and a faint midrib, and are provided with flattened spines (Figure S7B). Tophules are ovoid and smooth and located on the basal part of the thallus (Figure S7C). Receptacles were not observed.
Habitat: Ericaria zosteroides was found along the southern and north-western coasts of the island (corresponding to Zone B) from 20 to 42 m forming abundant patches.
Remarks: This species has shown a stable trend over time in the E-MPA, as was recorded in all previous records (Catra et al. 2006; Giaccone 1970; Giaccone and Sortino 1974).
Gongolaria montagnei var. compressa (Ercegović) Verlaque, Blanfuné, Boudouresque et Thibaut (Figure S8).
Morphological description of specimens: thalli are not caespitose attached to the substratum by a discoid holdfast (Figure S8A). The axis is short and can be simple or ramified. Apex is smooth, and not prominent (Figure S8B). Primary branches are cylindrical near the base and flattened in the distal portion. Higher order branches are flattened with an alternate arrangement and a central midrib. They can often carry spinose appendages. Tophules are located near the base of the axis and are oblong and spinose. Receptacles were not observed.
Habitat: Gongolaria montagnei var. compressa was found along the eastern side of the island (in Zone C) and along the southern side (in Zone B), from 23.3 to 35.7 m deep generally with scattered individuals or more rarely forming abundant patches as at Secca del Cammello (SC).
Remarks: this species was not previously reported in the E-MPA.
Gongolaria sauvageauana (Hamel) Molinari et Guiry (Figure S9).
Morphological description of specimens: thalli are not caespitose, (Figure S9A), attached to the substratum by a discoid holdfast. Axis can be simple or branched and is spinose and very prominent in relation to the insertion of the primary branches (Figure S9B). Primary branches are cylindrical and slender. Higher order branches are cylindrical and carry delicate and divaricate spinose appendages (Figure S9B). Receptacles are cylindrical, diffuse, with a few spinose appendages.
Habitat: Gongolaria sauvageauana was found along all sides of the island, except the eastern coast, from 6.9 to 39.4 m deep, with scattered individuals, except in the site of Punta Cretazzo (CRE) where it formed abundant patches.
Remarks: this species was previously reported by Giaccone and Sortino (1974), but it was not subsequently found by Catra et al. (2006). It is likely that this species has recently found better conditions to thrive and recover after a period of regression.
Sargassum acinarium (Linnaeus) Setchell (Figure 6A).

Sargassum spp. (A) Sargassum acinarium, habit; (B–C) Sargassum cf furcatum, habit (B) and detail of foliose dichotomous branches (C); (D–E) Sargassum vulgare, habit (D) and detail of frond (E) with receptacles (white arrowheads).
Morphological description of specimens: thalli are not caespitose with a single axis attached to the substratum by a discoid holdfast. Axis is cylindrical, rough and knotty. Primary branches are cylindrical and have narrow foliaceous branches (Figure 6A). These have a central midrib, denticulate margin and acute apex. Aerocysts are located on the upper portion of the thallus (Figure 6A). They are spherical and are sited on a small cylindrical pedicel. Receptacles were cylindrical, slender and simple or bifurcated and borne at the base of a pedicel at the axil of the foliaceous branches.
Habitat: Sargassum acinarium was found on the north-eastern side (in Zone C) at 30.6 m forming abundant patches, and on the south-western side (Zone B) at 15.4 m deep with scattered individuals.
Remarks: Sargassum acinarium was previously reported by Giaccone and Sortino (1974) and was no longer observed. The finding of the species in only two sites on the island of Marettimo might suggest that this species has suffered a severe regression over the years and has been confined to only a few areas. Therefore, this species should be given more attention in the future.
Sargassum cf furcatum Kützing (Figure 6B and C).
Morphological description of specimens: thalli with a creeping habit not caespitose, attached to the substratum by a small discoid holdfast. Axis is knotty and short and divides into open and spaced branches, usually arranged in a fan shape (Figure 6B). Apex is smooth and slightly prominent. Primary and secondary branches are knotty and have foliose lanceolate or lobate branches with a central midrib, wavy margins, and an acute apex (Figure 6B). The foliose branches are dichotomously divided from one to four times and the midrib follows these bifurcations (Figure 6C). Aerocysts and receptacles were not observed during this study.
Habitat: Sargassum cf furcatum was found along the southern (Zone B) and north-eastern (Zone C) coasts, with scattered individuals, from 13.1 to 21.3 m deep.
Remarks: This species has never been reported before from the island. The identification at the species level was not certain because we did not find fertile specimens during this study. However, along the Ionian coast of Sicily, this species is usually fertile from June to August (Marletta et al. 2024). This could depend on the fact that the seawater temperature in June in the sites where the phenology of this species was studied is around 22–26 °C (Marletta et al. 2024), while in Marettimo it was around 17–22 °C in the same period. This is most likely due to the upwelling currents affecting the south and western parts of Sicily (Béranger et al. 2004). The finding of this species in Marettimo could further prove the expanding trend of S. furcatum in the Central Mediterranean.
Sargassum vulgare C. Agardh nom. illeg. (Figure 6D and E).
Morphological description of specimens: thalli are not caespitose, attached to the substratum by a discoid holdfast. The axis is knotty and can be simple or branched. Apex is smooth and not prominent. Primary branches are cylindrical and knotty and have secondary branches with alternate-distichous branching. Foliose branches are lanceolate, with an evident midrib, toothed or with wavy margins and acute apex (Figure 6D). Areocysts are spherical, with a short cylindrical pedicel, and are located at the axil of the foliose branches (Figure 6D). Receptacles are cylindrical or slightly flattened, warty, simple, or branched, and are borne on a short sterile pedicel (Figure 6E).
Habitat: Sargassum vulgare was found along all sides of the island (except the eastern coast, Zone C) from 0.5 to 25.5 m deep, with scattered individuals or forming dense patches.
Remarks: This species has shown a stable trend on the island over the years, as it was reported by Giaccone (1970), Giaccone and Sortino (1974), and Catra et al. (2006).
4 Discussion and conclusion
In this study, we explored the diversity of fucalean species on the island of Marettimo, about 20 years after the last floristic study conducted in the E-MPA. We found a total of 17 Fucales: four belonging to Cystoseira, eight to Ericaria, two to Gongolaria, and three to Sargassum. By the comparison with three previous studies (Catra et al. 2006; Giaccone 1970; Giaccone and Sortino 1974) emerged some differences, for instance, the disappearance of nine species between the studies carried out in the 1970s and that of Catra et al. (2006): Cystoseira foeniculacea f. latiramosa, Ericaria crinita, E. dubia, E. mediterranea, Gongolaria barbata, G. elegans, G. sauvageauana, Sargassum acinarium and S. hornschuchii. However, according to Catra et al. (2006), even if there was a regression of these species, the communities they formed were still well structured, unlike other islands (such as Pantelleria or Linosa), where a severe decline has already been reported (Alongi et al. 2004; Serio et al. 2006). In the present study, four of the species listed above were found more than 20 years after their disappearance: E. crinita, E. dubia, G. sauvageauana and S. acinarium. As suggested by Gianni et al. (2013), fragmented populations of fucalean species can persist over time in refuge areas, albeit at low densities. The transition to better management of the E-MPA could likely have allowed these species to thrive in more recent times. This phenomenon has been observed in other Mediterranean areas, such as the Medes Islands (Spain), the island of Ustica (Italy) (Hereu and Quintana 2012; Sala et al. 2012), and more recently, the island of Pantelleria (Italy) (Marletta and Lombardo 2023), where about 20 years after their loss there has been a natural recovery of these species.
During this study we did not find two species previously reported by Catra et al. (2006): G. montagnei and G. montagnei var. tenuior. Nevertheless, five species found during this study were not previously reported from either the island of Marettimo or the E-MPA as a whole: C. cf micheleae, E. barbatula, E. brachycarpa var. claudiae, G. montagnei var. compressa and S. cf furcatum. Furthermore, we observed that the areas with the highest number of species were along the southern side of the island, in Zone B. On the contrary, the eastern areas (encompassing Zone C) showed the lowest number of species, while the sites located along the northern (Zone B) and western (Zone A) coasts presented an intermediate number of species compared to the southern and eastern shores. These results indicate that the enlargement of Zone A towards the southern areas (Zone B) should be considered for better conservation of the species and their habitat.
If we compare the fucalean diversity of the present study with that found by Marletta and Lombardo (2023) at Pantelleria and by Marletta et al. (2023) at Lipari and Vulcano, it is possible to observe that the highest biodiversity occurs in the Sicily Channel. Indeed, at Pantelleria 19 species were found (Marletta and Lombardo 2023), compared with 17 at Marettimo and 13 at Lipari and Vulcano (Marletta et al. 2023). The Sicily Channel is indeed an area of mixing water from the Atlantic Sea and the eastern Mediterranean basin (Civitarese et al. 2010). This enhances the dispersion of species and their propagules from one side of the Mediterranean to the other (Ben Rais Lasram et al. 2008). Moreover, the similarity at species level found in these areas is noteworthy: Pantelleria and Marettimo share nine species, while Marettimo, Lipari, and Vulcano share 10. This may be connected to the water circulation of the Sicily Channel, which is influenced by the Algerian Current. This current brings the upper layer of the Atlantic Water (AW) eastwards and splits into two branches: one flowing to the Tyrrhenian Sea, and the other into the Sicily Channel. In the lower layer at the exit of the Sicily Channel, the Eastern Mediterranean Outflow Water (EOW), mainly composed of Levantine Intermediate Water (LIW), flows into the Tyrrhenian Sea and veers to the right, flowing along the northern coast of Sicily before reaching the Sardinia Channel (Béranger et al. 2004). This complex circulation scheme might explain the pattern of fucalean diversity present in these islands. Therefore, it would be crucial to continue monitoring these areas and those nearby in the near future to gain a complete overview of the distribution patterns and biogeography of Fucales. This could ultimately favour the conservation and, hopefully, the restoration of these species and their associated biodiversity.
Funding source: University of Catania, “PIACERI, PIAno di inCEntivi per la Ricerca di Ateneo 2024-2026, Linea di Intervento 1 “Progetti di ricerca collaborativa”.
Award Identifier / Grant number: unnumbered
About the authors

Giuliana Marletta has a PhD in Earth and Environmental Sciences at the University of Catania. Her research expertise is the field of marine ecology focusing particularly on conservation and ecological restoration of Mediterranean underwater forests. Furthermore, her research covers studies on the dynamics of benthic vegetation.

Andrea Lombardo is a PhD student at the Department of Biological, Geological and Environmental Sciences, University of Catania. His research concerns the study of Sicilian marine heterobranch (sea slugs) faunas. He is also interested and actively collaborates in studies on brown algae belonging to the order Fucales.

Donatella Serio is a researcher at the University of Catania, Italy. Her current research interests include marine benthic flora and vegetation. The floristic and vegetational searches deal with areas previously studied to estimate changes of biodiversity at both short and long term. In parallel with the above-mentioned research, she is carrying out studies aiming to clarify the taxonomy of some genera of marine macroalgae.
Acknowledgments
The authors wish to thank Prof. Giovanni Furnari for his constructive comments and suggestions. Moreover, they are very grateful to the staff of Marettimo Diving Center (Fabio, Gaia and Maria) and the lovely people met during this research.
-
Research ethics: The procedures are in accordance with the national laws.
-
Informed consent: Not applicable.
-
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and approved its submission. G.M. and A.L. conceived and designed the research; G.M and D.S. analysed the data; G.M. and A.L. wrote the early version of the manuscript; D.S. supervised the research, edited and finalized the manuscript.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors declare that they have no conflicts of interest.
-
Research funding: This work was supported by the University of Catania under the grant “PIACERI, PIAno di inCEntivi per la Ricerca di Ateneo 2024–2026, Linea di Intervento 1 “Progetti di ricerca collaborativa”.
-
Data availability: Not applicable.
References
Alongi, G., Catra, M., Cormaci, M., Furnari, G., and Serio, D. (2004). Spring marine vegetation on rocky substrata of Pantelleria Island (the straits of Sicily, Italy). Nova Hedwig 79: 447–478, https://doi.org/10.1127/0029-5035/2004/0079-0447.Search in Google Scholar
Ben Rais Lasram, F., Tomasini, J.A., Romdhane, M.S., Do Chi, T., and Mouillot, D. (2008). Historical colonization of the Mediterranean Sea by Atlantic fishes: do biological traits matter? Hydrobiologia 607: 51–62, https://doi.org/10.1007/s10750-008-9366-4.Search in Google Scholar
Béranger, K., Mortier, L., Gasparini, G.P., Gervasio, L., Astraldi, M., and Crépon, M. (2004). The dynamics of the Sicily Strait: a comprehensive study from observations and models. Deep-Sea Res. II: Top. Stud. Oceanogr. 51: 411–440, https://doi.org/10.1016/j.dsr2.2003.08.004.Search in Google Scholar
Bermejo, R., Chefaoui, R.M., Engelen, A.H., Buonomo, R., Neiva, J., Ferreira-Costa, J., Pearson, G.A., Marbà, N., Duarte, C.M., Airoldi, L., et al.. (2018). Marine forests of the Mediterranean-Atlantic Cystoseira tamariscifolia complex show a southern Iberian genetic hotspot and no reproductive isolation in parapatry. Sci. Rep. 8: 10427, https://doi.org/10.1038/s41598-018-28811-1.Search in Google Scholar PubMed PubMed Central
Blanfuné, A., Verlaque, M., Boudouresque, C.F., Rozis, E., and Thibaut, T. (2022). Les forêts marines de France et de Méditerranée. Guide de détermination des espèces-ingénieurs. Sargassaceae, Fucales, Phaeophyceae. Presses Universitaries de Provence, Marseille, France.Search in Google Scholar
Catra, M., Alongi, G., Serio, D., Cormaci, M., and Furnari, G. (2006). The benthic algal flora on rocky substrata of the Egadi islands, a marine protected archipelago off the western coast of Sicily (Italy, Mediterranean Sea). Nova Hedwig 82: 489–538, https://doi.org/10.1127/0029-5035/2006/0082-0489.Search in Google Scholar
Civitarese, G., Gacic, M., Lipizer, M., and Eusebi Borzelli, G.L. (2010). On the impact of the bimodal oscillating system (BiOS) on the biogeochemistry and biology of the Adriatic and Ionian seas (eastern Mediterranean). B.G. 7: 3987–3997.10.5194/bg-7-3987-2010Search in Google Scholar
Cormaci, M., Furnari, G., Catra, M., Alongi, G., and Giaccone, G. (2012). Flora marina bentonica del Mediterraneo: Phaeophyceae. Boll. Accad. Gioenia Sci. Nat. Catania 45: 1–508.Search in Google Scholar
D’Anna, G., Fernández, T.V., Pipitone, C., Garofalo, G., and Badalamenti, F. (2016). Governance analysis in the Egadi islands marine protected area: a Mediterranean case study. Mar. Policy 71: 301–309, https://doi.org/10.1016/j.marpol.2015.12.009.Search in Google Scholar
Furnari, G. and Cormaci, M. (2009). Floristic changes in the Mediterranean macroalgal flora. Bocconea 23: 85–92.Search in Google Scholar
Furnari, G. and Serio, D. (2022). Validation of Ericaria brachycarpa var. claudiae Boudouresque, Perret-Boudouresque & Blanfuné (Sargassaceae, Phaeophyceae). Not. Algarum 257: 1–4.Search in Google Scholar
Giaccone, G. (1970). Raccolte di fitobenthos sulla banchina continentale Italiana. Plant Biosyst. 103: 485514, https://doi.org/10.1080/11263506909430507.Search in Google Scholar
Giaccone, G. and Sortino, M. (1974). Zonazione della vegetazione marina delle Isole Egadi (Canale di Sicilia). Lav. Ist. Bot. Giard. Colon. Palermo 25: 166–183.Search in Google Scholar
Giaccone, G., Alongi, G., Cossu, A.V.L., Di Geronimo, R., and Serio, D. (1993). La Vegetazione marina bentonica nel Mediterraneo: 1: sopralitorale e mesolitorale: proposte di aggiornamento. Boll. Accad. Gioenia Sci. Nat. 26: 245–291.Search in Google Scholar
Gianni, F., Bartolini, F., Airoldi, L., Ballesteros, E., Francour, P., Guidetti, P., Meinesz, A., Tibaut, T., and Mangialajo, L. (2013). Conservation and restoration of marine forests in the Mediterranean Sea and the potential role of Marine Protected Areas. A.I.O.L. 4: 83–101, https://doi.org/10.1080/19475721.2013.845604.Search in Google Scholar
Gómez-Garreta, A., Barceló-Martí, M.C., Ribera-Siguan, M.A., and Rull-Lluch, J. (2001) Flora phycologica iberica. In: Gómez-Garreta, A. (Ed.), Cystoseira C. Agardh. Universidade de Murcia. Múrcia, Spain, pp. 99–166.Search in Google Scholar
Guidetti, P., Milazzo, M., Bussotti, S., Molinari, A., Murenu, M., Pais, A., Spanò, N., Balzano, R., Agardy, T., Boero, F., et al.. (2008). Italian Marine reserve effectiveness: does enforcement matter? Biol. Conserv. 141: 699–709, https://doi.org/10.1016/j.biocon.2007.12.013.Search in Google Scholar
Guiry, M.D. and Guiry, G.M. (2024). AlgaeBase. World-wide electronic publication. University of Galway, Available at: https://www.algaebase.org (accessed 2 December 2024).Search in Google Scholar
Halpern, B.S. (2003). Impact of marine reserves: do re-serves work and does reserve size matter? Ecol. Appl. 13: 117–137, https://doi.org/10.1890/1051-0761(2003)013[0117:tiomrd]2.0.co;2.10.1890/1051-0761(2003)013[0117:TIOMRD]2.0.CO;2Search in Google Scholar
Harmelin-Vivien, M.L., Harmelin, J.G., Chauvet, C., Duval, C., Galzin, R., Lejeune, P., Barnabé, G., Blanc, F., Chevalier, R., Duclerc, J., et al.. (1985). The underwater observation of fish communities and fish populations: methods and problems. Rev. Ecol. Terre. Vie. 40: 467–540.10.3406/revec.1985.5297Search in Google Scholar
Hereu, B. and Quintana, X. (2012). El fons marí de les Illes Medes i el Montgrí, quatre dècades de recerca per a la conservació. Càtedra d’Ecosistemes Litorals Mediterranis, Universitat de Barcelona, Spain.Search in Google Scholar
La Mesa, G., Salvati, E., Agnesi, S., and Tunesi, L. (2017). Assessment of coastal fish assemblages before the establishment of a new marine protected area in central Mediterranean: its role in formulating zoning proposal. Mediterr. Mar. Sci. 18: 11–21, https://doi.org/10.12681/mms.1788.Search in Google Scholar
Mancuso, F.P., Strain, E.M.A., Piccioni, E., De Clerck, O., Sarà, G., and Airoldi, L. (2018). Status of vulnerable Cystoseira populations along the Italian infralittoral fringe, and relationships with environmental and anthropogenic variables. Mar. Pollut. Bull. 129: 762–771, https://doi.org/10.1016/j.marpolbul.2017.10.068.Search in Google Scholar
Mannino, A.M. and Balistreri, P. (2017). An updated overview of invasive Caulerpa taxa in Sicily and circum-Sicilian Islands, strategic zones within the NW Mediterranean Sea. Flora Mediterr. 27: 221–240.10.7320/FlMedit27.221Search in Google Scholar
Mannino, A.M. and Balistreri, P. (2018). Citizen science: a successful tool for monitoring invasive alien species (IAS) in Marine Protected Areas. The case study of the Egadi Islands MPA (Tyrrhenian Sea, Italy). Biodiversity 19: 42–48.10.1080/14888386.2018.1468280Search in Google Scholar
Mannino, A.M., Donati, S., and Balistreri, P. (2016). The project “Caulerpa cylindracea in the Egadi Islands”: citizens and scientists working together to monitor marine alien species. Biodivers. J. 7: 907–912.Search in Google Scholar
Mannino, A.M., Parasporo, M., Crocetta, F., and Balistrieri, P. (2017). An updated overview of the marine alien and cryptogenic species from the Egadi islands marine protected area (Italy). Mar. Biodiv. 47: 469–480, https://doi.org/10.1007/s12526-016-0496-z.Search in Google Scholar
Marletta, G. and Lombardo, A. (2020). Assessment of grazing impact on deep canopy-forming species in the western Ionian Sea, Central Mediterranean. Int. J. Aquat. Biol. 8: 365–376.Search in Google Scholar
Marletta, G. and Lombardo, A. (2023). The Fucales (Ochrophyta, Phaeophyceae) of the island of Pantelleria (Sicily Channel, Mediterranean Sea): a new contribution. Ital. Botanist. 15: 137–163, https://doi.org/10.3897/italianbotanist.15.103217.Search in Google Scholar
Marletta, G., Lombardo, A., Serio, D., and Bianchelli, S. (2023). Diversity of Fucales (Ochrophyta, Phaeophyceae) along the Coasts of Lipari and Vulcano (Aeolian Archipelago), Tyrrhenian Sea (Central Mediterranean Sea). J. Mar. Sci. Eng. 11: 2222, https://doi.org/10.3390/jmse11122222.Search in Google Scholar
Marletta, G., Lombardo, A., and Serio, D. (2024). Ecology and phenology of the subtidal brown alga Sargassum furcatum (Ochrophyta, Fucales), a likely non-indigenous species from the Mediterranean Sea. J. Mar. Sci. Eng. 12: 640, https://doi.org/10.3390/jmse12040640.Search in Google Scholar
Montagne, C. (1838). Cryptogames algériennes, ou plantes cellulaires recueillies par M. Roussel aux environs d’Alger, et publiées par le Docteur Camille Montagne. Ann. Sci. Nat. Bot. 2: 268–279.Search in Google Scholar
Neiva, J., Bermejo, R., Medrano, A., Capdevila, P., Milla Figueras, D., Afonso, P., Ballesteros, E., Sabour, B., Serio, D., Nóbrega, E., et al.. (2022). DNA barcoding reveals cryptic diversity, taxonomic conflicts and novel biogeographical insights in Cystoseira s.l. (Phaeophyceae). Eur. J. Phycol. 58: 351–375, https://doi.org/10.1080/09670262.2022.2126894.Search in Google Scholar
OECD (2017). Marine Protected Areas: economics, management and effective policy mixes. OECD Publishing, Paris.10.1787/9789264276208-enSearch in Google Scholar
Rasheed, A.R. (2020). Marine protected areas and human well-being: a systematic review and recommendations. Ecosyst. Serv. 41: 101048, https://doi.org/10.1016/j.ecoser.2019.101048.Search in Google Scholar
Rodríguez-Prieto, C., Ballesteros, E., Boisset, F., and Afonso Carrilo, J. (2013). Guía de las macroalgas y fanerógamas marinas del Mediterráneo Occidental. Ediciones Omega, Barcelona, Spain.Search in Google Scholar
Sala, E., Ballesteros, E., Dendrinos, P., Di Franco, A., Ferretti, F., Foley, D., Fraschetti, S., Friedlander, A., Garrabou, J., Guclusoy, H., et al.. (2012). The structure of Mediterranean rocky reef ecosystems across environmental and human gradients, and conservation implications. PLoS ONE 7: e32742, https://doi.org/10.1371/journal.pone.0032742.Search in Google Scholar PubMed PubMed Central
Sales, M., Cebrian, E., Tomas, F., and Ballesteros, E. (2011). Pollution impacts and recovery potential in three species of the genus Cystoseira (Fucales, Heterokontophyta). Estuar. Coast. Shelf Sci. 92: 347–357, https://doi.org/10.1016/j.ecss.2011.01.008.Search in Google Scholar
Sellam, L.N., Blanfuné, A., Boudouresque, C.F., Thibaut, T., Rebzani Zahaf, C., and Verlaque, M. (2017). Cystoseira montagnei J. Agardh and C. spinosa Sauvageau (Phaeophyceae, Sargassaceae): a taxonomic reappraisal of misused names, with the proposal of Cystoseira michaelae Verlaque et al. nom. et stat. nov. Cryptogam. Algol. 38: 133–157.10.7872/crya/v38.iss2.2017.133Search in Google Scholar
Serio, D., Alongi, G., Catra, M., Cormaci, M., and Furnari, G. (2006). Changes in the benthic algal flora of Linosa Island (Straits of Sicily, Mediterranean Sea). Bot. Mar. 49: 135–144, https://doi.org/10.1515/bot.2006.018.Search in Google Scholar
Sortino, M., Calvo, S., and Drago, D. (1976). Sulla corologia, ecologia e fitosociologia di Phyllaria reniformis Rostafinsky nel bacino occidentale del Mediterraneo. Mem. Biol. Mar. Oceanogr. Univ. Messina 6: 125–133.Search in Google Scholar
Thibaut, T., Pinedo, S., Torras, X., and Ballesteros, E. (2005). Long-term decline of the populations of Fucales (Cystoseira spp. and Sargassum spp.) in the Albères coast (France, North-western Mediterranean). Mar. Pollut. Bull. 50: 1472–1489, https://doi.org/10.1016/j.marpolbul.2005.06.014.Search in Google Scholar PubMed
Thibaut, T., Blanfune, A., Boudouresque, C.F., and Verlaque, M. (2015). Decline and local extinction of Fucales in the French Riviera: the harbinger of future extinctions? Mediterr. Mar. Sci. 16: 206–224, https://doi.org/10.12681/mms.1032.Search in Google Scholar
Thibaut, T., Blanfuné, A., Boudouresque, C.F., Cottalorda, J.M., Hereu, B., Susini, M.L., and Verlaque, M. (2016). Unexpected temporal stability of Cystoseira and Sargassum forests in Port-Cros, one of the oldest Mediterranean marine National Parks. Cryptogam. Algol 37: 61–90.10.7872/crya/v37.iss1.2016.61Search in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2024-0105).
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
- Supplementary Material Details
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX