Interactive effects of temperature and light on reattachment success in the brown alga Fucus radicans
-
Ellen Schagerström
Ellen Schagerström has a PhD in Plant Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science and at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on biodiversity both at the genetic and species level in
Fucus communities as well as adaptation and speciation. Furthermore, her research covers studies of dynamics of benthic vegetation as well as restoration of shallow benthic communities. and
Tiina Salo
and
Tiina Salo
Tiina Salo works on understanding ecological and eco-evolutionary effects of stress on individuals, populations and communities. She uses macroalgae, seagrasses and invertebrates as her model organisms to study organismal and community traits. She is currently a post-doctoral fellow at Stockholm University, Sweden.

Abstract
Fucus radicans is an endemic habitat-forming brown macroalga in the Baltic Sea that commonly complements its sexual reproduction with asexual reproduction. Asexual reproduction in F. radicans takes place through formation of adventitious branches (hereafter fragments), but the exact mechanisms behind it remain unknown. We assessed experimentally the importance of two environmental factors determining the re-attachment success of F. radicans fragments. By combining different light conditions (daylength and irradiance; high or low light) and water temperature (+14°C and +4°C), we mimicked ambient light and temperature conditions of winter, spring/autumn and summer for F. radicans. Fragments were able to re-attach in all tested conditions. Temperature and light had an interactive impact on re-attachment: the combination of high temperature and high light level resulted in the highest re-attachment success, while light level had no effects on re-attachment success in cooler water temperature and the re-attachment success in high temperature under low light levels was very low. The results suggest that rhizoid formation, and thus re-attachment success, may depend on the net primary production (metabolic balance) of the fragment. However, whether the re-attachment and asexual reproduction success simply depends on photosynthetic capacity warrants further mechanistic studies. Understanding the mechanisms of asexual reproduction in F. radicans is important in order to assess the dispersal capacity of this foundation species.
Introduction
Asexual reproduction through vegetative propagation is most common in sub-optimal conditions, suggesting an evolutionary trait-environment relationship (Klimeš et al. 1997). Some marine macroalgae are known to complement their sexual reproduction by dispersal of fragments that later re-attach, forming new thalli (Santelices 2004). Such asexual reproduction has been reported in e.g. Codium spp. (Gagnon et al. 2014), Caulerpa spp. (Ceccherelli and Piazzi 2001), Halimeda spp. (Walters et al. 2002), Gracillaria chilensis (Santelices and Varela 1993, Santelices et al. 1995) and Bostrychia radicans (Collado-Vides 2001).
The Baltic Sea is a marine marginal environment with low yet stable salinity (Johannesson and Andre 2006) and many macroalgal species have adapted to the environmental conditions in the area by alterations in their life cycles. For example, Furcellaria lumbricalis (Kostamo 2008) and Ceramium tenuicorne (Bergström et al. 2003) have a higher proportion of vegetative reproduction in the northern Baltic Sea compared to more central populations. Asexual reproduction in the brown macroalgal genus Fucus was recently discovered in the Bothnian Sea basin in the Baltic Sea by Tatarenkov et al. (2005), who described vegetative reproduction of Fucus vesiculosus and Fucus radicans. Several studies have documented the physiological adaptations to low salinity of fucoids in the Baltic Sea in comparison to Atlantic populations. Differences in the reproductive traits show adaptations in gametes to low salinity and the lack of a tidal induced gamete release trigger (i.e. Andersson et al. 1994, Serrão et al. 1996, 1997, 1999) but other physiological differences such as lower photosynthesis, lower optimal temperatures and lower growth have also been documented (i.e. Raven and Samuelsson 1988, Nygård and Ekelund 2006, Nygård and Dring 2008, Gylle et al. 2011). The Baltic fucoids also tend to have a lower tolerance to desiccation compared to the more central Atlantic populations (Pearson et al. 2000).
Fucus radicans is a foundation species in the species poor Baltic Sea ecosystem, providing a perennial structure and habitat (Dayton 1972). It is endemic to the Baltic Sea (Pereyra et al. 2009) and only found in the Bothnian Sea and around the Estonian island of Saaremaa, within a salinity range of 3–5 in the Bothnian Sea and 5–6 around Estonia (Feistel et al. 2010, Forslund et al. 2012). Fucus radicans can exhibit high, up to 84%, clonality compared to e.g. Fucus vesiculosus where clonality of 7% has been described but only within a few marginal populations (Bergström et al. 2005, Tatarenkov et al. 2005, Johannesson et al. 2011). The high clonality is unusual among fucoids and, as it has not been described elsewhere, it indicates that asexual dispersal clearly is advantageous for reproduction in F. radicans populations in low saline environments (Tatarenkov et al. 2005, Johannesson et al. 2011).
Asexual reproduction in Fucus radicans takes place through detached adventitious branches (called fragments once severed from the parental thallus). These adventitious branches are numerous (ca. 8 adventitious branches per g wet weight thallus; Forslund and Kautsky 2013) and most often formed in the lower, older parts of the thallus, but can also be found higher up, emerging from the midrib (see Tatarenkov et al. 2005 for illustrations). The adventitious branches are presumably severed from the main thalli and become free-floating fragments either by wave action or grazing and re-attach to the substrate by forming new rhizoids from the severed basal part (Tatarenkov et al. 2005). As sexual reproduction in F. radicans is likely to be as spatially limited as in Fucus vesiculosus (sexual dispersal range of a couple of meters; Serrão et al. 1997), the large distances between thalli of the same clone (Johannesson et al. 2011) suggest that asexual reproduction has the potential to substantially expand the spatial coverage of F. radicans. Further, the lack of gas-filled bladders in F. radicans, that allow F. vesiculosus thalli to float for extensive time periods (up to 200 days), increasing its dispersal (Vandendriessche et al. 2006), diminishes the dispersal capacity of F. radicans compared to F. vesiculosus.
Despite the common occurrence of asexual reproduction in Fucus radicans in the Baltic Sea, it is yet unknown which, if any, environmental cues determine the success of this reproductive mode. The reproductive strategy and life cycles of fucoids growing in temperate regions are known to be affected by seasonality, mainly light and temperature, either as single or interacting factors (e.g. Berger et al. 2001, Kraufvelin et al. 2012). For example, thallus growth in Fucus vesiculosus is initiated at ca. 10°C in spring (Munda and Kremer 1977) and fertilized eggs of the same species require temperatures between ca. 8 and 14°C to attach to the substrate (Serrão et al. 1999). Further, rhizoid formation in Fucus sp. zygotes is light specific, needing wavelengths within the visible region (400–750 nm) in order to form (Whitaker 1942). However, the impact of seasonal cues such as daylength and temperature on asexual reproduction success in fucoids is still unknown. In this study we investigate experimentally whether the re-attachment of F. radicans fragments is affected by water temperature and light conditions (irradiance in combination with daylength). As metabolism and somatic growth in photosynthetic organisms increase when temperature and light levels approach optimal levels, we hypothesize that conditions with higher water temperature and higher irradiance together with long daylength will enhance the re-attachment success.
Materials and methods
Thalli of Fucus radicans Bergström et Kautsky (n=15; an individual was defined as a physically separate unit, irrespective of genotype) were collected during late July 2012 at Järnäs (63°26′6.6″N:19°40′8.6″E) and early August 2013 at Skagsudde (63°11′32.7″N:19°0′15.7″E) and Drivan (63°26′59.1″N:19°20′2.4″E). Previous samplings for genotyping at these sites revealed between 9 and 13 different genotypes for 50 sampled thalli within each site (Ardehed et al. 2015). To ensure that at least two genotypes were represented in the (non-genotyped) material, we collected both male and female thalli from all three donor sites for the experiment. The donor sites are covered with ice during winter (HELCOM 2013) and the surface water temperature during the summer months varies between 5 and 14°C (Serrão et al. 1999). The salinity at these sites ranges from 2.8 to 4.5 (Serrão et al. 1999, Forslund et al. 2012). The natural light levels during a sunny summer day at 2–4 m depth (where the Fucus stands are most dense) varies between 70 and 600 μmol photons m−2 s−1 (Serrão et al. 1999).
The collected thalli were transported within 24 h in cool, dark and moist conditions to Askö Laboratory (58°49′19.85″N, 17°38′8.46″E), where they were kept submerged in seawater flow through outdoor tanks (260 l h−1) with natural salinity of ca. 6 until the start of the experiment. As Fucus radicans grows in salinities of up to 6, this slight increase in salinity during the preliminary cultivation was not considered to cause extensive stress to the algae. One thallus was defined as several ramets (i.e. parts of thallus) growing from one holdfast. For transportation from Askö Laboratory to the experimental facilities at Roskilde University, Denmark, one or two ramets rich with adventitious branches were cut off from each thallus and placed in an individual plastic bag with moist tissue paper. The ramets were kept dark and cool during the transport (22 h). Fucoids are generally robust to drying, and they are able to recover rapidly after transportation in cool, moist and dark conditions (pers. obs.). Upon arrival the ramets were placed in individual beakers filled with water with salinity 5 and kept at +15°C in ca. 80 μmol photons m−2 s−1 for 3 days until the start of the experiment (Nygård and Dring 2008). At the beginning of the experiment ca. 100 fragments (length: 5–10 mm), were picked from each ramet and used in the experiment (see below).
The experiment was designed to mimic the average light and temperature conditions during the different seasons in the Bothnian Sea, i.e. summer (warm water temperature, high light), winter (cold water temperature, low light) and spring/autumn (cold water temperature, high light). The fourth treatment with warm water temperature and low light was necessary for balanced full factorial experimental design. Thus, the setup consisted of four treatment combinations with two levels of both factors light (high, low) and temperature (high, low). Light was provided by lamps with halogen spots (OSRAM Decostar 51:12V, 35W). The light regime was 16:8 (light:dark) for high light treatments and 6:18 for low light treatments. Further reduction of irradiance for the low light treatments was created by placing the light source higher up from the tank and placing fine black mesh over the aquaria. Irradiance levels for high and low light treatments were 96–100 and 30–33 μmol m−2 s−1, respectively (2π, quantitherm PAR/temperature sensor, Hansatech Instruments). These are equivalent to a clear summer day (Serrão et al. 1999) and under ice conditions, respectively (Tulonen et al. 1994) in the donor region. As we simultaneously manipulated both irradiance and daylength, the design did not allow us to estimate the individual impacts of these potentially confounding factors on Fucus radicans re-attachment. Stable target water temperatures for both high (+14°C) and low temperature treatments (+4°C) were obtained by placing aquaria in larger water baths (three aquaria in each bath) each equipped with a heater (Julabo ED, Julabo Laortechnik GmbH, Seelbach, Germany), a cooler and a water pump to maintain uniform temperature in the water bath (Figure 1). These temperatures correspond to the average summer and winter temperatures in the Bothnian Sea, that range from 8 to 15°C and from 1 to 4°C, respectively (HELCOM 2013). The higher range of each temperature interval was chosen as target temperatures in order to avoid freezing in the cold water treatment. Both irradiance and temperature were monitored with temperature and light loggers (HOBO Pendant®, 8K, Onset) throughout the experiment.

A schematic description of the experimental setting.
The experiment consisted of four such units equipped with a heater, a cooler and a water pump. Each jar (cylinder) represents a pseudoreplicate (n=8) for a true replicate (n=3). A tile with four fragments was placed in each jar (the small embedded figure).
An experimental unit consisted of transparent plastic jars with lid (V=0.45 l) and a white 4×4 cm ceramic tile with rough surface on the bottom as substrate. Each unit was filled with diluted seawater with a salinity of 5. The water was prepared by mixing seawater from the North Sea (salinity of 30) with non-chlorinated tap water with a naturally high dissolved inorganic carbon (DIC) concentration. The photosynthetic rate of Fucus in the Bothnian Sea increases with salinity (Nygård and Ekelund 2006). Therefore, the slightly higher salinity compared to the ambient salinity of the collected thalli, both before and during the experiment, should not negatively affect the physiological performance of the fragments. Ceramic tiles were chosen as similar rough, alkaline substrate has been observed to increase settling of Fucus vesiculosus germlings (Malm et al. 2003). The tiles were soaked for 24 h before the experiment. Four randomized Fucus radicans fragments from different thalli were placed on the tile in each experimental unit. No thallus was represented more than once in each unit. The units (n=96) were randomly placed in 12 aquaria (eight units in each) which were randomized for both light and temperature treatments (n=3 per treatment combination). The experiment was run for 7 weeks, which has been shown to be sufficient time for rhizoid formation and attachment for F. radicans fragments (Tatarenkov et al. 2005).
As re-attachment of fragments requires low water movement (Santelices 2004), and the natural circulation of water is reduced by ice cover in winter (Kelley 1997), we avoided excessive water movement in the experimental units. Low or absent water movement creates a thicker boundary layer around alga, which affects the nutrient uptake (Wheeler 1980). While previous studies have shown high survival and fitness of Fucus radicans fragments in still water (Tatarenkov et al. 2005), we analyzed the DIC concentrations in randomly selected experimental units from each treatment combination to control for sufficient carbon supply (~2.0 mol m−3; Bidwell and McLachlan 1985). Further, to minimize the risk for depletion of carbon and other nutrients over time, two thirds of the water in each unit was changed after 4 weeks. To avoid disturbing the re-attachment process, water was first carefully removed and then replaced by newly mixed aerated water by using a 5-ml pipette.
After 7 weeks, we counted the number and percentage of fragments (0, 25, 50, 75 or 100%) per experimental unit that had re-attached to the substrate. A fragment was defined as re-attached if it remained attached to the substrate after being gently rinsed with water.
Initially we analyzed the effects of temperature and light on re-attachment using univariate permutational nested analysis in PERMANOVA+ in PRIMER 6. Temperature and light were considered fixed factors and aquaria a random factor, nested under temperature and light treatments to adjust for the eight pseudoreplicates (i.e. non-independent experimental units) per aquaria. However, as aquaria had no significant effect on the re-attachment success (p0.05) and only explained a minor part (8.5%) of the variance, this nested factor was excluded from the final analysis. The final analysis thus consisted of a two-way univariate permutational analysis where we used the mean re-attachment values for each aquarium. Significant main factor interactions were followed by pairwise permutational post hoc comparisons (based on permutational t-statistics; Anderson et al. 2008). Resemblance matrix was based on Euclidean distance and the dispersion of data was inspected using PERMDISP. All analyses were conducted using 9999 permutations and with significance level of 0.05. Given values are mean±SE.
Results
Re-attachment experiment
The fragments attached mostly by rhizoids formed from the basal parts, while a few fragments with rhizoids failed to attach to the substratum. In addition, many fragments had formed long, thick bunches of cryptostomata hairs. Further, we observed that somatic growth in terms of produced side shoots had begun on most fragments. These fragments had formed new shoots, often several, from the basal part. The formation of these side shoots also took place in fragments showing no rhizoid formation.
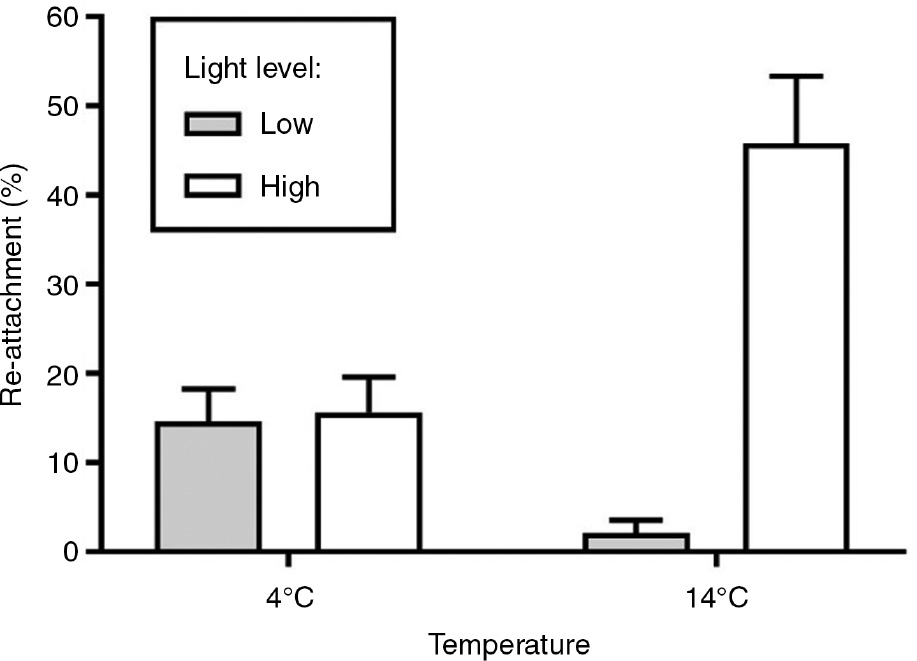
The average re-attachment percentage of fragments was 19.5% across all treatments. Temperature and light had an interactive impact on re-attachment (Table 1). At low temperatures, light had no effect on re-attachment (pairwise post hoc comparison, p>0.05, Figure 2) and the two light treatments showed similar re-attachment success. At high temperatures, however, light treatment affected re-attachment success significantly with higher re-attachment in high light compared to low light condition (pairwise post hoc comparison, p=0.0059).
Fucus radicans response to light and temperature treatment.
| Factor | df | MS | Pseudo-F | p-Value |
|---|---|---|---|---|
| Temperature | 1 | 0.3763 | 2.470 | 0.1519 |
| Light | 1 | 2.4076 | 15.803 | 0.0042 |
| Temperature×light | 1 | 2.1888 | 14.368 | 0.0045 |
| Residual | 8 | 0.1523 |
Results from univariate PERMANOVA using mean values from each aquarium. p-Values in bold indicate significant results at α=0.05.
Fucus radicans.
Percentage (±standard error) of re-attached fragments after 7 weeks cultivation in four different treatment combinations. The x-axis indicates the temperature treatment (4°C or 14°C) while the gray and white bars indicate the low (ca. 33 μmol m−2 s−1, 6 h:18 h light:dark regime) and high light level (ca. 100 μmol m−2 s−1, 16 h:8 h light:dark regime) treatments, respectively. n=3 (with eight pseudoreplicates each) for each treatment combination.
Discussion
Understanding which factors control re-attachment is not only important for mechanistic understanding of the reproductive biology of Fucus radicans. It is also necessary when estimating the population dynamics of this habitat forming foundation species. In this study we determined how two important environmental factors, temperature and light affect re-settling success of asexual F. radicans fragments. The fragments were able to re-attach in all the tested conditions, but the re-attachment was interactively affected by temperature and light. The re-attachment success was much higher in the conditions corresponding to summer (high temperature and high light level) compared to all the other treatment combinations. Re-attachment success was equal in conditions mimicking winter (low temperature and low light level) and spring or autumn (low temperature and high light level). High water temperature under low light conditions resulted in relatively poor re-attachment success.
Fragment re-attachment and growth
By the end of the experiment, most fragments had attached by rhizoids, as expected, and many of the re-attached fragments had also formed long, thick bunches of cryptostomata hairs. The exact function of these has not been established, but they have been suggested to enhance nutrient uptake (Chapman 1995 and references therein), as with euryhaline hairs in Ceramium sp. (DeBoer and Whoriskey 1983). The rich abundance of cryptostomata hairs might thus assist the growth of the small fragment by facilitating nutrient uptake, in a similar way to apical hairs found on sexually produced germlings (Hurd et al. 1993). The production of new side shoots from the basal part of the fragments was common in the current study, irrespective of rhizoid formation. Similar basal proliferations have been shown to occur on Fucus vesiculosus fragments as a response to wounding (Moss 1961). If Fucus radicans fragments fail to reattach successfully, they might be able to form benthopleustophytic populations, as are documented for F. vesiculosus in the Baltic Proper (Svedelius 1901, Waern 1952).
Impact of environmental conditions
The re-attachment pattern in different light and temperature combinations suggests that, instead of a specific light or temperature threshold, the re-attachment success may depend simply on the net primary production (metabolic balance), and is enhanced in conditions favorable for positive net primary production. Both temperature and light are key factors affecting a multitude of processes in photosynthetic organisms. As somatic growth, including rhizoid formation, is an energy requiring process, re-settlement was, as we hypothesized, enhanced in conditions favorable for growth in photosynthetic organisms. Organismal metabolism increases with temperature in ectothermic organisms (Brown et al. 2004) simultaneously increasing the light requirements in photosynthetic organisms. In light-saturated settings, higher temperature may lead to more efficient photosynthesis (Davison et al. 1991). Accordingly, the re-attachment success of fragments was highest when exposed to both high light and high temperature. Due to the higher compensation point for photosynthesis in high temperatures compared to lower temperatures, limited light availability will reduce the net primary production (Davison et al. 1991). For photosynthetic organisms, low irradiance levels may thus not be sufficient to produce enough energy to keep up with metabolism in high temperatures, leading to decreased functionality, growth or survival. Consequently, the re-attachment success of fragments was much lower in the experimental treatment with high temperature and low light levels compared to all other treatment combinations, potentially due to energy depleting conditions (i.e. high metabolism and low photosynthetic rates). Similarly, rhizoid formation in the green filamentous algae Spirogyra has been observed to be light dependent and optimal in temperatures of 20°C (Nagata 1973a,b). Further, the lack of impact of light level on re-attachment success in the low temperature conditions suggests that low light levels under ice are likely high enough or even saturating for the lowered metabolism in Fucus radicans fragments during winter. Accordingly, the optimal rate of photosynthesis for Fucus in the Bothnian Sea is lower (4–10°C) in low salinities (salinity of 5), compared to more central (the Irish Sea) populations, that reach maximum photosynthetic rate in higher temperatures (ca. 15–20°C; Nygård and Dring 2008).
Benthic environments are complex systems, with a multitude of physical and biological mechanisms determining the success of different species. Besides factors affecting photosynthesis, other factors such as hydrodynamics and biotic interactions have been suggested to contribute to the re-attachment success of vegetative fragments. For example, Bergström et al. (2005) speculated that re-attachment might be promoted by long periods of calm water, such as conditions under the ice, since the fragments are easily re-suspended. In our experiment, the fragments were able to re-attach in all tested conditions, which should be advantageous for the individual fitness as, at least theoretically, this plasticity can provide a temporal escape from competition or too harsh environmental settings (i.e. low salinity). In addition to the flexibility in timing of asexual reproduction, the larger body size of asexual fragments might be beneficial in stressful environments compared to the smaller sexual life stages. Fragments may be able to withstand sedimentation better than small germlings, as larger organisms are not equally easily buried under sediment. Relatively large body size might also decrease the vulnerability to grazing. Gastropods have been shown to graze on Fucus germlings up to 1 mm, affecting the post-settlement mortality (Malm et al. 1999). However, whether and how potential seasonal refuge from stress and the larger size of asexual life stages compared to the asexual life stages contributes to individual fitness requires further investigation.
Conclusions
Estimating the relative importance, mechanisms and potential of vegetative reproduction compared to sexual reproduction is required for the understanding of the population and dispersal dynamics of Fucus radicans. We here made the first effort to shed light on the mechanisms determining how environmental settings affect the success of vegetative reproduction in this endemic foundation species. Out results suggest that, more than light or temperature cues per se, light and temperature also affect re-settling of asexual fragments interactively. We suggest a potential mechanism for reattachment through a combination of enhanced metabolic activity and positive net primary production.
About the authors

Ellen Schagerström has a PhD in Plant Ecology and is a researcher at Stockholm University, Department of Ecology, Environment and Plant Science and at the Baltic Sea Centre. Her research expertise is within the field of marine ecology focusing on biodiversity both at the genetic and species level in Fucus communities as well as adaptation and speciation. Furthermore, her research covers studies of dynamics of benthic vegetation as well as restoration of shallow benthic communities.

Tiina Salo works on understanding ecological and eco-evolutionary effects of stress on individuals, populations and communities. She uses macroalgae, seagrasses and invertebrates as her model organisms to study organismal and community traits. She is currently a post-doctoral fellow at Stockholm University, Sweden.
Acknowledgements
The authors are grateful to M.F. Pedersen for comments on the experimental design and L. Kautsky for comments on the manuscript. Data from the Swedish environmental monitoring and the database dBotnia at Umeå Marine Sciences Centre was used to estimate seasonal average temperature in the Bothnian Sea. This study was funded by C.F Lundström Foundation (ES), Stockholm University Baltic Sea Centre (ES) and Roskilde University (TS). We also thank two anonymous reviewers who helped improve the manuscript.
References
Andersson, S., L. Kautsky and A. Kalvas. 1994. Circadian and lunar gamete release in Fucus vesiculosus in the atidal Baltic Sea. Mar. Ecol. Prog. Ser. 110: 195–201.10.3354/meps110195Search in Google Scholar
Anderson, M.J., R.N. Gorley and K.R. Clarke. 2008. PERMANOVA+ for PRINMER: Guide to software and statistical methods. PRIMER-E. Plymouth, UK.Search in Google Scholar
Ardehed, A., D. Johansson, E. Schagerström, L. Kautsky, K. Johannesson and R.T. Pereyra. 2015. Complex spatial clonal structure in the macroalgae Fucus radicans with both sexual and asexual recruitment. Ecol. Evol. 5: 4233–4245.10.1002/ece3.1629Search in Google Scholar
Berger, R., T. Malm and L. Kautsky. 2001. Two reproductive strategies in Baltic Fucus vesiculosus (Phaeophyceae). Eur. J. Phycol. 36: 265–273.10.1080/09670260110001735418Search in Google Scholar
Bergström, L., E. Bruno, B. Eklund and L. Kautsky. 2003. Reproductive strategies of Ceramium tenuicorne near its inner limit in the brackish Baltic Sea. Bot. Mar. 46: 125–131.10.1515/BOT.2003.013Search in Google Scholar
Bergström, L., A. Tatarenkov, K. Johannesson, R.B. Jonsson and L. Kautsky. 2005. Genetic and morphological identification of Fucus radicans sp Nov (Fucales, Phaeophyceae) in the brackish Baltic Sea. J. Phycol. 41: 1025–1038.10.1111/j.1529-8817.2005.00125.xSearch in Google Scholar
Bidwell, R. and J. McLachlan. 1985. Carbon nutrition of seaweeds: photosynthesis, photorespiration and respiration. J. Exp. Mar. Biol. Ecol. 86: 15–46.10.1016/0022-0981(85)90040-1Search in Google Scholar
Brown, J.F., J.F. Gillooly, A.P. Allen, V.M. Savage and G.B. West. 2004. Toward metabolic theory of ecology. Ecology 85: 1771–1789.10.1890/03-9000Search in Google Scholar
Ceccherelli, G. and L. Piazzi. 2001. Dispersal of Caulerpa racemosa fragments in the Mediterranean: lack of detachment time effect on establishment. Bot. Mar. Book 44: 209–213.10.1515/BOT.2001.027Search in Google Scholar
Chapman, A. 1995. Functional ecology of fucoid algae: twenty-three years of progress. Phycologia 34: 1–32.10.2216/i0031-8884-34-1-1.1Search in Google Scholar
Collado-Vides, L. 2001. Clonal architecture in marine macroalgae: ecological and evolutionary perspectives. Evol. Ecol. 15: 531–545.10.1007/978-94-017-1345-0_17Search in Google Scholar
Davison, I.R., R.M. Greene and E.J. Podolak. 1991. Temperature acclimation of respiration and photosynthesis in the brown alga Laminaria saccharina. Mar. Biol. 110: 449–454.10.1007/BF01344363Search in Google Scholar
Dayton, P.K. 1971. Toward an understanding of community resilience and the potential effects of enrichments to the benthos at McMurdo sound, Antarctica. In: (B.C. Parker, ed) Proc Conservation problems in Antarctica. Allen Press Imc., Lawrence, Kansas.Search in Google Scholar
DeBoer, J.A. and F.G. Whoriskey. 1983. Production and role of hyaline hairs in Ceramium rubrum. Mar. Biol. 77: 229–234.10.1007/BF00395811Search in Google Scholar
Feistel, R., S. Weinreben, H. Wolf, S. Seitz, P. Spitzer, B. Adel, G. Nausch, B. Schneider and D. Wright. 2010. Density and absolute salinity of the Baltic Sea 2006–2009. Ocean Sci. 6: 3–24.10.5194/os-6-3-2010Search in Google Scholar
Forslund, H. and L. Kautsky. 2013. Reproduction and reproductive isolation in Fucus radicans (Phaeophyceae). Mar. Biol. Res. 9: 321–326.10.1080/17451000.2012.731694Search in Google Scholar
Forslund, H., O. Eriksson and L. Kautsky. 2012. Grazing and geographic range of the Baltic seaweed Fucus radicans (Phaeophyceae). Mar. Biol. Res. 8: 386–394.10.1080/17451000.2011.637565Search in Google Scholar
Gagnon, K., C. McKindsey and L. Johnson. 2014. Roles of dispersal mode, recipient environment and disturbance in the secondary spread of the invasive seaweed Codium fragile. Biol. Invasions. 17: 1–14.10.1007/s10530-014-0782-2Search in Google Scholar
Gylle, A.M., S. Rantamäki, N.G.A. Ekelund and E. Tyystjärvi. 2011. Fluorescence emission spectra of marine and brackish-water ecotypes of Fucus vesiculosus and Fucus radicans (Phaeophyceae) reveal differences in light harvesting apparatus. J. Phycol. 47: 98–105.10.1111/j.1529-8817.2010.00928.xSearch in Google Scholar PubMed
HELCOM. 2013. Climate change in the Baltic Sea Area: HELCOM thematic assessment in 2013. Baltic Sea Environment Proceedings, No. 143.Search in Google Scholar
Hurd, C.L., R.S. Galvin, T.A. Norton and M.J. Dring. 1993. Production of hyaline hairs by intertidal species of Fucus (Fucales) and their role in phosphate uptake. J. Phycol. 29: 160–165.10.1111/j.0022-3646.1993.00160.xSearch in Google Scholar
Johannesson, K. and C. Andre. 2006. Life on the margin: genetic isolation and diversity loss in a peripheral marine ecosystem, the Baltic Sea. Mol. Ecol. 15: 2013–2029.10.1111/j.1365-294X.2006.02919.xSearch in Google Scholar PubMed
Johannesson, K., D. Johansson, K.H. Larsson, C.J. Huenchuñir, J. Perus, H. Forslund, L. Kautsky and R.T. Pereyra. 2011. Frequent clonality in fucoids (Fucus radicans and Fucus vesiculosus; Fucales, Phaeophyceae) in the Baltic Sea. J. Phycol. 47: 990–998.10.1111/j.1529-8817.2011.01032.xSearch in Google Scholar PubMed
Kelley, D.E. 1997. Convection in ice-covered lakes: effects on algal suspension. J. Plankton Res. 19: 1859–1880.10.1093/plankt/19.12.1859Search in Google Scholar
Klimeš, L., J. Klimešová, R. Hendriks and J. van Groenendael. 1997. Clonal plant architecture: a comparative analysis of form and function. In: (H. de Kroon and J. van Groenendael, eds) The ecology and evolution of clonal plants. Backhuys Publishers, Leiden.Search in Google Scholar
Kostamo, K. 2008. The life cycle and genetic structure of the red alga Furcellaria lumbricalis on a salinity gradient. W. and A. De Nottbeck Foundation Sci. Rep. 33: 1–34.Search in Google Scholar
Kraufvelin, P., A.T. Ruuskanen, S. Bäck and G. Russell. 2012. Increased seawater temperature and light during early springs accelerate receptacle growth of Fucus vesiculosus in the northern Baltic proper. Mar. Biol. 159: 1795–1807.10.1007/s00227-012-1970-1Search in Google Scholar
Malm, T., R. Engkvist and L. Kautsky. 1999. Grazing effects of two freshwater snails on juvenile Fucus vesiculosus in the Baltic Sea. Mar. Ecol. Prog. Ser. 188: 63–71.10.3354/meps188063Search in Google Scholar
Malm, T., L. Kautsky and T. Claesson. 2003. The density and survival of Fucus vesiculosus L. (Fucales, Phaeophyta) on different bedrock types on a Baltic Sea moraine coast. Bot. Mar. 46: 256–262.10.1515/BOT.2003.023Search in Google Scholar
Moss, B.L. 1961. Wound healing and regeneration in Fucus vesiculosus L. 4th International Seaweed Symposium, Biarritz.Search in Google Scholar
Munda, I.-M. and B. Kremer. 1977. Chemical composition and physiological properties of fucoids under conditions of reduced salinity. Mar. Biol. 42: 9–15.10.1007/BF00392009Search in Google Scholar
Nagata, Y. 1973a. Rhizoid differentiation in Spirogyra I. Basic features of rhizoid formation. Plant cell. Physiol. 14: 531–541.Search in Google Scholar
Nagata, Y. 1973b. Rhizoid differentiation in Spirogyra II. Photoreversibility of rhizoid induction by red and far-red light. Plant Cell. Physiol. 14: 543–554.Search in Google Scholar
Nygård, C.A. and M.J. Dring. 2008. Influence of salinity, temperature, dissolved inorganic carbon and nutrient concentration on the photosynthesis and growth of Fucus vesiculosus from the Baltic and Irish Seas. Eur. J. Phycol. 43: 253–262.10.1080/09670260802172627Search in Google Scholar
Nygård, C.A. and N. Ekelund. 2006. Photosynthesis and UV-B tolerance of the marine alga Fucus vesiculosus at different sea water salinities. Eighteenth International Seaweed Symposium. Springer. pp. 235–241.10.1007/978-1-4020-5670-3_29Search in Google Scholar
Pearson, G., L. Kautsky and E. Serrão. 2000. Recent evolution in Baltic Fucus vesiculosus: reduced tolerance to emersion stresses compared to intertidal (North Sea) populations. Mar. Ecol. Prog. Ser. 202: 67–79.10.3354/meps202067Search in Google Scholar
Pereyra, R.T., L. Bergström, L. Kautsky and K. Johannesson. 2009. Rapid speciation of a brown alga in the Baltic Sea. Phycologia 48: 301.Search in Google Scholar
Raven, J.A. and G. Samuelsson. 1988. Ecophysiology of Fucus vesiculosus L. Close to Its Northern Limit in the Gulf of Bothnia. Bot Mar. 31: 399–410.10.1515/botm.1988.31.5.399Search in Google Scholar
Santelices, B. 2004. A comparison of ecological responses among aclonal (unitary), clonal and coalescing macroalgae. J. Exp. Mar. Biol. Ecol. 300: 31–64.10.1016/j.jembe.2003.12.017Search in Google Scholar
Santelices, B. and D. Varela. 1993. Intra-clonal variation in the red seaweed Gracilaria chilensis. Mar. Biol. 116: 543–552.10.1007/BF00355472Search in Google Scholar
Santelices, B., D. Aedo and D. Varela. 1995. Causes and implications of intra-clonal variation in Gracilaria chilensis (Rhodophyta). J. Appl. Phycol. 7: 283–290.10.1007/BF00004002Search in Google Scholar
Serrão, E.A., L. Kautsky and S.H. Brawley. 1996. Distributional success of the marine seaweed Fucus vesiculosus L in the brackish Baltic Sea correlates with osmotic capabilities of Baltic gametes. Oecologia 107: 1–12.10.1007/BF00582229Search in Google Scholar
Serrão, E.A., L. Kautsky, T. Lifvergren and S.H. Brawley. 1997. Gamete dispersal and pre-recruitment mortality in Baltic Fucus vesiculosus. Phycologia 36: 101–102.Search in Google Scholar
Serrão, E.A., S.H. Brawley, J. Hedman, L. Kautsky and G. Samuelson. 1999. Reproductive success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. J. Phycol. 35: 254–269.10.1046/j.1529-8817.1999.3520254.xSearch in Google Scholar
Svedelius, N. 1901. Studier Öfver Östersjöns Hafsalgflora. Uppsala Nya Tidnings Aktiebolag, Uppsala.10.5962/bhl.title.63707Search in Google Scholar
Tatarenkov, A., L. Bergström, R.B. Jonsson, E.A. Serrão, L. Kautsky and K. Johannesson. 2005. Intriguing asexual life in marginal populations of the brown seaweed Fucus vesiculosus. Mol. Ecol. 14: 647–651.10.1111/j.1365-294X.2005.02425.xSearch in Google Scholar
Tulonen, T., P. Kankaala, A. Ojala and L. Arvola. 1994. Factors controlling production of phytoplankton and bacteria under ice in a humic, boreal lake. J. Plankton Res. 16: 1411–1432.10.1093/plankt/16.10.1411Search in Google Scholar
Vandendriessche, S., M. Vincx and S. Degraer. 2006. Floating seaweed in the neustonic environment: a case study from Belgian coastal waters. J. Sea Res. 55: 103–112.10.1016/j.seares.2005.09.002Search in Google Scholar
Waern, M. 1952. Rocky-shore algae in the Öregrund archipelago. PhD, Uppsala University, Uppsala.Search in Google Scholar
Walters, L.J., C.M. Smith, J.A. Coyer, C.L. Hunter, K.S. Beach and P.S. Vroom. 2002. Asexual propagation in the coral reef macroalga Halimeda (Chlorophyta, Bryopsidales): production, dispersal and attachment of small fragments. J. Exp. Mar. Biol. Ecol. 278: 47–65.10.1016/S0022-0981(02)00335-0Search in Google Scholar
Wheeler, W. 1980. Effect of boundary layer transport on the fixation of carbon by the giant kelp Macrocystis pyrifera. Mar. Biol. 56: 103–110.10.1007/BF00397128Search in Google Scholar
Whitaker, D. 1942. Counteracting the retarding and inhibitory effects of strong ultraviolet on Fucus eggs by white light. J. Gen. Physiol. 25: 391–397.10.1085/jgp.25.3.391Search in Google Scholar PubMed PubMed Central
©2019 Ellen Schagerström et al., published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 License.
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Baltic algae
- Physiology and ecology
- Limited response of a spring bloom community inoculated with filamentous cyanobacteria to elevated temperature and pCO2
- Fucus vesiculosus adapted to a life in the Baltic Sea: impacts on recruitment, growth, re-establishment and restoration
- Acclimation limits of Fucus evanescens along the salinity gradient of the southwestern Baltic Sea
- Interactive effects of temperature and light on reattachment success in the brown alga Fucus radicans
- Phylogeny and biogeography
- Surveying seaweeds from the Ulvales and Fucales in the world’s most frequently used artificial waterway, the Kiel Canal
- New records from the southern North Sea and first records from the Baltic Sea of Kornmannia leptoderma
- Genetic variability of charophyte algae in the Baltic Sea area
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Baltic algae
- Physiology and ecology
- Limited response of a spring bloom community inoculated with filamentous cyanobacteria to elevated temperature and pCO2
- Fucus vesiculosus adapted to a life in the Baltic Sea: impacts on recruitment, growth, re-establishment and restoration
- Acclimation limits of Fucus evanescens along the salinity gradient of the southwestern Baltic Sea
- Interactive effects of temperature and light on reattachment success in the brown alga Fucus radicans
- Phylogeny and biogeography
- Surveying seaweeds from the Ulvales and Fucales in the world’s most frequently used artificial waterway, the Kiel Canal
- New records from the southern North Sea and first records from the Baltic Sea of Kornmannia leptoderma
- Genetic variability of charophyte algae in the Baltic Sea area