Fucus vesiculosus adapted to a life in the Baltic Sea: impacts on recruitment, growth, re-establishment and restoration
-
Lena Kautsky
Lena Kautsky is Professor Emeritus in Marine Plant Ecology at Stockholm University, Department of Ecology, Environment and Plant Science and senior advisor at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on benthic vegetation and dynamics in
Fucus communities. Furthermore, her research covers studies of biodiversity both at the genetic and species level, as well as effects and impact of pollution on benthic vegetation and issues on integrated aquaculture and restoration of shallow benthic communities ,
Susanne Qvarfordt
,
Susanne Qvarfordt
Susanne Qvarfordt has a PhD in Baltic Sea Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science. Her research is mainly focused on vegetation in the Baltic Sea, both algae and vascular communities, and how these communities vary over time.
Ellen Schagerström has a PhD in Plant Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science and at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on biodiversity both at the genetic and species level in
Fucus communities as well as adaptation and speciation. Furthermore, her research covers studies of dynamics of benthic vegetation as well as restoration of shallow benthic communities.

Abstract
Fucus vesiculosus is common both on the tidal coasts of the North Atlantic and in the Baltic Sea, where it has adapted to low salinity and nontidal conditions over the last 7000 years. During the late 1970s and early 1980s, extensive declines of F. vesiculosus populations were reported in the Baltic Proper, mainly attributed to high nutrient loads. During the past 30–40 years, considerable efforts have been made to reduce nutrient runoff to coastal areas but few successful initiatives to restore F. vesiculosus populations have been performed. In this paper, we present how substratum manipulation, i.e. clean rocky surfaces, brushing rocks, Hildenbrandia rubra cover and different filamentous algae, as well as different algal exudates, affect the recruitment and survival of juvenile F. vesiculosus. Further, we show through a 5-year field experiment that it will take at least 4–5 years to reach reproductive age for F. vesiculosus in the Baltic Sea. We also present transplantation studies from two different areas, showing that epiphytic load, light, grazing and type of substratum are some of the factors that need to be taken into consideration in order to achieve successful restoration of F. vesiculosus.
Introduction
Fucus vesiculosus L., together with Fucus serratus L. and Fucus radicans Bergström et Kautsky are the only large marine perennial macroalgae that have been able to establish and adapt to the low salinity of the Baltic Sea. Fucus vesiculosus, together with F. serratus in the southern part and the endemic F. radicans in the Bothnian Sea, are all considered foundation species in the Baltic Sea ecosystem (Wahl et al. 2011, Schagerström et al. 2014). The marine fucoids entered the Baltic Sea basin around 7000 years ago, and have ever since been adapting to the strong evolutionary pressures this environment presents (Russell 1985, Snoeijs 1999). Both F. vesiculosus and F. serratus have substantially higher tolerance to low salinity compared to North Sea populations (Serrão et al. 1996a, 1999, Malm et al. 2001). Genetic studies have shown a distinct ecotypic difference between populations of F. vesiculosus on the marine (salinity>30) west coast of Sweden compared to the populations in the Baltic Sea (Johannesson and André 2006).
On the Swedish west coast, Fucus vesiculosus grows in the intertidal down to about 0.5 m, its lower distribution limit governed mainly by competition from Fucus serratus, and other large perennial brown algae. In the Baltic Sea, however, F. vesiculosus covers large areas from the surface down to ca. 10 m in more open areas and 2–4 m in nutrient-enriched areas (Kautsky et al. 1986, Torn et al. 2006). The lack of competition from other fucoids and kelp species at greater depths increases the importance of light and substratum availability as the limiting factors for depth distribution (Waern 1952, Snoeijs 1999). Like marine fucoids growing in the intertidal zone, Baltic fucoids are exposed daily to stress factors such as desiccation, freezing or ice scraping linked to seasonal changes in air temperature (Pearson et al. 2000, Wahl et al. 2011) and refs. therein). In the nontidal Baltic Sea, extended periods of meteorologically high and low water levels expose the uppermost part of the F. vesiculosus populations to risk of long periods of freezing and desiccation during low water periods in spring (Malm and Kautsky 2003). Since the Baltic F. vesiculosus has a reduced tolerance to desiccation and freezing (Pearson et al. 2000, Lago-Leston et al. 2010), this is a substantial factor for limiting the upwards expansion. Varying and extended periods of high and low water periods also have a marked impact on shallow growing filamentous annual algae (Lago-Leston et al. 2010) which, by reducing competition for space, may have an indirect impact on the establishment of fucoid germlings.
The main reproductive period varies among Fucus vesiculosus populations on a local scale. On the Swedish west coast and in the Baltic Sea, reproduction and recruitment of new germlings occur in early summer, i.e. May – June (Bäck et al. 1991). However, some populations have been reported to also reproduce during autumn (September–October) in areas of heated water outside a power plant on the Swedish west coast (Carlson 1991) or in rock pools close to the sea (L. Kautsky, personal obs.). In the Baltic Sea, a specific form of F. vesiculosus reproduces during September–October (Berger et al. 2001). A few populations bearing mature receptacles throughout the reproductive season, from May to October, have been reported from Poel in Germany (Maczassek 2014) and have also been found around Gotland (E. Schagerström, personal obs.).
Mortality during the microscopic stages is very high; less than 0.015% reach a visible size as a germling (Chapman 1995). No study to our knowledge has followed a newly established Fucus vesiculosus population until its first reproduction to document the age at the first sexual reproduction in the Baltic Sea. Vegetative regeneration from holdfasts is also common in F. vesiculosus and a way to recover from mechanical damage, mainly ice scouring (Kiirikki and Ruuskanen 1996, Malm and Kautsky 2004) in shallow Baltic populations.
The cover of filamentous algae, both species composition and biomass, differs during the above mentioned periods of F. vesiculosus recruitment (Berger et al. 2001, Råberg et al. 2005). This cover might vary the negative impact on zygotes attaching to the substratum. For example, Pylaiella littoralis (L.) Kjellman negatively affects juvenile survival both by its mere physical presence and by producing exudates (Berger et al. 2003, Råberg et al. 2005), thus reducing the successful establishment of F. vesiculosus as well as other species (Qvarfordt 2006). Filamentous algae may also trap more sediment particles, thus further obstructing successful settlement and the recruitment of a new population (Berger et al. 2003, Eriksson and Johansson 2003). In nutrient enriched areas of the Baltic Sea, the autumn-reproducing populations could have a higher probability for successful establishment of new populations, due to less filamentous algae and sediment on rocky surfaces during this time of year (Berger et al. 2004).
During the 1970s and 1980s, several reports described substantial decreases of Fucus distribution in Estonian (Kukk and Martin 1992), Finnish (Kangas et al. 1982, Haahtela 1984), German (Vogt and Schramm 1991), Lithuanian (Olenin and Klovaité 1998), Polish (Plinski and Florczyk 1984) and Swedish coastal areas (Lindvall 1984, Kautsky et al. 1986, Rosemarin et al. 1986, Nilsson et al. 2004). In some areas of the Archipelago Sea, a continued decline of Fucus vesiculosus has been reported (Vahteri and Vuorinen 2016). Direct and indirect effects of nutrient enrichment (Torn et al. 2006, Alexandridis et al. 2012), grazing by isopods from the genus Idotea (Engkvist et al. 2000), and toxic compounds (e.g. chlorinate from pulp mill effluents; Kautsky 1992, Wikström et al. 2000), have been suggested to be responsible for these declines. At the same time, the amount of filamentous algae in the coastal areas has increased, especially during spring and summer, which could effectively reduce the space available for the settling of Fucus germlings (Berger et al. 2001). Natural recovery has since been recorded locally both in Finland (Kangas and Niemi 1985, Rönnberg et al. 1985) and along the Swedish Baltic Proper coast (Eriksson et al. 1998, Engkvist et al. 2002).
The most pronounced effect of increased nutrient loads is reduced light penetration due to increased turbidity from phytoplankton blooms (Larsson et al. 1985, Sandén and Håkansson 1996). Several long-term studies have shown how Fucus vesiculosus depth distribution and areal cover has been negatively affected by reduced Secchi depth (Kautsky et al. 1986, Bergström et al. 2013). The impact of large-scale nutrient enrichment on F. vesiculosus may be more pronounced in the Baltic Sea than in tidal areas, e.g. the North Sea, due to the deeper occurrence of fucoids. In the Baltic Sea, decreasing Secchi depth caused by phytoplankton blooms has been recorded on local scales in many shallow nutrient enriched areas, e.g. outside sewage treatment plants, but also for the Baltic Sea basins on a larger scale (Andersen et al. 2017). Local disappearance or reduction in cover may also be attributed to changes in top-down regulation through the cascading effect of fish feeding on mesograzers. Reduced abundances of mesograzers will have positive effects on the growth of competing filamentous algae (Eriksson et al. 1998, Worm et al. 1999) and thus have an indirect negative effect on F. vesiculosus germlings (Malm et al. 1999, Korpinen et al. 2007).
Over the past 30–40 years, substantial efforts have been made to reduce the nutrient load to the Baltic Sea. Positive results have been noted in the increased depth distribution of F. vesiculosus in some coastal areas (Eriksson et al. 1998, Nilsson et al. 2004). However, at the same time, F. vesiculosus has not recovered in some other areas and heavy grazing by Idotea spp. has been recorded (Engkvist et al. 2004, Torn et al. 2006). The nutrient load can be high in coastal areas, thus indirectly limiting the growth of F. vesiculosus by favouring fast growing epiphytic filamentous algae, which cover and shade adult Fucus thalli, thereby reducing its growth (Hemmi et al. 2005, Rohde et al. 2008). Also, invertebrates favoured by high loads of particles, e.g. barnacles and bryozoans, settle on the Fucus thalli, both shading the thallus and weighing it down and thereby making it more susceptible to sediment cover (Oswald et al. 1984, Worm and Lotze 2006).
Because of the vital role of Fucus vesiculosus as a foundation species and, possibly, both a direct carbon source and a potential carbon donor to receiver sites (Rönnbäck et al. 2007, Hill et al. 2015), management activities for re-introducing Fucus to areas where it has disappeared are needed. With expected future climate changes and investigations relating to storage of carbon in the sea, a new focus of interest has been on activities restoring and managing Fucus belts and communities (Rönnbäck et al. 2007). So far there are few reports of active re-introduction of Fucus into areas where it has been completely lost (e.g. outside pulp mills) or reduced in abundance, e.g. outside sewage treatment outlets (Kautsky 1992, Wikström et al. 2000).
There is a need to increase our knowledge regarding the impacts on different life stages of Fucus vesiculosus in order to succeed with the management, protection, and restoration of Fucus habitats in the nontidal Baltic Sea. The aim is to increase further our understanding of factors impacting successful recruitment and re-establishment of F. vesiculosus, by showing how settlement, attachment, and survival in F. vesiculosus germlings are affected by some common species of macroalgae in the Baltic Sea.
In this paper, we present a set of field experiments on recruitment, juvenile growth, and survival. We also examine the effects of competing filamentous and crust-forming macroalgal species through physical interference and the possible chemical impact of their exudates on the early stages of F. vesiculosus. Further, we summarise a few early transplantation trials in two different areas, focusing on the growth and survival of adult Fucus.
Materials and methods
Study areas
Field experiments and studies were performed in the northern Baltic Proper, on the Swedish coast in the Trosa archipelago south of Stockholm. The area around Askö island is a marine nature reserve and the area outside the small town of Trosa has a gradient of nutrient-enriched wastewater from the sewage treatment plant (Sites 1–2, Figure 1A, B). A field experiment was also conducted at Räfsnäs (Site 3, Figure 1A) in the northern part of the Stockholm archipelago. In the whole area, Fucus vesiculosus (hereafter Fucus) occurs in the sub-littoral, covering large parts of the rocky surface between 0.5 and 6.0 m depth. The dense Fucus communities, with a high biomass per m2 harbour a large number of associated flora and fauna (Wallentinus 1979, Wikström and Kautsky 2007, Schagerström et al. 2014). The benthic macroalgal cover below Fucus is dominated by many filamentous algae together with the crust forming red alga Hildenbrandia rubra (Sommerfelt) Meneghini (hereafter Hildenbrandia) (Berger et al. 2001). The transplantation trials were performed in Trosa archipelago (Sweden) and Gdansk Bay (Poland) (Site 5, Figure 1). In the Puck Bay area of Gdansk Bay, the formerly rich vegetation of Fucus deteriorated rapidly from 1970 mainly due to high nutrient load and low Secchi depth (Plinski and Florczyk 1984), see refs. in Schramm and Nienhuis 1996) while no large-scale reduction of the cover of Fucus had been recorded in the less nutrient enriched Trosa archipelago (Wallentinus 1979).

Maps showing areas and sites for field experiments presented in this paper.
(A) Map of the Baltic Sea showing areas 1–5. (B) Area 1 with Sites A–G outside Trosa town (black) and Area 2 with control Site H near Askö laboratory on Askö island. (C) Area 5 with Sites I–J in Gdansk Bay.
Survival and growth of a naturally recruited Fucus population
Survival and growth patterns of a naturally recruited population of Fucus germlings were studied at Askö Laboratory (Figure 1B, Site H) from 1991 to 1996. The germlings recruited in the field on stones and small rocks in 1991, growing in a sheltered shallow bay at ca. 0.5 m depth (mean water level, MWL) in an area of 5×5 m. Thalli were collected during summer after 12, 24, 38, 50 and 62 months. The collection was performed randomly to ensure that all sizes occurring were harvested. Increase in length of the thallus, the circumference at the upper, thickest part of the thallus, number of dichotomies, and biomass [g dry weight (DW)] was measured in the laboratory. Thallus volume (cm3) was calculated as a cone, using frond length and circumference. The numbers of thalli harvested in the first 4 years were 50, 60, 40 and 30, respectively. At the last harvest, in 1996, only 10 thalli were measured. These differences were due to an extended low water period that few individuals survived. At the same time, changes in density per m2 were estimated.
Effects of Fucus vegetation cover on Fucus settlement and survival
In 1994 a re-establishment experiment was performed along a moderately sheltered rocky shore close to the Askö Laboratory. Inside a natural Fucus belt, six 1-m wide zones reaching 5 m out into the water from the shoreline were prepared on June 5th. Four different treatments were created in each 1×5-m zone:
Cladophora-covered substratum: at the uppermost part of the zone, the vegetation consisted of almost 100% cover of Cladophora glomerata (Linneus) Kützing (hereafter Cladophora). Any other algae were removed.
Cleaned substratum: the bedrock surface was completely cleared from both Fucus and any understorey vegetation of filamentous algae using brushes and putty knives.
Manipulated Fucus canopy: the bedrock was completely cleared as in the Cleaned substratum treatment, and two Fucus thalli were then attached to an anchored rope, creating a Fucus canopy without any understorey.
Natural (control) treatment: both Fucus and understorey vegetation remained intact. In the Natural treatment, filamentous algae [Cladophora, Pylaiella and Ceramium tenuicorne (Kützing) Waern (hereafter Ceramium)] dominated the understorey vegetation, and Hildenbrandia partly covered the substratum.
Six settlement discs (28 cm2, Sea Going Poxy Putty; Serrão et al. 1996b) were placed in each treatment on June 5th at about 1 m depth (MWL), except those that were placed in the Cladophora-covered substratum at about 0.2 m depth (MWL). Ripe Fucus receptacles were collected from the area, brought to the laboratory and the sex determined. Receptacles were kept cold, dark and moist and egg release was performed according to Andersson et al. (1994). For the release of gametes, female receptacles were kept in large glass trays with natural seawater, and male receptacles were stored without water in small glass beakers, all kept in a temperature controlled room at +15°C. After gamete release, the egg and sperm suspensions were filtered and mixed (Andersson and Kautsky 1996). After fertilisation for 2 h, the zygote suspension was dispersed over the settlement disks in the field via a tube placed below the Fucus canopy or about 30 cm above the disk surface under calm evening conditions (Pearson et al. 1998). Dispersal of zygote suspensions was performed on two occasions, July 24th and 30th. On five dates, July 13th and 27th, August 10th, September 8th and November 2nd, the disks were collected for counting germlings and were then replaced. During the study, the disks were colonised mainly by Cladophora and Ceramium.
Data were analysed with a Repeated Measures ANOVA with the factor Treatment nested under Time of sampling. There were differences (p<0.001) between these factors but, due to the high temporal variation within the system, data were further analysed by one-way ANOVAs for each Time of sampling. All analyses in this paper were made using the car library in R version 3.2.3. (R development Core Team 2014), unless otherwise stated.
Effects of filamentous vegetation cover on Fucus settlement and survival
To test whether cleaning of rocky substrates before Fucus gametes settle can improve the survival of Fucus, a field experiment was performed simultaneously at Räfsnäs in the northern Baltic Proper, salinity 5.5 and at Askö, salinity 6.2. Fertile branches of Fucus were collected in late May 2017 and attached to an anchored net (1×2 m) floating close to the surface. For substratum, similar sized stones 8–12 cm across (n=6 at Räfsnäs, n=10 at Askö) with a cover of either Cladophora, Pylaiella littoralis (hereafter Pylaiella) or Hildenbrandia were collected. Filamentous algae were removed from some of the stones with a stiff brush (brushed treatment) and clean stones collected on land without any biofilm were used as controls. Also, ceramic tiles were used as an artificial substratum. On June 1st stones and tiles were placed at about 1 m depth (MWL) below the floating net that held reproductive branches of both male and female Fucus. The net was removed after 4 weeks to avoid shading of settled juveniles. Stones and tiles were collected after 3 or 5 months (i.e. on September 8th at Räfsnäs and on November 1st at Askö) and brought to the laboratory where the number of germlings was counted on an area of 6×6 cm2 on each stone and expressed per dm2.
Data were log transformed to meet the assumption of variance homogeneity and analysed with a one-way ANOVA and Tukey HSD post-hoc tests.
Effects of exudate from filamentous algae and Hildenbrandia on Fucus germination
Hypothesising that exudates from many common macroalgae negatively affect germination and survival of Fucus eggs, a laboratory experiment was performed in June 1997. Exudates from 4 species of filamentous algae Cladophora, Ulva intestinalis L., (hereafter Ulva), Pylaiella and Ceramium and from the crust-forming Hildenbrandia were prepared by collecting fresh material of the filamentous species and stones covered by Hildenbrandia at the Askö Laboratory, Sweden. These were placed in separate aquaria filled with aerated seawater from the area (salinity 6.5, temperature +15°C±1, 16:8 Light:Dark cycle). Approximately 0.5 g DW filamentous algae l−1 water was used. For the Hildenbrandia treatment, stones covered in Hildenbrandia were placed in an aquarium containing 10 l of water. After 48 h, the water containing exudates was filtered off and used in the germination test.
Ripe Fucus receptacles (>50 thalli) were collected in the Askö area, brought to the laboratory and stored moist and dark after determining and separating the sexes. On the day of the experiment, receptacles were covered by seawater. Eggs and sperm were released in separate containers, mixed, and the fertilised eggs were filtered through a 100-μm nylon net and rinsed in seawater. The zygote suspension was concentrated by sedimentation and decantation. The suspension was then added to the water containing exudate from the different algal treatments and the control. After about 1–2 h, 5 ml of the suspension of fertilised eggs in each treatment was transferred to acid-washed Petri dishes, pre-filled with the corresponding treatment water (n=6 dishes per treatment). The number of zygotes in each Petri dish was counted 24 h after fertilisation. Germination and percentage of surviving germlings were calculated after 7 days. Data were analysed with a one-way ANOVA and Tukey HSD post-hoc tests.
Transplantation trials
Eight sites (A–H) were selected along a nutrient and salinity gradient from the open outer archipelago to the small inner bays close to Trosa (Figure 1B). Site A was located at the Trosa river mouth, and was thus affected by freshwater and nutrient runoff. Site B was the point of discharge of the local sewage treatment plant. Sites C, D, and E (site E with a natural stand of Fucus) were placed in a nutrient gradient from site A. Sites F and G, both with natural stands of Fucus, were situated in a shallow bay north of the Trosa estuary. The bay lacks any major inflow of freshwater but is affected by terrestrial nutrient runoff from surrounding agricultural activities. The control site H was located near the Askö Laboratory.
From Site H, 40 Fucus thalli attached to small rocks were collected on May 23rd, 2000 and marked with coloured plastic bands around the stipe. Punching two holes just below the floating bladders marked all apical tips. The thalli were kept in running seawater during the marking process, after which five thalli were transplanted to each site and placed at 1 m depth (MWL). After 10 weeks in the field (August 7th), the thalli were collected, and the growth increase in mm of the marked apical tips from above the floating bladder of the ten longest ramets on each thalli were measured. Data were analysed using Welch’s t-test in R (package BSDA), comparing growth at each site to the control site.
An additional transplantation study was performed in Gdansk Bay, Poland (Site 5, Figure 1A) during 1999–2000. Previously existing Fucus populations in Gdansk Bay, covering large areas within the bay, disappeared during the late 1970s as a result of eutrophication and pollution (Plinski and Florczyk 1984). The transplantation was made in order to determine which factors were most severely hindering re-establishment and to assess potential growth and natural recruitment. The transplantations of Fucus into Gdansk Bay were made through moving rocks with established adult thalli from Kalmar Sound, Sweden (Site 4, Figure 1A) with a similar salinity of ca. 7, and growing at similar depth (0.8–1 m MWL). The transplantations were made at two sites within Gdansk Bay (Figure 1C) where Fucus had previously occurred. Site I was in the port of Gdynia, which is the 3rd largest port in Poland. This site was strongly impacted by turbid, polluted water with a Secchi depth of about 1 m. Site J, located outside the village of Oslonino, was less eutrophicated but there were large impacts from sand abrasion, especially after strong winds due to recent shore erosion. Three attempts at transplantation were performed. In 1999, 100 thalli were placed at site I on September 9th, and 50 thalli were placed at site J on September 12th. On May 9th 2000, an additional 100 thalli were placed at site I. At all sites, Fucus tips were marked by punching a small hole in order to measure apical growth increase. After 12, 18 and 40 weeks, any remaining thalli were retrieved by SCUBA divers and brought ashore for measuring. The number of epiphytes and invertebrate fauna was studied at site I in Gdynia harbour, and is reported in Stachnik (2002 in Polish, abstract in English).
Results and discussion
Survival and growth of a naturally recruited Fucus population
The natural mortality was high in the studied population. The estimated density of germlings was 9600±500 per m2 after 1 year and reduced successively (Table 1). The frond length (cm) during the 5-year study shows the increasing growth in length over time and the formation of dichotomies. No dichotomies were produced during year 1 but, during the following years, the number more or less doubled each year (Table 1). The first pair of bladders was produced after 2 years during spring in some of the taller thalli (10–12 cm, 2–3 dichotomies). Only one frond was produced per holdfast during the study period of 5 years, suggesting that holdfasts with several fronds are older than 5 years. The first reproductive structures were recorded after 3 years when some of the fronds had reached a size of 26–36 cm. In year 5, most of the individuals produced reproductive structures, having reached frond lengths of 37–52 cm.
Fucus vesiculosus population development at a sheltered shallow site (H, Figure 1B) during the years 1992–1996, Askö. All numbers are mean values.
| Year | Age (months) | Total area sampled (dm2) | Number of holdfasts (mean/m2±s.e.) | Frond length (cm) (±s.e.) | Number of dichotomies (±s.e.) | n |
|---|---|---|---|---|---|---|
| 1992 | 12 | 0.5 | 9600±500 | 1.79 (±0.22) | 0 (±0) | 50 |
| 1993 | 24 | 1 | 853±123 | 5.88 (±0.36) | 0.93 (±0.13) | 60 |
| 1994 | 38 | 1.5 | 640±97 | 16.61 (±1.37) | 3.30 (±0.31) | 40 |
| 1995 | 50 | 6 | 480±88 | 19.35 (±1.55) | 4.00 (±0.35) | 30 |
| 1996 | 62 | 25 | 10 | 42.20 (±1.61) | 10.00 (±0.45) | 10 |
Figure 2A–D shows the relationship between thallus volume (cm3) and biomass (g DW), in thalli of different ages, and shows the large size differences within one generation. After 4 years, there is still a large proportion of small individuals (volume less than 50 cm3) occurring in the population, and only a few individuals have reached reproductive size.
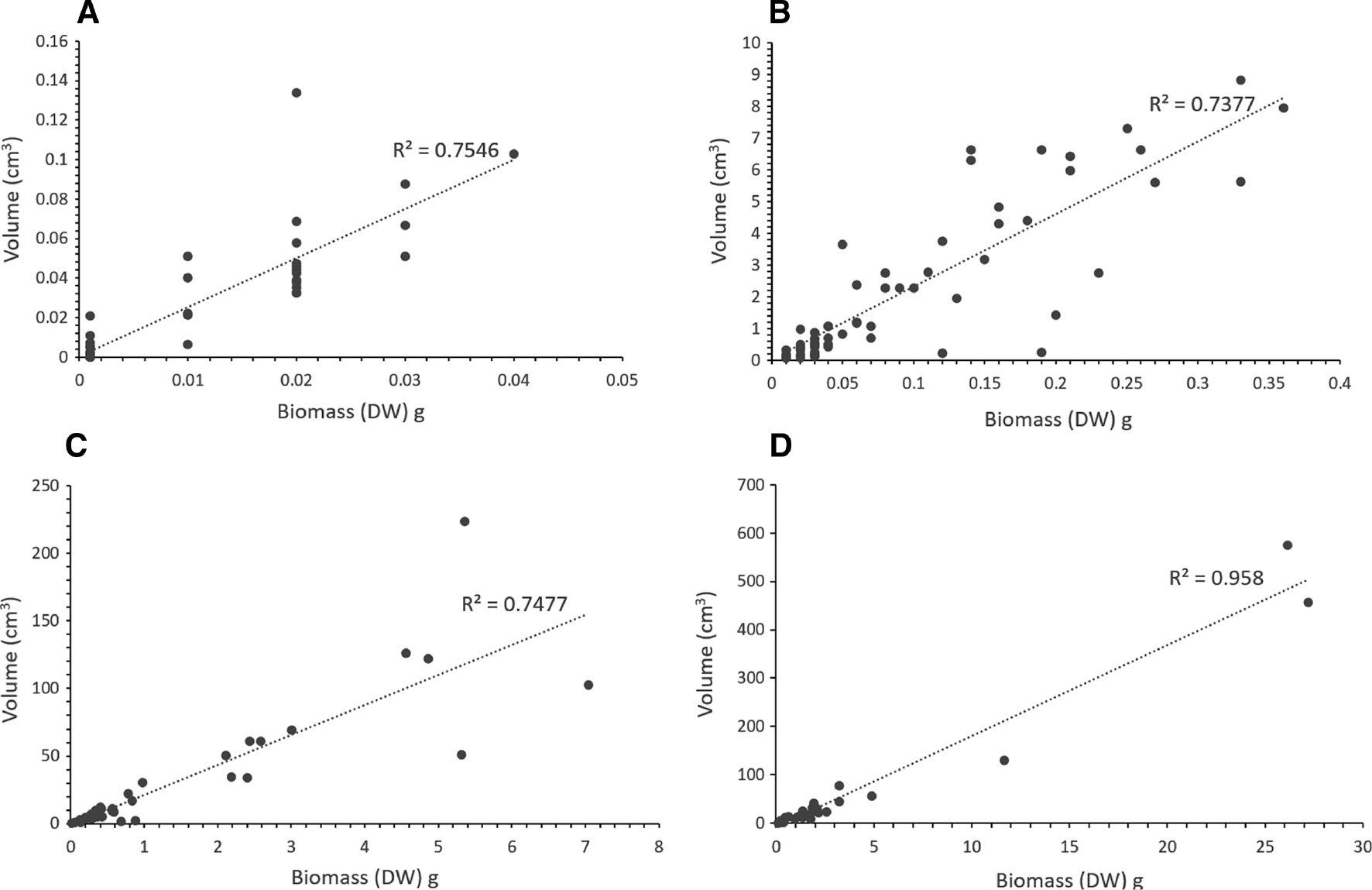
Fucus vesiculosus increase in size over time during 4 years.
Fucus vesiculosus: volume (calculated as a cone from thallus height and circumference) plotted against biomass (g dry weight) with linear regression for (A) 1-year-old, (B) 2-year-old, (C) 3-year-old and (D) 4-year-old thalli grown in the field at Askö during 1991–1994.
The maintenance and re-establishment of Fucus populations depend on several critical steps during early recruitment. One such step is the early survival of individual thalli and thus their potential to reach reproductive stage (Creed et al. 1996). The results from our 5-year survival and growth study of the naturally recruited Fucus vesiculosus population (Table 1, Figure 2) show high mortality in the early stages combined with only a few thalli reaching the reproductive stage after 4–5 years. According to Creed et al. (1996), marine F. vesiculosus populations have a “seed bank” of microscopic germlings, similar to many other seaweed species (Hoffmann and Santelices 1991, Worm et al. 2001) but this seems not to be the case in Baltic F. vesiculosus (Malm et al. 1999).
Vegetative generation, i.e. the ability to produce many new fronds from the same holdfasts, is more common in the Baltic Sea than in the North Sea (Malm and Kautsky 2004). If severed, the holdfasts produce a large number of fronds and regrowth is very fast (Ruuskanen and Bäck 1999, Malm and Kautsky 2004). This ability has contributed to fronds surviving ice scraping in the Baltic Sea (Kiirikki 1996b, Kiirikki and Ruuskanen 1996). For practical reasons, such as transportation, transplanting whole thalli might be unnecessary, as transplantations of only holdfasts on rocks may have the potential to produce large thalli capable of sexual reproduction within a season or two.
Effects of Fucus vegetation cover on Fucus settlement and survival
Regardless of whether the understorey was removed or kept intact, settlement and survival were around 10 times higher in the treatments containing Fucus thalli (the “Manipulated Fucus” and “Natural Fucus” treatments, see Materials and methods; Figure 3A), than in the Cladophora-covered substratum and the Cleaned treatments (Figure 3B). Only at the last measurement in November was there also a difference between the Cladophora-covered substratum and cleaned treatments (p=0.05). The two treatments containing Fucus did not differ at any time (Figure 3A). The survival was lowest (14%) in the treatment where the substratum was cleaned below the Fucus canopy. The reduction in survival was less dramatic on the disks placed below a natural Fucus stand (with 64% survival). Our results indicate a high survival below Fucus canopy, supporting previous studies, which reported enhanced recruitment under thinned fucoid canopies (Chapman 1989, 1990, Ang Jr 1991). A Fucus canopy may, through its sweeping and whiplash effects, hinder the establishment of competing filamentous algae and the settlement of sediment particles (Kiirikki 1996a). The whiplash effect may also reduce the number of grazing snails, favouring the growth of Fucus juveniles but a reduction of snail grazing could also reduce the top-down control on competing filamentous algae. In all treatments, except for the natural Fucus canopy with understorey growth, the number of juveniles was difficult to count due to large amounts of new dense growth of Cladophora, which occurred on the settlement disks before the sampling in late July. This situation possibly resulted in an underestimation of the number of Fucus juveniles at this sampling, explaining the higher number of individuals found on the subsequent sampling in August (Figure 3).

Boxplot showing number of Fucus vesiculosus juveniles per dm2 surviving from mid-July to early November, 1994 in Askö.
Treatments in (A) “Manipulated Fucus” without understorey (diagonal stripes) and “Natural Fucus (control)” with understorey (white) and (B) “Cladophora-covered substratum” (chequered) and “Cleaned substratum” cleared from both Fucus and understorey vegetation (vertical stripes). Note major difference in scales for y-axes in each panel. n=6 for all treatments. *indicates significant difference in density of juveniles between two treatments at one date (p<0.05). One-way ANOVA of treatments at each date showed a higher number of juveniles in treatments containing Fucus (i.e. Figure 3A) on all dates compared to treatments without Fucus (i.e. Figure 3B; p<0.001).
Effects of filamentous vegetation cover on Fucus settlement and survival
Settlement and survival of Fucus vesiculosus on new substratum were highest in the Control treatment of clean stones without any existing biofilm (Askö, 5 months, Figure 4A) and on Ceramic tiles (Räfsnäs, 3 months, Figure 4B) between June and September or November 2017. Pylaiella cover was detrimental and hindered settlement of juveniles almost completely at both sites. Low settlement was also noted for treatments Hildenbrandia and Brushed stone at Räfsnäs. The Brushed treatment, where filamentous algae were removed, resulted in higher recruitment of Fucus juveniles compared to the Pylaiella-covered substratum. The main recruitment period of summer-reproducing Fucus individuals in the northern Baltic Proper occurs just after the main growth period of Pylaiella, when the filamentous algae have high biomass and has begun detaching. This time of year, large areas of rocky substratum can be completely covered with a dense mat of loose lying filamentous algae (Qvarfordt 2006), presumably reducing the reproductive success of Fucus.

Boxplot showing number of Fucus vesiculosus juveniles per dm2 on five different types of substratum.
(A) Askö, after 5 months, n=10 except ceramic tile (n=7) and Hildenbrandia treatment (n=9). (B) Räfsnäs after 4 months, sample size n=6. Note major difference in scales for y-axes in each panel. Letters (A–E) above bars show groups that differ significantly (at p=0.05) within each site according to Tukey HSD post hoc test.
In nutrient enriched areas, sedimentation of carbon originating from primary production has increased five–10-fold (Jonsson and Carman 1994), negatively affecting establishment and survival of Fucus juveniles (Eriksson et al. 1998, Berger et al. 2003, Eriksson and Johansson 2003). Several studies (Heiskanen et al. 1998, Heiskanen and Tallberg 1999, Berger et al. 2003) show a clear seasonality in sedimentation rate, with one peak during November from re-suspension, and another peak in May and early June, originating from the deposition of the spring bloom. The latter occurs shortly before the main Fucus recruitment period in the Baltic Proper. Also, the combined effect of filamentous vegetation and sediment deposition, where a higher amount of sediment particles get trapped in Pylaiella and other filamentous species than on bare substratum, further inhibits Fucus settlement and survival (Eriksson and Johansson 2003).
The results suggest that, in order to facilitate for new recruitment, it would be better to provide new, clean substratum without any previous biofilm or algal cover, than to use already submerged substratum with an existing biofilm and cover of algae. Cleaning or brushing away filamentous algal cover from existing submerged rocky substrates before Fucus gametes settle, may also lead to an increased settlement success. We suggest this as a measure to be taken into consideration when planning a restoration activity in an area where the Fucus population has diminished or disappeared.
Time of substratum submersion is another important factor, which affects the species composition. Substratum submerged in October resulted in communities dominated by Pylaiella and a subsequent low Fucus recruitment (Qvarfordt 2006). Before any attempt to restore Fucus, the cover of Pylaiella or other common filamentous algae and level of sedimentation during the Fucus reproductive time at the proposed site should be taken into account (Worm et al. 1999, Worm et al. 2001, Berger et al. 2003). If populations of autumn reproductive Fucus are present in the area, they might be considered as seeding material since the rocky surfaces have a lower cover of Pylaiella during this season (Råberg et al. 2005). As growth rate in low temperatures is slow (Altamirano et al. 2003), new autumn produced germlings will be exposed to grazing by, e.g. Theodoxus fluviatilis and Radix balthica for a longer period than for summer germlings before reaching a large enough size to be safe from grazing (Malm et al. 1999), However, summer germlings are produced during the peak of grazing activity and might thus be exposed to a higher grazing pressure from the start.
The type of bedrock, e.g. sandstone compared to crystalline bedrock such as granite and amphibolite (Malm et al. 2003), and bedrock relief (Fletcher and Callow 1992) are other factors which should be considered when selecting substratum material, since large Fucus zygotes may have a higher survival and density on a rough surface than small green algal propagules.
Effects of exudate from filamentous algae and Hildenbrandia on Fucus germination
Exudate from four filamentous algae and the crust-forming Hildenbrandia were shown to have different negative effects on the germination of Fucus germlings during 7 days of exposure under laboratory conditions (Figure 5). The most pronounced effects were found in the treatments with exudate from the two red algal species, Ceramium and Hildenbrandia followed by the exudate of Pylaiella. In the treatments with exudates from the green algae Cladophora and Ulva, germination was higher but still significantly lower than in the control (p=0.001 and p<0.001, respectively). Ceramium has been documented to contain high amounts of polybrominated substances (Enhus et al. 2012) that may negatively affect the survival of Fucus germlings. When exposed to stress, such as grazing, strong light or low salinity as in the Baltic Sea, Ceramium was shown to produce higher levels of methoxylated diphenyl ethers and 2,3,6- tribromophenols than algae collected from the Swedish west coast (Enhus et al. 2012). In the northern Baltic Proper, Ceramium starts to grow in May–June and reaches the highest cover in July– August (Qvarfordt 2006). Similar substances have been found also in Pylaiella and Cladophora (Malmvärn et al. 2008, Löfstrand 2011). Contrary to our results, Chapman (1990) found no effect of Hildenbrandia on the recruitment density of Fucus distichus growing in littoral fringe pools or under the cover of ephemeral algae, again pointing to the complex interactions among macroalgae in marine habitats, with intense competition for space and different susceptibility to grazing (Lubchenco 1986).

Boxplot showing germination (%) of Fucus vesiculosus zygotes after 7 days of exposure under laboratory conditions to exudates from Cladophora glomerata, Ulva intestinalis, Pylaiella littoralis, Ceramium tenuicorne and Hildenbrandia rubra.
All treatments differed (p<0.001) from the control (one way ANOVA). n=6 for all treatments.
Transplantation trials
In the Trosa area, no growth was observed in the transplanted thalli at the innermost site (Site A), just outside the mouth of Trosa River (Figure 1B, Table 2). Also, all thalli were completely covered by the bryozoan Einhornia crustulenta Pallas and the barnacle Amphibalanus improvisus Darwin. The tissue colour of the thalli was black-greyish. At site B nearest to the outlet from the sewage treatment plant, the thalli were dark brown in colour but with living meristems. In site C the thalli were only slightly covered by E. crustulenta and showed the lowest growth. At sites C, D and E the thalli had a normal dark green colour. At sites D and E the thalli were covered by some epiphytic green algae. The thalli placed at sites G and F were black-brown and partly covered by E. crustulenta. Fucus thalli in the control site (H), close to the Askö Laboratory, did not have an epiphytic cover on the new growth, yet did not have the highest growth. The growth (Table 2) was higher at sites F (t=−2.40, p=0.047) and G (t=−2.41, p=0.043) than at the control site (H), and lower at site C (t=2.41, p=0.043). There were no significant differences in growth between sites B, D or E and site H.
Fucus vesiculosus growth after different time periods following thallus transplantation in Trosa (Sweden) site A–H and Gdansk Bay (Poland) sites I–J.
| Site | Placed in field | Period (weeks) | No. of thalli | Depth (m) | Salinity | Total N (mg l−1) | Total P (mg l−1) | Growth (cm period−1) (±SD) |
|---|---|---|---|---|---|---|---|---|
| A | May–August 2000 | 10 | 5 | 1 | 0.5 | 0.82 | 0.066 | 0 |
| B | 5 | 1 | 4.5 | 0.49 | 0.048 | 5.3±1.1 | ||
| C | 5 | 1 | 4.5 | 0.44 | 0.019 | 2.6±1.0 | ||
| D | 5 | 1 | 3.5 | 0.49 | 0.043 | 3.9±1.3 | ||
| E | 5 | 1 | 5.0 | 0.36 | 0.025 | 4.0±1.5 | ||
| F | 5 | 1 | 5.3 | 0.34 | 0.024 | 5.7±0.9 | ||
| G | 5 | 1 | 5.3 | 0.34 | 0.024 | 5.8±1.0 | ||
| H | 5 | 1 | 6.5 | – | – | 4.2±1.1 | ||
| I | September–December 1999 | 12 | 86 | 0.8 | 7.5 | – | – | ca 1–2 |
| I | May–September 2000 | 18 | 30 | 0.8 | 7.5 | – | – | ca 2–3 |
| J | September 1999–May 2000 | 40 | 17 | 0.8 | 7.5 | – | – | ca 1–1.5 |
The results of the transplantation experiment show that growth conditions at the innermost site A close to Trosa had too low a salinity for Fucus to survive. The results do not support any direct negative effects of high nutrient levels on Fucus growth transplanted at 1 m depth. Sites F and G show positive growth and only small amounts of epiphytes, mainly the green algae Ulva spp. and Cladophora spp., both commonly forming floating algal mats in nutrient-enriched waters (Vahteri et al. 2000). It should, however, be noted that nutrient data was measured the previous year before the transplantation study. Overall, however, the results suggest that a restoration attempt by transplantation might be possible in nutrient-enriched waters as long as the apical tips of Fucus with the meristem are not too heavily colonised by E. crustulenta and A. improvisus.
The growth and survival varied in the different studied areas depending on local conditions for several reasons. The transplantation studies (Table 2) emphasise that the local conditions affecting Fucus need to be identified for each specific site and taken into consideration to achieve a positive outcome for a transplantation effort.
In Gdansk Bay in Poland, the main reasons for poor survival of the transplanted thalli were most likely related to low growth at site I in the harbour of Gdynia (Table 2), as suggested by Stanchnik (2002). Apical growth in both sites was very low compared to growth reported from the Baltic Proper along the Swedish coast (Wallentinus 1979, Carlson 1991). Large amounts of Pylaiella settled on Fucus fronds, and Ulva spp. densely covered the stones in the first year (1999) at both Polish sites. Grazing was low, only five–six gammarids were found per thallus (Stanchnik 2002 in Polish). The second year (2000) in site I, all thalli transplanted the previous year had disappeared and a new transplantation was made in May. After the growth season, new fronds emerged from the holdfasts in September on the few remaining thalli. No grazing marks were noted, and only three–four gammarids and three–four Idotea chelipes were found per thallus (for details of fauna see Stanchnik 2002 in Polish). It is likely that shading by epiphytes, dominated by Pylaiella, Ulva spp., Einhornia and Amphibalanus, and an indirect effect of high nutrients and turbid water in the harbour resulted in low growth, and no surviving thalli after less than a year.
Compared to the rocky shores of Trosa archipelago, the Polish coast is very sandy, and the effect of sand abrasion was strong at site J (Figure 1C, Table 2). The stones with attached Fucus thalli got buried in the sand by wave action and had to be dug up by divers. Consequently, only a few thalli survived from September 1999 to May 2000 and the growth was very low (Table 2). The results from these transplantation trials suggest that the nutrient load will have to be further reduced before new transplantations and restoration experiments are attempted. It is worth considering that, at sites which have been hydrographically altered (e.g. by stone reef harvesting), thus changing the impact and patterns of waves or currents, any chance of restoring Fucus might be lost.
As a comparison, a replanting attempt for Fucus vesiculosus was performed during 1996–1999 in the area outside Mönsterås, Kalmar Sound, after the chlorine emissions from the pulp industry had ceased (Wikström et al. 2000). Already after 4 months, the 200 transplanted thalli showed signs of heavy grazing from high Idotea balthica abundance (Nilsson et al. 1999, Engkvist et al. 2004). A few new juvenile recruits were recorded after 1 year, but the grazing by the gastropods Theodoxus fluviatilis and Radix balthica on small germlings recruited from autumn reproducing thalli was high and, after another 2 years, no thalli or juveniles were left (Malm et al. 1999). According to Worm et al. (1999), grazing snails may graze down the filamentous algae and thereby promote successful recruitment of Fucus. However, at a high nutrient load, the filamentous algal growth is too fast. Thus, summer reproducing thalli may be limited by higher amounts of Pylaiella during their recruitment period (Qvarfordt 2006). Similar grazing damage from Idotea has been reported from other areas in the Kalmar sound on the Swedish east coast, here suggested to be caused by low fish predation (Engkvist et al. 2000). As observed in these transplantation examples with adult thalli, differences in environmental factors, such as low light availability in highly nutrient enriched areas, high abundance of grazers and/or overgrowth by epiphytes may reduce or hinder a successful re-establishment, alone or in combination.
Conclusion
The studies and summary of transplantation trials in our paper show that to maintain and re-establish Fucus populations, critical steps during early recruitment and adult survival of Fucus need to be assessed, taking into consideration the actual local conditions at the site in question. Here we present the first study showing that it will take 4–5 years until a sexually recruited thallus reaches reproductive age, clearly pointing to the slow process in recovery of Fucus stands in the Baltic Sea. The slow growth suggests that seeding should be done in combination with transplantation of adult thalli in eutrophicated environments, as the presence of adult fronds increases juvenile survival. Under these conditions, recruitment and re-establishment may be further improved by using new substrata, i.e. without biofilm, or by brushing submerged rocky substrata before Fucus gametes settle. Using transplantation material from both summer- and autumn-reproducing Fucus populations further increases the chances of germling survival depending on the site-specific grazing pressure and the cover of Pylaiella. Finally, as observed in the few transplantation trials performed with adult thalli, local differences in environmental factors will have a tremendous impact on the success. There is an obvious need to consider, and measure, factors such as salinity, light and nutrient loads together with biotic factors, e.g. grazing pressure, sediment, and epiphytic cover, when planning a restoration activity in an area where the Fucus population has been reduced or has completely disappeared.
About the authors

Lena Kautsky is Professor Emeritus in Marine Plant Ecology at Stockholm University, Department of Ecology, Environment and Plant Science and senior advisor at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on benthic vegetation and dynamics in Fucus communities. Furthermore, her research covers studies of biodiversity both at the genetic and species level, as well as effects and impact of pollution on benthic vegetation and issues on integrated aquaculture and restoration of shallow benthic communities

Susanne Qvarfordt has a PhD in Baltic Sea Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science. Her research is mainly focused on vegetation in the Baltic Sea, both algae and vascular communities, and how these communities vary over time.

Ellen Schagerström has a PhD in Plant Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science and at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on biodiversity both at the genetic and species level in Fucus communities as well as adaptation and speciation. Furthermore, her research covers studies of dynamics of benthic vegetation as well as restoration of shallow benthic communities.
Acknowledgements
This paper was funded by the Baltic Sea 2020 foundation. The authors wish to acknowledge Claes Dannbeck and the many people involved over the years in these projects and studies here summarised. We are very grateful to S. Wikström for valuable input. The study in Poland (L. Kautsky) was supported by WWF Sweden.
References
Alexandridis, N., A. Oschlies and M. Wahl. 2012. Modeling the effects of abiotic and biotic factors on the depth distribution of Fucus vesiculosus in the Baltic Sea. Mar. Ecol. Prog. Ser. 463: 59–72.10.3354/meps09856Search in Google Scholar
Altamirano, M.A., A. Flores-Moya and F.L. Figueroa. 2003. Effects of UV radiation and temperature on growth of germlings of three species of Fucus (Phaeophyceae). Aquat. Bot. 75: 9–20.10.1016/S0304-3770(02)00149-3Search in Google Scholar
Andersen, J.H., J. Carstensen, D.J. Conley, K. Dromph, V. Fleming-Lehtinen, B.G. Gustafsson, A.B. Josefson, A. Norkko, A. Villnäs and C. Murray. 2017. Long-term temporal and spatial trends in eutrophication status of the Baltic Sea. Biol. Rev. 92: 135–149.10.1111/brv.12221Search in Google Scholar PubMed
Andersson, S., L. Kautsky and A. Kalvas. 1994. Circadian and lunar gamete release in Fucus vesiculosus in the atidal Baltic Sea. Mar. Ecol. Prog. Ser. 110: 195–201.10.3354/meps110195Search in Google Scholar
Andersson, S. and L. Kautsky. 1996. Copper effects on reproductive stages of Baltic Sea Fucus vesiculosus. Mar. Biol. 125: 171–176.10.1007/BF00350771Search in Google Scholar
Ang Jr, P.O. 1991. Natural dynamics and matrix models of a Fucus distichus (Phaeophyceae, Fucales) population in Vancouver, British Columbia, Canada. The University of British Columbia, Vancouver.10.3354/meps078071Search in Google Scholar
Bäck, S., J.C. Collins and G. Russell. 1991. Aspects of the reproductive biology of Fucus vesiculosus from the coast of SW Finland. Ophelia 34: 129–141.10.1080/00785326.1991.10429701Search in Google Scholar
Berger, R., T. Malm and L. Kautsky. 2001. Two reproductive strategies in Baltic Fucus vesiculosus (Phaeophyceae). Eur. J. Phycol. 36: 265–273.10.1080/09670260110001735418Search in Google Scholar
Berger, R., E. Henriksson, L. Kautsky and T. Malm. 2003. Effects of filamentous algae and deposited matter on the survival of Fucus vesiculosus L. germlings in the Baltic Sea. Aquat. Ecol. 37: 1–11.10.1023/A:1022136900630Search in Google Scholar
Berger, R., L. Bergström, E. Graneli and L. Kautsky. 2004. How does eutrophication affect different life stages of Fucus vesiculosus in the Baltic Sea? a conceptual model. Hydrobiologia 514: 243–248.10.1007/978-94-017-0920-0_22Search in Google Scholar
Bergström, U., G. Sundblad, A.L. Downie, M. Snickars, C. Boström and M. Lindegarth. 2013. Evaluating eutrophication management scenarios in the Baltic Sea using species distribution modelling. J. Appl. Ecol. 50: 680–690.10.1111/1365-2664.12083Search in Google Scholar
Carlson, L. 1991. Seasonal variation in growth, reproduction and nitrogen content of Fucus vesiculosus L. in the Öresund, Southern Sweden. Bot. Mar. 34: 447–454.10.1515/botm.1991.34.5.447Search in Google Scholar
Chapman, A. 1989. Abundance of Fucus spiralis and ephemeral seaweeds in a high eulittoral zone: effects of grazers, canopy and substratum type. Mar. Biol. 102: 565–572.10.1007/BF00438359Search in Google Scholar
Chapman, A.R.O. 1990. Effects of grazing, canopy cover and substratum type on the abundances of common species of seaweeds inhabiting littoral fringe tide pools. Bot. Mar. 33: 319–326.10.1515/botm.1990.33.4.319Search in Google Scholar
Chapman, A.R.O. 1995. Functional ecology of fucoid algae: twenty-three years of progress. Phycologia 34: 1–32.10.2216/i0031-8884-34-1-1.1Search in Google Scholar
Creed, J.C., T.A. Norton and S.P. Harding. 1996. The development of size structure in a young Fucus serratus population. Eur. J. Phycol. 31: 203–209.10.1080/09670269600651391Search in Google Scholar
Engkvist, R., T. Malm and S. Tobiasson. 2000. Density dependent grazing effects of the isopod Idotea baltica Pallas on Fucus vesiculosus L. in the Baltic Sea. Aquat Ecol. 34: 253–260.10.1023/A:1009919526259Search in Google Scholar
Engkvist, R., J. Nilsson and S. Tobiasson. 2002. Temporal and geographical patterns of distribution of the brown macroalgae F. vesiculosus and F. serratus in the counties of Blekinge and Kalmar, SE Sweden. ISSN:1402-6198, Rapport 2002:4 (in Swedish).Search in Google Scholar
Engkvist, R., T. Malm and J. Nilsson. 2004. Interaction between isopod grazing and wave action: a structuring force in macroalgal communities in the southern Baltic Sea. Aquat. Ecol. 38: 403–413.10.1023/B:AECO.0000035162.07481.1fSearch in Google Scholar
Enhus, C., E. Boalt, D. Lindqvist, B. Eklund and L. Asplund. 2012. Natural production of brominated substances in the red algae Ceramium tenuicorne. 2:2012, Swedish Museum of Natural History, Stockholm (in Swedish).Search in Google Scholar
Eriksson, B.K. and G. Johansson. 2003. Sedimentation reduces recruitment success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. Eur. J. Phycol. 38: 217–222.10.1080/0967026031000121688Search in Google Scholar
Eriksson, B.K., G. Johansson and P. Snoeijs. 1998. Long-term changes in the sublittoral zonation of brown algae in the southern Bothnian Sea. Eur. J. Phycol. 33: 241–249.10.1080/09670269810001736743Search in Google Scholar
Fletcher, R.L. and M.E. Callow. 1992. The settlement, attachment and establishment of marine algal spores. Brit. Phycol. J. 27: 303–329.10.1080/00071619200650281Search in Google Scholar
Haahtela, I. 1984. A hypothesis of the decline of the bladder wrack (Fucus vesiculosus L.) in SW Finland in 1975–1981. Limnologica 15: 345–350.Search in Google Scholar
Heiskanen, A.-S. and P. Tallberg. 1999. Sedimentation and particulate nutrient dynamics along a coastal gradient from a fjord-like bay to the open sea. Hydrobiologia 393: 127–140.10.1007/978-94-017-0912-5_13Search in Google Scholar
Heiskanen, A., J. Haapala and K. Gundersen. 1998. Sedimentation and pelagic retention of particulate C, N and P in the coastal northern Baltic Sea. Estuar. Coast. Shelf S. 46: 703–712.10.1006/ecss.1997.0320Search in Google Scholar
Hemmi, A., A. Mäkinen, V. Jormalainen and T. Honkanen. 2005. Responses of growth and phlorotannins in Fucus vesiculosus to nutrient enrichment and herbivory. Aquat. Ecol. 39: 201–211.10.1007/s10452-004-3526-zSearch in Google Scholar
Hill, R., A. Bellgrove, P.I. Macreadie, K. Petrou, J. Beardall, A. Steven and P.J. Ralph. 2015. Can macroalgae contribute to blue carbon? An Australian perspective. Limnol. Oceanogr. 60: 1689–1706.10.1002/lno.10128Search in Google Scholar
Hoffmann, A.J. and B. Santelices. 1991. Banks of algal microscopic forms: hypotheses on their functioning and comparisons with seed banks. Mar. Ecol. Prog. Ser. 79: 185–194.10.3354/meps079185Search in Google Scholar
Johannesson, K. and C. André. 2006. Life on the margin: genetic isolation and diversity loss in a peripheral marine ecosystem, the Baltic Sea. Mol. Ecol. 15: 2013–2029.10.1111/j.1365-294X.2006.02919.xSearch in Google Scholar
Jonsson, P. and R. Carman. 1994. Changes in deposition of organic matter and nutrients in the Baltic Sea during the twentieth century. Mar. Pollut. Bull. 28: 417–426.10.1016/0025-326X(94)90127-9Search in Google Scholar
Kangas, P. and A. Niemi. 1985. Observations of recolonization by the bladderwrack, Fucus vesiculosus, on the southern coast of Finland. Aqua Fenn.15: 133–141.Search in Google Scholar
Kangas, P., H. Autio, G. Hällfors, H. Luther, A. Niemi and H. Salemaa. 1982. A general model of the decline of Fucus vesiculosus at Tvärminne, South Coast of Finland, in 1977–81. Acta Bot. Fenn. 118: 1–27.Search in Google Scholar
Kautsky, H. 1992. The impact of pulp-mill effluents on phytobenthic communities in the Baltic Sea. Ambio 21: 308–313.Search in Google Scholar
Kautsky, N., H. Kautsky, U. Kautsky and M. Waern. 1986. Decreased depth penetration of Fucus vesiculosus (L) since the 1940s indicates eutrophication of the Baltic Sea. Mar. Ecol. Prog. Ser. 28: 1–8.10.3354/meps028001Search in Google Scholar
Kiirikki, M. 1996a. Experimental evidence that Fucus vesiculosus (Phaeophyta) controls filamentous algae by means of the whiplash effect. Eur J. Phycol. 31: 61–66.10.1080/09670269600651201Search in Google Scholar
Kiirikki, M. 1996b. Mechanisms affecting macroalgal zonation in the northern Baltic Sea. Eur J. Phycol. 31: 225–232.10.1080/09670269600651421Search in Google Scholar
Kiirikki, M. and A. Ruuskanen. 1996. How does Fucus vesiculosus survive ice scraping? Bot. Mar. 39: 133–139.10.1515/botm.1996.39.1-6.133Search in Google Scholar
Korpinen, S., V. Jormalainen and T. Honkanen. 2007. Effects of nutrients, herbivory, and depth on the macroalgal community in the rocky sublittoral. Ecology 88: 839–852.10.1890/05-0144Search in Google Scholar PubMed
Kukk, H. and G. Martin. 1992. Long-term dynamics of the phytobenthos in Pärnu Bay, the Baltic Sea. Proc. Estonian Acad. Sci. Ecol. 2: 110–118.10.3176/ecol.1992.3.04Search in Google Scholar
Lago-Leston, A., C. Mota, L. Kautsky and G.A. Pearson. 2010. Functional divergence in heat shock response following rapid speciation of Fucus spp. in the Baltic Sea. Mar. Biol. 157: 683–688.10.1007/s00227-009-1348-1Search in Google Scholar
Larsson, U., R. Elmgren and F. Wulff. 1985. Eutrophication in the Baltic Sea: causes and consequences. Ambio 14: 9–14.Search in Google Scholar
Lindvall, B. 1984. The condition of a Fucus-community in a polluted archipelago area on the east coast of Sweden. Ophelia 3: 147–150.Search in Google Scholar
Löfstrand, K. 2011. Trends and exposure of naturally produced brominated substances in Baltic biota-with focus on OH-PBDEs, MeO-PBDEs and PBDDs. PhD thesis, Department of Materials and Environmental Chemistry (MMK), Stockholm University, Sweden.Search in Google Scholar
Lubchenco, J. 1986. Relative importance of competition and predation: early colonization by seaweeds in New England. In Community ecology. Harper and Row, New York pp. 537–555.Search in Google Scholar
Maczassek, K. 2014. Environmental drivers of fertility, fertilization and germination of Fucus vesiculosus on the German coast. PhD thesis, Christian Albrechts University, Kiel, Germany.Search in Google Scholar
Malm, T. and L. Kautsky. 2003. Differences in life-history characteristics are consistent with the vertical distribution pattern of Fucus serratus and Fucus vesiculosus (Fucales, Phaeophyceae) in the central Baltic Sea. J. Phycol. 39: 880–887.10.1046/j.1529-8817.2003.02115.xSearch in Google Scholar
Malm, T. and L. Kautsky. 2004. Are Bladderwrack (Fucus vesiculosus L.) holdfasts that support several fronds composed of one or several genetic individuals? Aquat. Bot. 80: 221–226.10.1016/j.aquabot.2004.10.001Search in Google Scholar
Malm, T., R. Engkvist and L. Kautsky. 1999. Grazing effects of two freshwater snails on juvenile Fucus vesiculosus in the Baltic Sea. Mar. Ecol. Prog. Ser. 188: 63–71.10.3354/meps188063Search in Google Scholar
Malm, T., L. Kautsky and R. Engkvist. 2001. Reproduction, recruitment and geographical distribution of Fucus serratus L. in the Baltic Sea. Bot. Mar. 44: 101–108.10.1515/BOT.2001.014Search in Google Scholar
Malm, T., L. Kautsky and T. Claesson. 2003. The density and survival of Fucus vesiculosus L. (Fucales, Phaeophyta) on different bedrock types on a Baltic Sea moraine coast. Bot. Mar. 46: 256–262.10.1515/BOT.2003.023Search in Google Scholar
Malmvärn, A., Y. Zebühr, L. Kautsky, Å. Bergman and L. Asplund. 2008. Hydroxylated and methoxylated polybrominated diphenyl ethers and polybrominated dibenzo-p-dioxins in red alga and cyanobacteria living in the Baltic Sea. Chemosphere 72: 910–916.10.1016/j.chemosphere.2008.03.036Search in Google Scholar PubMed
Nilsson, J., R. Engkvist and L.-E. Persson. 1999. Restoration of Coastal environments. Dnr. (1699-2/95), (2063/96) Slutrapport 1996-1999, 1-17 (In Swedish).Search in Google Scholar
Nilsson, J., R. Engkvist and L.E. Persson. 2004. Long-term decline and recent recovery of Fucus populations along the rocky shores of southeast Sweden, Baltic Sea. Aquat. Ecol. 38: 587–598.10.1007/s10452-004-5665-7Search in Google Scholar
Olenin, S. and K. Klovaité. 1998. Introduction to the marine and coastal environment of Lithuania. Red List of Marine and Coastal Biotopes and Biotope Complexes of the Baltic Sea. Nordheim Hv., Boedeker D (eds). HELCOM. Baltic Sea Environ. Proc. 75: 39–43.Search in Google Scholar
Oswald, R., N. Telford, R. Seed and C. Happey-Wood. 1984. The effect of encrusting bryozoans on the photosynthetic activity of Fucus serratus L. Estuar. Coast. Shelf S. 19: 697–702.10.1016/0272-7714(84)90024-6Search in Google Scholar
Pearson, G.A., E.A. Serrão and S.H. Brawley. 1998. Control of gamete release in fucoid algae: Sensing hydrodynamic conditions via carbon acquisition. Ecology 79: 1725–1739.10.1890/0012-9658(1998)079[1725:COGRIF]2.0.CO;2Search in Google Scholar
Pearson, G., L. Kautsky and E. Serrão. 2000. Recent evolution in Baltic Fucus vesiculosus: reduced tolerance to emersion stresses compared to intertidal (North Sea) populations. Mar. Ecol. Prog. Ser. 202: 67–79.10.3354/meps202067Search in Google Scholar
Plinski, M. and I. Florczyk. 1984. Changes in the phytobenthos resulting from the eutrophication of the Puck Bay. Limnologica 15: 325–327.Search in Google Scholar
Qvarfordt, S. 2006. Phytobenthic communities in the Baltic Sea-seasonal patterns in settlement and succession. PhD thesis, Stockholm University. Stockholm, Sweden.Search in Google Scholar
R Development Core team. 2014. R: A language and environment for statistical computing. Retrieved from http://www.R-project.org/.Search in Google Scholar
Råberg, S., R. Berger-Jönsson, A. Bjorn, E. Graneli and L. Kautsky. 2005. Effects of Pilayella littoralis on Fucus vesiculosus recruitment: implications for community composition. Mar. Ecol. Prog. Ser. 289: 131–139.10.3354/meps289131Search in Google Scholar
Rohde, S., C. Hiebenthal, M. Wahl, R. Karez and K. Bischof. 2008. Decreased depth distribution of Fucus vesiculosus (Phaeophyceae) in the Western Baltic: effects of light deficiency and epibionts on growth and photosynthesis. Eur. J. Phycol. 43: 143–150.10.1080/09670260801901018Search in Google Scholar
Rönnbäck, P., N. Kautsky, L. Pihl, M. Troell, T. Söderqvist and H. Wennhage. 2007. Ecosystem goods and services from Swedish coastal habitats: identification, valuation, and implications of ecosystem shifts. AMBIO 36: 534–544.10.1579/0044-7447(2007)36[534:EGASFS]2.0.CO;2Search in Google Scholar
Rönnberg, O., J. Lehto and I. Haahtela. 1985. Recent changes in the occurrence of Fucus vesiculosus in the Archipelago Sea, SW Finland. Ann. Bot. Fenn. pp. 231–244.Search in Google Scholar
Rosemarin, A., J. Mattsson, K. Lehtinen, M. Notini and E. Nylen. 1986. Effects of pulp-mill chlorate (ClO3-) on Fucus vesiculosus-a summary of projects. Ophelia pp. 219–224.Search in Google Scholar
Russell, G. 1985. Recent evolutionary changes in the algae of the Baltic Sea. Brit. Phycol. J. 20: 87–104.10.1080/00071618500650111Search in Google Scholar
Ruuskanen, A. and S. Bäck. 1999. Does environmental stress affect fertility and frond regeneration of Fucus vesiculosus? Ann. Bot. Fenn. 36: 285–290.Search in Google Scholar
Sandén, P. and B. Håkansson. 1996. Long-term trends in Secchi depth in the Baltic Sea. Limnol. Oceanogr. 41: 346–351.10.4319/lo.1996.41.2.0346Search in Google Scholar
Schagerström, E., H. Forslund, L. Kautsky, M. Pärnoja and J. Kotta. 2014. Does thalli complexity and biomass affect the associated flora and fauna of two co-occurring Fucus species in the Baltic Sea? Estuar. Coast. Shelf S. 149: 187–193.10.1016/j.ecss.2014.08.022Search in Google Scholar
Schramm, W. and P. Nienhuis. 1996. Marine Benthic Vegetation: Recent Changed and Effects of Eutrophication. Ecological Studies 123. Springer Science & Business Media, Berlin Heidelberg, pp. 1–472.10.1007/978-3-642-61398-2Search in Google Scholar
Serrão, E.A., L. Kautsky and S.H. Brawley. 1996a. Distributional success of the marine seaweed Fucus vesiculosus L in the brackish Baltic Sea correlates with osmotic capabilities of Baltic gametes. Oecologia 107: 1–12.10.1007/BF00582229Search in Google Scholar PubMed
Serrão, E.A., G. Pearson, L. Kautsky and S.H. Brawley. 1996b. Successful external fertilization in turbulent environments. P. Natl. Acad. Sci. USA 93: 5286–5290.10.1073/pnas.93.11.5286Search in Google Scholar PubMed PubMed Central
Serrão, E.A., S.H. Brawley, J. Hedman, L. Kautsky and G. Samuelson. 1999. Reproductive success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. J. Phycol. 35: 254–269.10.1046/j.1529-8817.1999.3520254.xSearch in Google Scholar
Snoeijs, P. 1999. Marine and brackish waters. Acta Phytogeogr. Suec. 84: 187–212.Search in Google Scholar
Stachnik, M. 2002. Sukcesja BezkrEgowóc w populach Fucus vesiculosus w eksperymenchie translokacyjnym w zatoce Gdańskiej. Uniwesytet Gdanski Wydzial Biologii, Geografii I Oceanologii. Report – Abstract in English pp. 1–51.Search in Google Scholar
Torn, K., D. Krause-Jensen and G. Martin. 2006. Present and past depth distribution of bladderwrack (Fucus vesiculosus) in the Baltic Sea. Aquat. Bot. 84: 53–62.10.1016/j.aquabot.2005.07.011Search in Google Scholar
Vahteri, P. and I. Vuorinen. 2016. Continued decline of the bladderwrack, Fucus vesiculosus, in the Archipelago Sea, northern Baltic proper. Boreal Environ. Res. 21: 373–386.Search in Google Scholar
Vahteri, P., A. Mäkinen, S. Salovius and I. Vuorinen. 2000. Are drifting algal mats conquering the bottom of the Archipelago Sea, SW Finland? AMBIO 29: 338–343.10.1579/0044-7447-29.6.338Search in Google Scholar
Vogt, H. and W. Schramm. 1991. Conspicuous decline of Fucus in Kiel Bay (Western Baltic) - What are the causes Mar. Ecol. Prog. Ser. 69: 189–194.10.3354/meps069189Search in Google Scholar
Waern, M. 1952. Rocky- shore algae in the Öregrund archipelago. PhD thesis, Uppsala University, Uppsala, Sweden.Search in Google Scholar
Wahl, M., V. Jormalainen, B.K. Eriksson, J.A. Coyer, M. Molis, H. Schubert, M. Dethier, R. Karez, I. Kruse, M. Lenz, G. Pearson, S. Rohde, S.A. Wikström and J.L. Olsen. 2011. Chapter Two – Stress Ecology in Fucus: Abiotic, Biotic and Genetic Interactions. In (L. Michael ed) Advances in Marine Biology. Academic Press. pp. 37–105.10.1016/B978-0-12-385536-7.00002-9Search in Google Scholar
Wallentinus, I. 1979. Environmental influences on benthic macrovegetation in the Trosa-Askö area, northern Baltic Proper. II. The ecology of macroalgae and submersed phanerogams. Askö Laboratory, Department of Botany, University of Stockholm, Stockholm, Sweden.Search in Google Scholar
Wikström, S.A. and L. Kautsky. 2007. Structure and diversity of invertebrate communities in the presence and absence of canopy-forming Fucus vesiculosus in the Baltic Sea. Estuar. Coast. Shelf S. 72: 168–176.10.1016/j.ecss.2006.10.009Search in Google Scholar
Wikström, S., L. Kautsky and T. Malm. 2000. The effects of chlorine-free pulp mill effluents on reproduction and grazing interactions in Baltic Sea Fucus vesiculosus L. Ophelia 53: 173–179.10.1080/00785326.2000.10409447Search in Google Scholar
Worm, B. and H.K. Lotze. 2006. Effects of eutrophication, grazing, and algal blooms on rocky shores. Limnol. Oceanogr. 51: 569–579.10.4319/lo.2006.51.1_part_2.0569Search in Google Scholar
Worm, B., H. Lotze, C. Boström, R. Engkvist, V. Labanauskas and U. Sommer. 1999. Marine diversity shift linked to interactions among grazers, nutrients and propagule banks. Mar. Ecol. Prog. Ser. 185: 309–314.10.3354/meps185309Search in Google Scholar
Worm, B., H.K. Lotze and U. Sommer. 2001. Algal propagule banks modify competition, consumer and resource control on baltic rocky shores. Oecologia 128: 281–293.10.1007/s004420100648Search in Google Scholar PubMed
©2019 Lena Kautsky et al., published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 License.
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Baltic algae
- Physiology and ecology
- Limited response of a spring bloom community inoculated with filamentous cyanobacteria to elevated temperature and pCO2
- Fucus vesiculosus adapted to a life in the Baltic Sea: impacts on recruitment, growth, re-establishment and restoration
- Acclimation limits of Fucus evanescens along the salinity gradient of the southwestern Baltic Sea
- Interactive effects of temperature and light on reattachment success in the brown alga Fucus radicans
- Phylogeny and biogeography
- Surveying seaweeds from the Ulvales and Fucales in the world’s most frequently used artificial waterway, the Kiel Canal
- New records from the southern North Sea and first records from the Baltic Sea of Kornmannia leptoderma
- Genetic variability of charophyte algae in the Baltic Sea area
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Baltic algae
- Physiology and ecology
- Limited response of a spring bloom community inoculated with filamentous cyanobacteria to elevated temperature and pCO2
- Fucus vesiculosus adapted to a life in the Baltic Sea: impacts on recruitment, growth, re-establishment and restoration
- Acclimation limits of Fucus evanescens along the salinity gradient of the southwestern Baltic Sea
- Interactive effects of temperature and light on reattachment success in the brown alga Fucus radicans
- Phylogeny and biogeography
- Surveying seaweeds from the Ulvales and Fucales in the world’s most frequently used artificial waterway, the Kiel Canal
- New records from the southern North Sea and first records from the Baltic Sea of Kornmannia leptoderma
- Genetic variability of charophyte algae in the Baltic Sea area