Expanding known dinoflagellate distributions: investigations of slurry cultures from Caspian Sea sediment
-
Jane Lewis
Jane Lewis is a Professor in the Department of Life Sciences at the University of Westminster. Her research is focussed on the ecology and taxonomy of dinoflagellates. There have been two main themes for this research – the use of dinoflagellate cysts to interpret the fossil record and the role of microalgal life cycles in harmful algal blooms. Current research includes the role of temperature in modulating life cycle transitions in
Alexandrium tamarense in the Gulf of Maine and the morphological variation of cysts in various species within the Gonyaulacales. ,
Joe D. Taylor
,
Joe D. Taylor
Joe D. Taylor is an independent research fellow at the University of Salford, UK. He was awarded a PhD in Environmental Microbiology by the University of Essex, UK, working on microbial degradation of algal polysaccharides in estuarine littoral zones. He has worked on across multiple habitats and with multiple taxonomy groups. His current research is focused on micro-eukaryote diversity and function in oceanic, coastal and terrestrial food-webs.
Suzanne A.G. Leroy was Professor at Brunel University London (UK) from 2000 to 2016. She is now an associate researcher at the CEREGE, France. She is a palynologist, specialising in the Caspian Sea palaeo-environmental history since 1994. She has pioneered the work on dinocysts from Caspian sediment and has stimulated their taxonomical and phylogenetical analyses. Other areas of current research include palaeo-climates and lake-level changes in the Middle East and Central Asia.

Abstract
To investigate the disparity between plankton and cyst records, sediment slurry cultures were used to isolate the motile stage of dinoflagellates from Caspian Sea sediment. This has resulted in new records for this area of Kryptoperidinium foliaceum, Gymnodinium aureolum and Woloszynskia sp. and for the cyst record, Scrippsiella acuminata. Two Gonyaulax species were isolated, one was identified as Gonyaulax baltica and the other an unknown species. Cultures of Lingulodinium polyedra were also isolated. The approach of using slurries was useful to provide cultures from sediments that were relatively poor in dinoflagellate cysts with contents.
Introduction
The Caspian Sea is the largest inland body of water in the world with a surface area of 3.5 million km2. It is a complex lake system with three distinct basins, of which the southern one is the most saline (13), the warmest (surface waters 10–28°C) and the deepest (1025 m) (Kosarev and Yablonskaya 1994). The hydrography of the lake is driven by riverine inputs (largely in the north) and seasonal climatic changes over its long latitudinal range (36°33′–47°07′N). The biota of the lake is dominated by freshwater/brackish species and is characterised by a large amount of endemism (Dumont 1998). Particular ecological pressures have been experienced in the lake system from pollution, oil and water exploitation and also, more recently, introduced species through shipping and the development of the Volga-Don canal. A recent driver of change in the plankton community has come from the introduced ctenophore Mnemiopsis leidyi A. Agassiz (Kideys et al. 2008, Roohi et al. 2010, Nasrollahzadeh et al. 2014). Phytoplankton communities have changed over the long term with fluctuation of freshwater input, changes in nutrient inputs and introduction of new species (Kosarev and Yablonskaya 1994). From 1994 to 2005, the productivity of the southern basin changed from oligotrophic to meso-eutrophic (Nasrollahzadeh et al. 2008a,b, Bagheri et al. 2011). Phytoplankton diversity for the Caspian is dominated by cyanophytes (147 taxa) and diatoms (238 taxa) (Gogorev 2006). However, with respect to dinoflagellates, it is notable that in the various studies of the phytoplankton flora of the Caspian most papers list only some 20–30 taxa (Table 1 and references therein). The total number of species recorded is just under 50 contrasting with a checklist for the Black Sea comprising 267 species (Gómez and Boicenco 2004) and for the Mediterranean comprising 673 species (Gómez 2003). Although low in species diversity, dinoflagellates are an important component of the phytoplankton community in terms of abundance and biomass (Kideys et al. 2005, Bagheri et al. 2012b, Nasrollahzadeh et al. 2014) with Prorocentrum Ehrenberg species being the most abundant (Kosarev and Yablonskaya 1994, Kideys et al. 2005). Blooms of Prorocentrum cordatum (Ostenfeld) Dodge (Bagheri and Fallahi 2014) and Heterocapsa F. Stein (Bagheri et al. 2012a) have been recorded and Lingulodinium polyedra (Stein) Dodge contributed to a bloom dominated by Nodularia Mertens ex Bordet et Flahault in 2009 (Nasrollahzadeh et al. 2011).
Brackish/marine dinoflagellatesa recorded in the Caspian Sea in the phytoplankton and in recent sediments (linkages between the two are recorded where known).
| Phytoplanktonb | Sediments |
|---|---|
| Amphidinium cf. rhynchocephalum Annisomova1 | |
| Diplopsalis acuta Paulsen2,3,7,9 | |
| D. lenticula Bergh7,8,9 | |
| Exuviaella cordata Ostenfeld2 | |
| Glenodinium behningii (Lindemann) Kisselev2,3,5,6,7,9 | |
| G. caspicum (Ostenfeld) Schiller3,7,8,9 | |
| G. obliquum (Pouchet)1 | |
| G. paululum Lindemann1 | |
| G. penardii Lemmermann2,9 | |
| G. pilula (Ostenfeld) Schiller9 | |
| Gonyaulax baltica Ellegaard, Lewis et Harding15 | Impagidinium caspienense Marret10,11,12,15 |
| G. digitale Kofoid2,7,8,9 | |
| G. minima Matzenauer2 | |
| G. polygramma Stein1,4 | |
| G. spinifera Diesing2,7,8,9 | |
| Gonyostomum depressum Herdman2 | |
| G. semen Diesing7,8 | |
| Gymnodinium fuscum (Ehrenberg) Stein1 | |
| G. lacustre Schiller7,8,9 | |
| G. sanguineum Hirasaka1 | |
| G. variabile Herdman2,5,6,7,8,9 | |
| G. wulfii Schiller1 | |
| Gyrodinium fusiforme Kofoid et Swezy1 | |
| G. pingue (Schütt) Kofoid et Swezy1 | |
| Heteraulacus polyedricus (Pouchet) Drugg et Loeblich6 | |
| Heterocapsa rotundata (Lohmann) Hansen9 | |
| H. triquetra (Ehrenberg) Stein6 | |
| Kolkwitziella acuta (Apstein) Elbrächter14 | Cysts of Kolkwitziella acuta14 |
| Lingulodinium polyedra (Stein) Dodge2,6,7,8,9 | Lingulodinium machaerophorum (Deflandre et Cookson) Wall10,11,12 |
| Oblea rotunda (Lebour) Balech ex Sournia1,9 | |
| Peridiniella danica (Paulsen) Okolodkov et Dodge2,7,8,9 | |
| Peridiniopsis berolinense9 | |
| Peridinium minusculum Pavillard1 | |
| P. subsalum Ostenfeld2 | |
| Preperidinium meunieri (Pavillard) Elbrächter2,5,9 | |
| Prorocentrum compressum (Bailey) Abé ex Dodge1 | |
| P. cordatum (Ostenfeld) Dodge2,3,4,5,6,7,8,9 | |
| P. lima (Ehrenberg) Stein9 | |
| P. micans Ehrenberg1,2,7,9 | |
| P. obtusum Ostenfeld2,7,8,9 | |
| P. proximum Makarova2,5,6,9 | |
| P. scutellum Schröder1,2, 3,5,6,7,8,9 | |
| Protoperidinium achromaticum Levander3,6,7,9 | |
| P. granii Balech7 | |
| P. leonis Pavillard7,8 | |
| P. pallidum Balech7 | |
| Pyrophacus horologicum Stein9 | |
| Scrippsiella trochoidea (Stein) Loeblich1,2,6,9 | |
| Caspidinium rugosum Marret10,11,12 | |
| Pyxidiniopsis psilata (Wall et Dale) Head10,15 | |
| Spiniferites belerius Reid11 | |
| Spiniferites cruciformis Wall et Dale10,11,12 | |
| Cysts of Pentapharsodinium dalei Indelicato et Loeblich III10,11,12 | |
| Cysts of Scrippsiella plana Luo, Mertens, Bagheri et Gu13 | |
| Brigantedinium Reid10,11,12 |
aGogorev (2006) includes a further 14 freshwater species found in the north of the Caspian Sea.
bSpecies names updated according to AlgaeBase (Guiry and Guiry 2017).
1Kideys et al. (2005); 2Ganjian et al. (2010); 3Bagheri et al. (2012b); 4Pautova et al. (2015); 5Nasrollahzadeh et al. (2008a); 6Nasrollahzadeh et al. (2014); 7Bagheri and Fallahi (2014); 8Bagheri et al. (2010); 9Gogorev (2006); 10Marret et al. (2004); 11Leroy et al. (2013c); 12Kazancı et al. (2004); 13Luo et al. (2016); 14Mertens et al. (2015a,b); 15Mertens et al. (2017).
The sediment of the Caspian Sea has been the focus of a number of recent palynological studies investigating the history and geography of the basin (Leroy et al. 2006, 2007, 2011, 2013a,b,c, 2014), with descriptions and taxonomy of recent dinoflagellate cysts in Marret et al. (2004), Leroy et al. (2006) and Leroy (2010). Again the relative paucity of species is of note. A comparison of the literature from the plankton and sediment (Table 1) reveals that, with the exception of Gonyaulax baltica Ellegaard, Lewis et Harding, L. polyedra and Scrippsiella plana Luo, Mertens, Bagheri et Gu, little congruity exists between the two lists. This study was undertaken to explore this disparity and this paper documents preliminary investigations of southern Caspian Sea sediment designed to test the diversity of living dinoflagellates held in the sedimentary record.
Materials and methods
Grab samples were collected from a motorboat and the top 1-cm sediment was removed and placed in airtight, dark containers and kept in the cool and dark until processing. Stations 1–7 were taken along the Gorgan transect on 9 February 2014 with station 1 nearest to the shore (2 m deep) and station 7 furthest from the shore (13 m deep). Sample HCGA09 was taken 1.8 m depth in the Anzali Lagoon at 37°26′56.6N and 49°22′49.8E on 26 June 2008 (Figure 1).
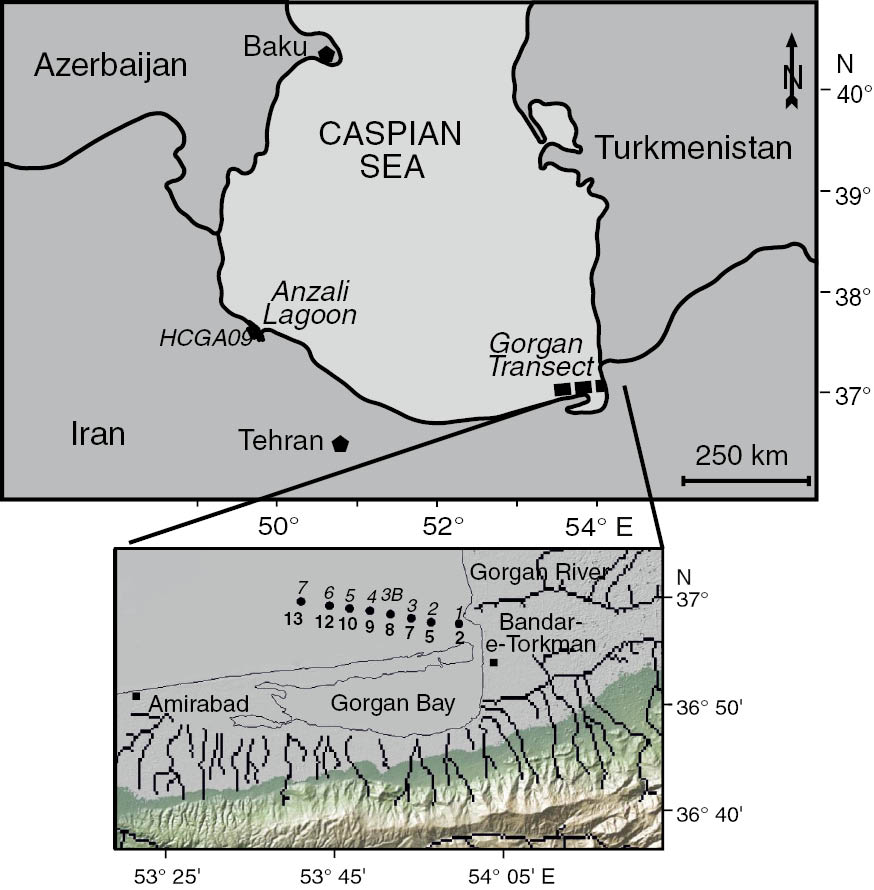
Location of grab samples in the Caspian Sea.
In the Gorgan transect, the station numbers are in italics and the water depths (m) in bold.
Initial work revealed that, apart from Lingulodinium polyedra, very low number of cysts with contents were found in the Caspian Sea sediment samples. Therefore, the approach of using slurry cultures to establish viability and diversity was initiated. For comparison, a single sediment sample from the Black Sea was also treated in the same way. Slurry cultures were established with approximately 1 cm3 wet sediment from each sample (Caspian Sea stations 1–7 and HCGA09, and Black Sea sample). These were briefly sonicated (ca. 2 min), sieved through a 80- and 20-μm mesh and the material retained on the 20-μm mesh was processed using the sodium polytungstate density gradient method as described in Bolch (1997). The recovered cysts were incubated as slurry cultures at 10°C, under fluorescent tubes, ~160 μmol photons m−2 s−1, 14:10 h light:dark cycle in 30-mm sterile Petri dishes with ca. 4 ml modified f/20 or f/2 medium without silicate (Guillard 1973). Medium was modified by the addition of sodium selenite (Na2SeO3, Sigma, St. Louis, MI, USA, final concentration 10−8m) and reducing copper sulfate (CuSO4·5H2O, BDH, Poole, UK, final concentration 10−8m) and prepared with 0.2-μm filtered natural seawater and distilled water – final salinity ca. 12). Single cells were isolated from the slurries using 96-well plates with 200 μl of modified f/20 or f/2 and incubated at 15°C, under fluorescent tubes, ~160 μmol photons m−2 s−1, 14:10 h light:dark cycle. Successful isolations were progressively scaled up through small Petri dishes to f/2 in 25-ml tissue culture flasks.
Cultures were observed on an inverted (Olympus IMT-2, Olympus, London, UK) or a dissecting microscope (Olympus SZH-ILLK, Olympus, London, UK). Detailed light microscopy of cells was carried out with an Olympus BH2 microscope (Olympus, London, UK) and photographed using a Zeiss Axioskop 2 microscope (Carl Zeiss Ltd., Cambridge, UK) with a Leica DFC290HS camera (Leica, London, UK) or a Nikon Eclipse Ci-L microscope, Nikon, Tokyo, Japan, fitted with a Nikon digital sight DS-Fi2 camera (Nikon, Tokyo, Japan) using NIS elements software (Nikon, Tokyo, Japan). Cell dissections were facilitated by the use of 2% sodium hypochlorite (Sigma-Aldrich, Gillingham, UK) and the plates were stained with Trypan blue (Sigma-Aldrich, Gillingham, UK) and examined under phase contrast optics. Dodge (1982), Steidinger (1997) and Hoppenrath et al. (2009) were used for initial identifications with follow up in primary literature as indicated in the species descriptions below.
For molecular work, cultures were harvested in the exponential growth phase. Approximately 15 ml of culture was centrifuged at 1537 g for 15 min. The supernatant was removed and the Deoxyribonuleic acid (DNA) was extracted using an Invisorb® Spin Plant Mini Kit (Invisorb, Berlin, Germany), where 0.4 ml of lysis buffer from the kit was used to re-suspend the cell pellet and the lysis buffer was added to screw cap micro-centrifuge tubes containing 0.2 g glass beads (600 μm), the tubes were placed in a BioSpec 3110BX Mini-BeadBeater-1 (BioSpec, Bartlesville, USA) and the machine was run for 60 s at 4800 oscillations min−1 to disrupt the cell membranes and lyse the DNA into solution. The remaining protocol was carried out in accordance with the manufacturer’s instructions and the DNA was re-suspended in 50 μl nuclease free water (Ambion, Thermo Fisher Scientific, Waltham, MA, USA). The primers D1R (5′ACCCGCTGAATTTAAGCATA 3′; Lenaers et al. 1989) and DC3Ca (5′ACGAACGATTTGCACGTCAG 3′; Scholin and Anderson 1994) were used to amplify a ca. 900 base pair region of the large-subunit (LSU) rRNA gene. The polymerase chain reaction (PCR) reactions contained a final volume of 50 μl consisting of 25 μl MYTAQ 2× master mix (Bioline, UK), 1–2 μl of template DNA (ca. 50 ng), 2 μl of each primer (0.5 μm final concentration) and made up to 50 μl with diethyl pyrocarbonate-treated water (Ambion, Thermo Fisher Scientific, Waltham, MA, USA). The reaction was then subjected to the following conditions in a thermo-cycler: an initial denaturation of 5 min at 95°C then 30 cycles of 94°C for 1 min, 55°C for 1 min, 72°C for 1 min and then a final extension step of 72°C for 3 min. The products were checked by electrophoresis using 1% agarose gel (prepared with 1× tris-borate-ethylenediaminetetraacetic acid) and then purified using an Invisorb® Fragment Cleanup. Sequences obtained in this study were subjected to an European Bioinformatics Institute (EMBL-EBI) Fasta 33 search (Pearson and Lipman 1988).
Phylogenetic trees were constructed using the Molecular Evolutionary Genetics Analysis (MEGA) version 7 (Kumar et al. 2016). Sequences from this study along with the matching top hits from the EMBL database and representative phylogenetic sequences from major groups of dinoflagellates were included in tree construction, Oxyrrhis marina Dujardin was used as an outgroup. The sequences were first aligned using ClustalW (Thompson et al. 1994). Phylogenetic analysis was performed using the maximum likelihood method based on the Tamura-Nei model (Tamura and Nei 1993). Bootstrap analysis (Felsenstein 1985) was also carried out with 500 replicates to provide confidence limits for tree branches. The tree was drawn to scale, with branch lengths measured in the number of substitutions per site. The analysis involved 43 nucleotide sequences. There were a total of 503 positions in the final dataset.
Results
Using the slurry culture approach, several hundred cells were isolated which resulted in some 30 successful cultures. Motile cells of Kryptoperidinium Lindemann, Gonyaulax Diesing, Gymnodinium F. Stein emend. Hansen et Moestrup, Lingulodinium Wall, Scrippsiella Balech and Woloszynskia Thompson were isolated (Table 2, Figures 2–31) and sequenced (Figure 32).
Dinoflagellate cultures isolated from the Caspian and Black Seas.
| Code | Station | Identification | Accession no. |
|---|---|---|---|
| CS-ST1-005 | Caspian Sea Station 1 | Gonyaulax baltica | KY921621 |
| CS-ST2-001 | Caspian Sea Station 2 | Gonyaulax sp. | |
| A3 | Caspian Sea HCGA09 | Gymnodinium aureolum | KY921616 |
| CS-ST7-009 | Caspian Sea Station 7 | Gymnodinium aureolum | KY921624 |
| CS-ST1-001 | Caspian Sea Station 1 | Kryptoperidinium foliaceum | KY921622 |
| CS-ST1-007 | Caspian Sea Station 1 | Kryptoperidinium foliaceum | |
| A6 | Caspian Sea HCGA09 | Kryptoperidinium foliaceum | KY921623 |
| CS-ST1-002–004 | Caspian Sea Station 1 | Lingulodinium polyedra | |
| CS-ST3-001 | Caspian Sea Station 3 | Lingulodinium polyedra | |
| CS-ST4-001–004 | Caspian Sea Station 4 | Lingulodinium polyedra | |
| CS-ST6-002–004 | Caspian Sea Station 6 | Lingulodinium polyedra | |
| D10 | Caspian Sea HCGA09 | Scrippsiella acuminata | KY921618 |
| B1 | Black Sea | Scrippsiella acuminata | KY921618 |
| CS-ST2-004 and 006 | Caspian Sea Station 2 | Scrippsiella acuminata | |
| D4 | Caspian Sea HCGA09 | Woloszynskia sp. | KY921615 |
| D5 | Caspian Sea HCGA09 | Woloszynskia sp. | KY921617 |
| D6 | Black Sea | Woloszynskia sp. | KY921620 |

Light micrographs of Caspian Sea dinoflagellates grown in culture from sediments.
(2–6) Gonyaulax baltica, culture CS-ST1-005. (2) Lateral view of cell showing cingulum offset. (3) Dorsal view of cell showing general shape and wide cingulum. (4) Antapical view of cell showing broad sulcus. (5) Ecdysed cell showing antapical spines. (6) Hypotheca of cell showing plate reticulation, smooth sulcus and intra plate growth bands. (7–10) Gonyaulax sp., culture CS-ST2-001. (7) Lateral view of cell showing general shape, cingulum offset, apical horn and antapical spine. (8) Ventral view of cell showing broad offset cingulum. (9) Dorsal view of cell in outline showing pronounced apical horn and definite shoulders. (10) Ecdysed theca showing solid antapical spine and heavy plate reticulation. (11–14) Gymnodinium aureolum. (11–13) Culture CS-ST7-009. (14) Culture A3. (11) Cell showing overall shape, central nucleus and radiating chloroplasts. (12) Ventral view of cell showing sulcal-cingulum arrangement. (13) Dorsal view of cell. (14) Cells in duplet. (15–20) Kryptoperidinium foliaceum. (15 and 19) Culture CS-ST1-007. (16 and 18) Culture CS-ST1-001. (17 and 20) Culture A6. (15) Ventral view of cell showing leaf-like curvature, central nucleus, median cingulum and eyespot. (16) Lateral view of cell showing dorso-ventral flattening and eyespot. (17) Theca stained with trypan blue. (18) Cysts in ventral view. (19) Cyst in lateral view. (20) Cysts in mucoid capsule. (21–25) Lingulodinium polyedra. (21 and 23) Culture CS-ST1-002. (22) Culture CS-ST1-004. (25) Culture CS-ST1-003. (21) Ventral view of cell showing cingulum offset and first apical plate. (22) Dorsal view of cell showing horse-shoe shaped nucleus. (23) Ecdysed hypotheca showing distinctive polyhedral shape and plate reticulation. (24) Squashed cell showing thecal plates. (25) Cyst from Station 1 sediment. (26–28) Scrippsiella acuminata. (26) Culture CS-ST2-006. (27–28) Culture D10. (26) Outline view of cell. (27) Outline view of cell. (28) Cyst formed in culture. (29–30) Woloszynskia sp. (29) Culture D3 showing cell with eyespot. (30) Cell showing general outline from culture D4. (31) Impagidinium caspienense from palynological preparation (core CS03/1 at 32 cm) showing archeopyle. Scale bars=10 μm.
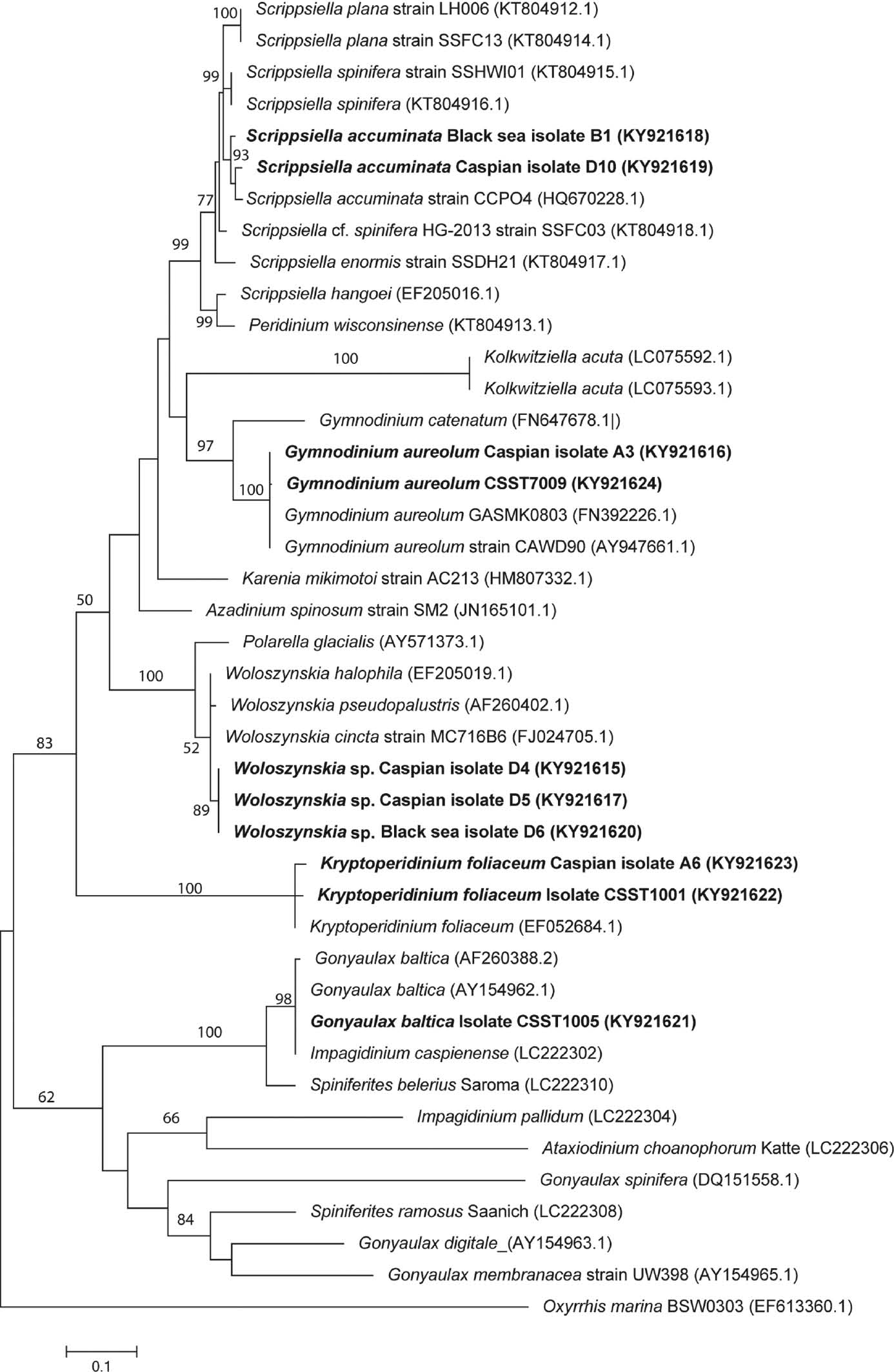
Molecular phylogeny of dinoflagellates isolated from the Caspian and Black Sea sediments inferred from partial large-subunit rDNA (LSU rDNA) sequences based on the maximum likelihood (ML) method.
Oxyrrhis marina was used as an outgroup. Numbers on branches represent ML bootstrap values for that node; bootstrap values >50% are shown. Sequences from isolates in this study are indicated in bold. The tree is drawn to scale, with branch lengths measured in the number of substitutions per site. Scale bar=nucleotide substitutions per site. The analysis involved 42 nucleotide sequences. Sequences generated in this study are available from GenBank, Accessions KY921615-KY921624. There were a total of 503 positions in the final dataset.
Gonyaulax Diesing 1866
Gonyaulax baltica Ellegaard, Lewis et Harding (Figures 2–6)
Motile cells were brownish-yellow in colour, 30–32 μm long and 25–28 μm wide. The cells had a conical epitheca with a short apical horn with slight shoulders and a rounded hypotheca (Figure 2). The wide median cingulum was offset by two cingulum widths (Figures 2 and 3). The sulcus is broad and smooth (Figures 4–6). Other thecal plates had clear reticulation (Figures 5 and 6). On some cells, short acuminate processes could be discerned on the hypotheca (Figure 5). Overall, the thecal morphology was attributable to G. baltica (Ellegaard et al. 2002). One strain was isolated from Caspian Sea material and was sequenced, emerging as identical to G. baltica isolated from the Baltic Sea and Impagidinium caspienense Marret isolated from the Caspian Sea (Figure 32).
Gonyaulax sp. (Figures 7–10)
Motile cells were brownish-yellow in colour, 35–40 μm long and 25–30 μm wide. Cells had a conical epitheca with a pronounced apical horn arising from distinct shoulders (Figures 7–9). The hypotheca was slightly flattened with a long (5–8 μm) single antapical horn. The wide median cingulum was offset by two cingulum widths. Thecal plates were strongly reticulate (Figure 10). This poorly growing strain did not survive to sequencing so observations as to identity can only be made on the basis of light microscopy. The cells can be compared to Gonyaulax digitale Kofoid and Gonyaulax elongata (Reid) Ellegaard, Daugbjerg, Rochon, Lewis et Harding for which cyst-theca relationships have been described (Lewis et al. 2001, Ellegaard et al. 2003). They resemble most closely the former with the exception of the single prominent antapical spine – no specimens being seen with two or more spines as is diagnostic for this species. Gonyaulax elongata is characterised by a single antapical flange but has a rather less pronounced apical horn with only weak shoulders in contrast to the cells observed here. Neither have either of these cyst types been recorded from Caspian Sea sediment. Table 1 shows there are several other gonyaulacoid cysts recorded in the sediment – Spiniferites belerius Reid, Spiniferites cruciformis Wall et Dale and Caspidinium rugosum Marret. From this study, it is not possible to determine which, if any, of these are linked to this Gonyaulax. Comparison to some 25 other described Gonyaulax species does not yield any clear affinities. At this point, we do not consider we have sufficient information to fully describe this species and prefer to wait to provide a description that would also include the cyst stage.
Gymnodinium F. Stein 1878 emend. Hansen et Moestrup 2000
Gymnodinium aureolum (Hulburt) Hansen (Figures 11–14)
Motile cells were yellow-brown in colour and were spherical in outline with a flattened antapex and slight dorso-ventral flattening (Figures 11–13). Cells were 30–38 μm long and 25–30 μm wide. The cingulum surrounded the middle of the cell with a slight offset (Figures 11 and 12). The nucleus was centrally placed (Figure 12). Divided cells were occasionally seen as duplets (Figure 14). Cells matched the description given for this species in Hansen et al. (2000). Two strains isolated from Caspian Sea material were successfully sequenced and are placed in the tree alongside strains of Gymnodinium aureolum from South Korea and New Zealand (Figure 32).
Kryptoperidinium Lindemann 1924
Kryptoperidinium foliaceum (F. Stein) Lindemann (Figures 15–20)
Cells were pale brown in colour with a red eye spot and a central nucleus (Figures 15 and 16). Cells were 30–50 μm long and 28–45 μm wide. Cells were strongly dorso-ventrally flattened and broadly circular in dorsal view (Figures 15 and 16). The cingulum was median and not offset (Figure 15). Cells had very thin thecae on which it was very difficult to discern any thecal tabulation (Figure 17) although it has been reported by Figueroa et al. (2009). Cysts were formed within our cultures – these were ovoid to spherical in dorsal view (Figure 18) and narrowly elliptical in apical view (Figures 19 and 20). Two strains from the Caspian were successfully sequenced which match those for Kryptoperidinium foliaceum (Figure 32).
Lingulodinium Wall 1967
Lingulodinium polyedra (F. Stein) Dodge (Figures 21–25)
Cells were brown in colour (Figure 21), 28–45 μm long and 28–45 μm wide. Cells showed a characteristic angular outline with flattened hypotheca and angular epitheca with a very small apical horn (Figure 22). Cingulum was median and offset by one cingulum width (Figure 21). Strong thecal plates with circular ridges around the trichocyst pores and ridges along plate boundaries (Figures 23 and 24). The nucleus was U-shaped and lying across the middle of the cell (Figure 22). Cysts were found in the sediment samples examined (Figure 25), and various spine lengths were noted, probably driven by the low salinities in the Caspian, as has been recorded by Mertens et al. (2012). Cultures of Lingulodinium were the most numerous in this study indicating their common occurrence in the sediment as well as ease of culturing.
Scrippsiella Balech 1959
Scrippsiella acuminata (Ehrenberg) Kretschmann, Elbrächter, Zinssmeister, S. Soehner, Kirsch, Kusber et Gottschling (Figures 26–28)
Cells were brown in colour with a pale central nucleus (Figure 26) and 21–26 μm long and 20–28 μm wide. Cells had a conical epitheca and a rounded hypotheca and were circular when seen in apical view (Figures 26 and 27). The cingulum was median and only slightly offset. Characteristic cysts were formed in one isolate (Figure 28). One Caspian and one Black Sea isolate were successfully sequenced and placed in the tree alongside Scrippsiella trochoidea (F. Stein) Loeblich III (considered a heterotypic synonym of S. acuminata by Kretschmann et al. 2015) from the Lafeyette River in the USA (Figure 32). Two other Scrippsiella species are of note to review here – S. plana and S. spinifera Honsell et Cabrini. In their paper describing S. plana from the Caspian Sea for the first time, Luo et al. (2016) clearly illustrate both species which each have characteristic motile cells. Scrippsiella plana has a distinctive flattened morphology and S. spinifera an elongate morphology with small antapical spines. The cultures developed in this study did not show these thecal morphologies; furthermore, a sequenced isolate formed characteristic S. trochoidea cysts. The taxonomy of Scrippsiella sensu lato remains enigmatic with further resolution awaiting sequencing of key species (Luo et al. 2016).
Woloszynskia Thompson 1951
Woloszynskia sp. (Figures 29 and 30)
Cells were small in comparison to other isolated species, averaging 9–10 μm long and 7–8 μm wide, and gymnodinioid in shape (Figures 29 and 30). Cells were orange/brown in colour. Eyespots were visible in the centre of cells (Figure 29) as has been reported in other Woloszynskia species (Siano et al. 2009). Cells swam fast with a distinctive whip-like movement. Cysts were ovoid in shape, brown in colour and ca. 9 μm long and 7–8 μm wide. Two isolates were obtained from the Caspian and one from the Black Sea and all three were successfully sequenced being placed in the phylogenetic tree alongside other Woloszynskia species (Figure 32).
Discussion
Gonyaulax baltica, Kolkwitziella acuta (Apstein) Elbrächter and Lingulodinium polyedra are the only species that have previously been recognised in both planktonic and sediment samples (Table 1). Of these, we have confirmed the presence of G. baltica and L. polyedra in Caspian Sea sediment. The cyst-theca relationship for Impagidinium caspienense (Figure 31) was recently elucidated by Mertens et al. (2017) as being linked to G. baltica. In common with other spiny gonyaulacoid cysts, the cyst morphology of G. baltica is strongly influenced by salinity and the form found in the Caspian is at one end of the spectrum of spine bearing with no process development evident. Thus when first described from Caspian sediment, rather than attribution to Spiniferites Mantell, it was attributed to the genus Impagidinium Stover et Evitt (Marret et al. 2004). However, the cyst form of G. baltica in its original description by Ellegaard et al. (2002) from Baltic sediment was of a more typical Spiniferites morphology although description was provided in that paper for substantial variation in form. Attribution to Impagidinium is questioned by Mertens et al. (2017) on the basis of some morphological features and also geography (other Impagidinium species being typically found offshore rather than in more coastal habitats). Further, molecular and detailed taxonomic studies involving both cysts and thecae in the wider Gonyaulax group provide evidence for the polyphyletic nature of Gonyaulax sensu lato. For now, however, they suggest the simplest solution is to retain the current dual classification of the cyst and motile stage, awaiting a more fundamental overhaul of Gonyaulax in the future. That there remains a great deal of work to clarify species in the Gonyaulacales has previously been highlighted (e.g. Lewis et al. 2001, Mertens et al. 2015a) and the presence of another unidentified Gonyaulax in this study also confirms that further work on the cyst-theca relationships in the Gonyaulacales is necessary.
Scrippsiella acuminata is recorded from Caspian sediment for the first time. Scrippsiella acuminata has previously been recognised in the plankton but not recorded in sediment samples. Scrippsiella species are common constituents of coastal sediment (Lewis 1991). Caspian sediment has largely been investigated using palynological techniques (references in Table 1). As a calcareous cyst with a thin inner organic wall, it seems likely that these cysts have been destroyed by the processing techniques used for these studies and hence the previous lack of records. This is the first record for Kryptoperidinium foliaceum, Gymnodinium aureolum and Woloszynskia sp. from the Caspian Sea. Woloszynskia pascheri (Suchlandt) von Stosch has been recorded from the Iranian rivers which flow into the Caspian (Zarei Darki 2009). It is possible that all of these species have been previously overlooked in the plankton samples – a number of Glenodinium Ehrenberg and Gymnodinium species have been recorded (see Table 1) some of which might be confused with these species. It can be especially difficult to speciate Gymnodinium species because of the difficulties of preserving them successfully. This was illustrated by Sundström et al. (2009) describing a new species of Gymnodinium from the extensively studied Baltic Sea which had not been recognised despite it being a relatively common member of the spring phytoplankton flora. Cysts have been described for both K. foliaceum (Figueroa et al. 2009) and G. aureolum (Tang et al. 2008), but neither has been recorded from Caspian sediment. Cysts are also known from the Woloszynskia genus (e.g. Kremp et al. 2005) but none has been recorded from Caspian sediment. Previous studies in the Caspian have been for geological purposes and so sediment has been processed by palynological means. It seems likely that the hyaline cysts of these species did not survive such harsh techniques or if they did, given their lack of paratabulation, they might not be recognised as such. However, the presence of cysts in sediment can be a useful way of providing an integrated record of occurrence of species that might be missed in the plankton and indeed K. foliaceum was recognised for the first time in the Mediterranean Sea by this route (Satta et al. 2013). Given the variety of Gymnodinium species recorded in the plankton, it would seem likely other cysts in this genus might also be present in the sediment.
Reviewing Table 1, other gaps can be highlighted between the two lists. Species of the following genera, Oblea Balech ex Loeblich et Loeblich III, Diplopsalis Bergh and some Protoperidinium Bergh, recorded in the plankton could be represented by the Brigantedinium Reid recorded in the dinocyst record. However, the presence of Protoperidinium leonis Pavillard in the plankton would suggest the distinctive Quinquecuspis Harland might also be recorded in the dinocyst record. It is hard to reconcile this lack – unless it is due to rarity. Heterocapsa triquetra (Ehrenberg) F. Stein recorded in the plankton also is not recorded in sediments. The presence of Pentapharsodinium dalei Indelicato et Loeblich III in the sedimentary record would suggest it should similarly be recorded in the plankton. Here there could be confusion with S. acuminata that is similar in morphology in the motile stage.
The approach of using sediment slurries has been invaluable for investigating the diversity of dinoflagellates in a region where intact cysts were relatively rare. Single cyst isolations are very helpful but, where there is a paucity of intact cysts, they are challenging. Using slurries offers the opportunity to develop cultures that can provide a wealth of taxonomic information as well as the possibility of cyst formation as demonstrated in this study. Our results also highlight the need for culturing to learn more about the interactions between motile stages and cysts, which would allow understanding of species origination over time linked to the palaeohydrological history of the Caspian Sea.
About the authors

Jane Lewis is a Professor in the Department of Life Sciences at the University of Westminster. Her research is focussed on the ecology and taxonomy of dinoflagellates. There have been two main themes for this research – the use of dinoflagellate cysts to interpret the fossil record and the role of microalgal life cycles in harmful algal blooms. Current research includes the role of temperature in modulating life cycle transitions in Alexandrium tamarense in the Gulf of Maine and the morphological variation of cysts in various species within the Gonyaulacales.

Joe D. Taylor is an independent research fellow at the University of Salford, UK. He was awarded a PhD in Environmental Microbiology by the University of Essex, UK, working on microbial degradation of algal polysaccharides in estuarine littoral zones. He has worked on across multiple habitats and with multiple taxonomy groups. His current research is focused on micro-eukaryote diversity and function in oceanic, coastal and terrestrial food-webs.

Suzanne A.G. Leroy was Professor at Brunel University London (UK) from 2000 to 2016. She is now an associate researcher at the CEREGE, France. She is a palynologist, specialising in the Caspian Sea palaeo-environmental history since 1994. She has pioneered the work on dinocysts from Caspian sediment and has stimulated their taxonomical and phylogenetical analyses. Other areas of current research include palaeo-climates and lake-level changes in the Middle East and Central Asia.
Acknowledgements
We thank M. Naderi-Beni (INIOAS, Tehran, Iran) for taking the grab samples; Fabienne Marret and Lee Bradley for stimulating our interest in the Caspian and for initial provision of sediment, and Sarah Rowing for assistance with culturing at the University of Westminster.
References
Bagheri, S. and M. Fallahi. 2014. Checklist of phytoplankton taxa in the Iranian waters of the Caspian Sea. Caspian J. Env. Sci. 12: 81–97.Search in Google Scholar
Bagheri, S., M. Mansor, M. Makaremi, A. Mirzajani, H. Babaei, H. Negarestan and W.O. Wan Maznah. 2010. Distribution and composition of phytoplankton in the Southwestern Caspian Sea during 2001–2002, a comparison with previous surveys. World J. Fish Mar. Sci. 2: 416–426.Search in Google Scholar
Bagheri, S., M. Mansor, M. Makaremi, J. Sabkara, W.O. Wan Maznah, A. Mirzajani, S.H. Khodaparast, H. Negarestan, A. Ghandi and A. Khalilpour. 2011. Fluctuations of phytoplankton community in the coastal waters of Caspian Sea in 2006. Am. J. Appl. Sci. 8: 1328–1336.10.3844/ajassp.2011.1328.1336Search in Google Scholar
Bagheri, S., M. Mansor, M. Turkoglu, M. Makaremi and H. Babaei. 2012a. Temporal distribution of phytoplankton in the south-western Caspian Sea during 2009 – 2010: a comparison with previous surveys. J. Mar. Biol. Ass. UK 92: 1243–1255.10.1017/S0025315412000094Search in Google Scholar
Bagheri, S., M. Mansor, M. Turkoglu, M. Makaremi, W.O. Wan Maznah and H. Negarestan. 2012b. Phytoplankton composition and abundance in the Southwestern Caspian Sea. Ekoloji 21: 32–43.10.5053/ekoloji.2012.834Search in Google Scholar
Bolch, C.J.S. 1997. The use of polytungstate for the separation and concentration of living dinoflagellate cysts from marine sediments. Phycologia 36: 472–478.10.2216/i0031-8884-36-6-472.1Search in Google Scholar
Dodge, J.D. 1982. Marine dinoflagellates of the British Isles. Her Majesty’s Stationery Office, London.Search in Google Scholar
Dumont, H.J. 1998. The Caspian Lake: history, biota, structure, and function. Limnol. Oceanogr. 43: 44–52.10.4319/lo.1998.43.1.0044Search in Google Scholar
Ellegaard, M., J. Lewis and I. Harding. 2002. Cyst-theca relationship, life cycle, and effects of temperature and salinity on the cyst morphology of Gonyaulax baltica sp. nov. (Dinophyceae) from the Baltic Sea area. J. Phycol. 38: 775–789.10.1046/j.1529-8817.2002.01062.xSearch in Google Scholar
Ellegaard, M., N. Daugbjerg, A. Rochon, J. Lewis and I. Harding. 2003. Morphological and LSU rDNA sequence variation within the Gonyaulax spinifera-Spiniferites group (Dinophyceae) and proposal of G. elongata comb. nov. and G. membranacea comb. nov. Phycologia 42: 151–164.10.2216/i0031-8884-42-2-151.1Search in Google Scholar
Felsenstein, J. 1985. Confidence limits on phylogenies: An approach using the bootstrap. Evolution 39: 783–791.10.1111/j.1558-5646.1985.tb00420.xSearch in Google Scholar PubMed
Figueroa, R.I., I. Bravo, S. Fraga, E. Garcés and G. Llaveria. 2009. The life history and cell cycle of Kryptoperidinium foliaceum, a dinoflagellate with two eukaryotic nuclei. Protist 160: 285–300.10.1016/j.protis.2008.12.003Search in Google Scholar PubMed
Ganjian, A., W.O. Wan Maznah, K. Yahya, H. Fazli, M. Vahedi, A. Roohi and S.M.V. Farabi. 2010. Seasonal and regional distribution of phytoplankton in the southern part of the Caspian Sea. Iran. J. Fish. Sci. 9: 382–401.Search in Google Scholar
Gogorev, R. 2006. Check-list for Caspian Sea phytoplankton. In: Caspian Sea Biodiversity Project; http://www.zin.ru/projects/caspdiv/caspian_phytoplankton.html [accessed 22 February 2017].Search in Google Scholar
Gómez, F. 2003. Checklist of Mediterranean free-living dinoflagellates. Bot. Mar. 46: 215–242.10.1515/BOT.2003.021Search in Google Scholar
Gómez, F. and L. Boicenco. 2004. An annotated checklist of dinoflagellates in the Black Sea. Hydrobiologia 517: 43–59.10.1023/B:HYDR.0000027336.05452.07Search in Google Scholar
Guillard, R.R.L. 1973. Methods for microflagellates and nano-plankton. In: (J.R. Stein, ed.) Handbook of Phycological Methods: Culture Methods and Growth Measurements, Cambridge University Press, Cambridge. pp. 69–85.Search in Google Scholar
Guiry, M.D. and G.M. Guiry. 2017. AlgaeBase. World-wide electronic publication, National University of Ireland, Galway. http://www.algaebase.org [accessed between 15 February and 10 March 2017].Search in Google Scholar
Hansen, G., N. Daugbjerg and P. Henriksen. 2000. Comparative study of Gymnodinium mikimotoi and Gymnodinium aureolum, comb. nov. (=Gyrodinium aureolum) based on morphology, pigment composition, and molecular data. J. Phycol. 36: 394–410.10.1046/j.1529-8817.2000.99172.xSearch in Google Scholar
Hoppenrath, M., M. Elbrächter and G. Drebes. 2009. Marine Phytoplankton. Selected species from the North Sea around Helgoland and Sylt. Kleine Senckenberg-Reihe, Band 49, Stuttgart.Search in Google Scholar
Kazancı, N., T. Gulbabazadeh, S.A.G. Leroy and Ö. Ileri. 2004. Sedimentary and environmental characteristics of the Gilan-Mazenderan plain, northern Iran: influence of long- and short-term Caspian water level fluctuations on geomorphology. J. Mar. Syst. 46: 145–168.10.1016/j.jmarsys.2003.12.002Search in Google Scholar
Kideys, A.E., N. Soydemir, E. Eker, V. Vladymyrov, D. Soloviev and F. Mélin. 2005. Phytoplankton distribution in the Caspian Sea during March 2001. Hydrobiologia 543: 159–168.10.1007/s10750-004-6953-xSearch in Google Scholar
Kideys, A.E., A. Roohi, E. Eker-Develi, F. Mélin and D. Beare. 2008. Increased chlorophyll levels in the Southern Caspian Sea following an invasion of jellyfish. Res. Lett. Ecol. 2008. Article ID 185642. Doi: 10.1155/2008/185642.10.1155/2008/185642Search in Google Scholar
Kosarev, A.N. and E.A. Yablonskaya. 1994. The Caspian Sea. Transl. from Russian by A.K. Wistin. SPB Academic Publishing, The Hague.Search in Google Scholar
Kremp, A., M. Elbrächter, M. Schweikert, J.L. Wolny and M. Gottschling. 2005. Woloszynskia halophile (Biecheler) comb. nov.: a bloom-forming cold-water dinoflagellate co-occurring with Scrippsiella hangoei (Dinophyceae) in the Baltic Sea. J. Phycol. 41: 629–642.10.1111/j.1529-8817.2005.00070.xSearch in Google Scholar
Kretschmann, J., M. Elbrächter, C. Zinssmeister, S. Soehner, M. Kirsch, W-H Kusber and M. Gottschling. 2015. Taxonomic clarification of the dinophyte Peridinium acuminatum Ehrenb., ≡ Scrippsiella acuminata, comb. nov. (Thoracosphaeraceae, Peridiniales). Phytotaxa 220: 239–256.10.11646/phytotaxa.220.3.3Search in Google Scholar
Kumar, S., G. Stecher and K. Tamura. 2016. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for bigger datasets. Mol. Biol. Evol. 33: 1870–1874.10.1093/molbev/msw054Search in Google Scholar PubMed PubMed Central
Lenaers, G., L. Maroteaux, B. Michot and M. Herzog. 1989. Dinoflagellates in evolution. A molecular phylogenetic analysis of large subunit ribosomal RNA. J. Mol. Evol. 29: 40–51.10.1007/BF02106180Search in Google Scholar PubMed
Leroy, S.A.G. 2010. Palaeoenvironmental and palaeoclimatic changes in the Caspian Sea region since the Late glacial from palynological analyses of marine sediment cores. Geography, Environment, Sustainability 2: 32–41.10.24057/2071-9388-2010-3-2-32-41Search in Google Scholar
Leroy, S.A.G., F. Marret, S. Giralt and S.A. Bulatov. 2006. Natural and anthropogenic rapid changes in the Kara-Bogaz Gol over the last two centuries by palynological analyses. Quat. Int. 150: 52–70.10.1016/j.quaint.2006.01.007Search in Google Scholar
Leroy, S.A.G., F. Marret, E. Gibert, F. Chalié, J.-L. Reyss and K. Arpe. 2007. River inflow and salinity changes in the Caspian Sea during the last 5500 years. Quat. Sci. Rev. 26: 3359–3383.10.1016/j.quascirev.2007.09.012Search in Google Scholar
Leroy, S.A.G., H.A.K. Lahijani, M. Djamali, A. Naqinezhad, M.V. Moghadam, K. Arpe, M. Shah-Hosseini, M. Hosseindoust, Ch.S. Miller, V. Tavakoli, P. Habibi and M. Naderi Beni. 2011. Late Little Ice Age palaeoenvironmental records from the Anzali and Amirkola lagoons (south Caspian Sea): vegetation and sea level changes. Palaeogeogr. Palaeoclimatol. Palaeoecol. 302: 415–434.10.1016/j.palaeo.2011.02.002Search in Google Scholar
Leroy, S.A.G., A.A. Kakroodi, S.B. Kroonenberg, H.A.K. Lahijani, H. Alimohammadian and A. Nigarov. 2013a. Holocene vegetation history and sea level changes in the SE corner of the Caspian Sea: relevance to SW Asia climate. Quat. Sci. Rev. 70: 28–47.10.1016/j.quascirev.2013.03.004Search in Google Scholar
Leroy, S.A.G., H.A.K. Lahijani, J.-L. Reyss, F. Chalié, S. Haghani, M. Shah-Hosseini, S. Shahkarami, A. Tudryn, K. Arpe, P. Habibi, H.S. Nasrollahzadeh and A. Makhlough. 2013b. A two-step expansion of the dinocyst Lingulodinium machaerophorum in the Caspian Sea: the role of changing environment. Quat. Sci. Rev. 77: 31–45.10.1016/j.quascirev.2013.06.026Search in Google Scholar
Leroy, S.A.G., A. Tudryn, F. Chalié, L. López-Merino and F. Gasse. 2013c. From the Allerød to the mid-Holocene: palynological evidence from the south basin of the Caspian Sea. Quat. Sci. Rev. 78: 77–97.10.1016/j.quascirev.2013.07.032Search in Google Scholar
Leroy, S.A.G., L. López-Merino, A. Tudryn, F. Chalié and F. Gasse. 2014. Late Pleistocene and Holocene palaeoenvironments in and around the Middle Caspian Basin as reconstructed from a deep-sea core. Quat. Sci. Rev. 101: 91–110.10.1016/j.quascirev.2014.07.011Search in Google Scholar
Lewis, J. 1991. Cyst-theca relationships in Scrippsiella (Dinophyceae) and related orthoperidinioid genera. Bot. Mar. 34: 91–106.10.1515/botm.1991.34.2.91Search in Google Scholar
Lewis, J., A. Rochon, M. Ellegaard, P. Mudie and I. Harding. 2001. The cyst–theca relationship of Bitectatodinium tepikiense (Dinophyceae). Eur. J. Phycol. 36: 137–146.10.1017/S0967026201003171Search in Google Scholar
Luo, Z., K.N. Mertens, S. Bagheri, H. Aydin, Y. Takano, K. Matsuoka, F.M.G. McCarthy and H. Gu. 2016. Cyst-theca relationship and phylogenetic positions of Scrippsiella plana sp. nov. and S. spinifera (Peridiniales, Dinophyceae). Eur. J. Phycol. 51: 1–15.Search in Google Scholar
Marret, F., S. Leroy, F. Chalié and F. Gasse. 2004. New organic-walled dinoflagellate cysts from recent sediments of Central Asian seas. Rev. Palaeobot. Palynol. 129: 1–20.10.1016/j.revpalbo.2003.10.002Search in Google Scholar
Mertens, K.N., L.R. Bradley, Y. Takano, P.J. Mudie, F. Marret, A.E. Aksu, R.N. Hiscott, T.J. Verleye, E.A. Mousing, L.L. Smyrnova, S. Bagheri, M. Mansor, V. Pospelova and K. Matsuoka. 2012. Quantitative estimation of Holocene surface salinity variation in the Black Sea using dinoflagellate cyst process length. Quat. Sci. Rev. 39: 45–59.10.1016/j.quascirev.2012.01.026Search in Google Scholar
Mertens, K.N., H. Aydin, S. Uzar, Y. Takano, A. Yamaguchi and K. Matsuoka. 2015a. Relationship between the dinoflagellate cyst Spiniferites pachydermus and Gonyaulax ellegaardiae sp. nov. from Izmir Bay, Turkey, and molecular characterization. J. Phycol. 51: 560–573.10.1111/jpy.12304Search in Google Scholar PubMed
Mertens, K.N., Y. Takano, A. Yamaguchi, H. Gu, K. Bogus, A. Kremp, S. Bagheri, G. Matisho and K. Matsuoka. 2015b. The molecular characterization of the enigmatic dinoflagellate Kolkwitziella acuta reveals an affinity to the Excentrica section of the genus Protoperidinium. Syst. Biodivers. 13: 509–524.10.1080/14772000.2015.1078855Search in Google Scholar
Mertens, K.N., Y. Takano, H. Gu, S. Bagheri, V. Pospelova, A.J. Pienńkowski, S. Leroy and K. Matsuoka. 2017. Cyst-theca relationship and phylogenetic position of Impagidinium caspienense incubated from Caspian Sea surface sediments: relation to Gonyaulax baltica and evidence for heterospory within gonyaulacoid dinoflagellates. J. Eukaryot. Microbiol. 64: 829–842.10.1111/jeu.12417Search in Google Scholar PubMed
Nasrollahzadeh, H.S., Z.B. Din, S.Y. Foong and A. Makhlough. 2008a. Trophic status of the Iranian Caspian Sea based on water quality parameters and phytoplankton diversity. Cont. Shelf Res. 28: 1153–1165.10.1016/j.csr.2008.02.015Search in Google Scholar
Nasrollahzadeh, H.S., Z.B. Din, S.Y. Foong and A. Makhlough. 2008b. Spatial and temporal distribution of macronutrients and phytoplankton before and after the invasion of the ctenophore, Mnemiopsis leidyi, in the Southern Caspian Sea. Chem. Ecol. 24: 1–14.10.1080/02757540802310967Search in Google Scholar
Nasrollahzadeh, H.S., A. Maklough, R. Pourgholam, F. Vahedi, A. Qanqermeh and S.Y. Foong. 2011. The study of Nodularia spumigena bloom event in the southern Caspian Sea. App. Ecol. Env. Res. 9: 141–155.10.15666/aeer/0902_141155Search in Google Scholar
Nasrollahzadeh, H.S., A. Makhlough, F. Eslami and S.A.G. Leroy. 2014. Features of the phytoplankton community in the Southern Caspian Sea, a decade after the invasion of Mnemiopsis leidyi. Iran. J. Fish. Sci. 13: 145–167.Search in Google Scholar
Pautova, L.A., M.D. Kravchishina, S.V. Vostokov, V.V. Zernova and V.A. Silkin. 2015. Features of the vertical phytoplankton structure in the deep-sea parts of the Caspian Sea in summer. Doklady Earth Sci. 462: 604–608.10.1134/S1028334X15060057Search in Google Scholar
Pearson, W.R. and D.J. Lipman. 1988. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA 85: 2444–2448.10.1073/pnas.85.8.2444Search in Google Scholar PubMed PubMed Central
Roohi, A., A.E. Kideys, A. Sajjadi, A. Hashemian, R. Pourgholam, H. Fazli, A.G. Khanari and E. Eker-Develi. 2010. Changes in biodiversity of phytoplankton, zooplankton, fishes and macrobenthos in the Southern Caspian Sea after the invasion of the ctenophore Mnemiopsis Leidyi. Biol. Invasions 12: 2343–2361.10.1007/s10530-009-9648-4Search in Google Scholar
Satta, C.T., S. Anglès, A. Lugliè, J. Guillén, N. Sechi, J. Camp and E. Garcés. 2013. Studies on dinoflagellate cyst assemblages in two estuarine Mediterranean bays: a useful tool for the discovery and mapping of harmful algal species. Harmful Algae 24: 65–79.10.1016/j.hal.2013.01.007Search in Google Scholar
Scholin, C.A. and D.M. Anderson. 1994. Identification of group- and strain-specific genetic markers for globally distributed Alexandrium (Dinophyceae). RFLP analysis of SSU rRNA genes. J. Phycol. 30: 999–1011.10.1111/j.0022-3646.1994.00999.xSearch in Google Scholar
Siano, R., W.H.C.F. Kooistra, M. Montresor and A. Zingone. 2009. Unarmoured and thin-walled dinoflagellates from the Gulf of Naples, with the description of Woloszynskia cincta sp. nov. (Dinophyceae, Suessiales). Phycologia 48: 44–65.10.2216/08-61.1Search in Google Scholar
Steidinger, K.A. 1997. Dinoflagellates. In: (C.R. Tomas, ed.) Identifying Marine Phytoplankton. Academic Press, San Diego. pp. 387–584.10.1016/B978-012693018-4/50005-7Search in Google Scholar
Sundström, A.M., A. Kremp, N. Daugbjerg, Ø. Moestrup, M. Ellegaard, R. Hansen and S. Hajdu. 2009. Gymnodinium corollarium sp. nov. (Dinophyceae) – A new cold-water dinoflagellate responsible for cyst sedimentation events in the Baltic Sea. J. Phycol. 45: 938–952.10.1111/j.1529-8817.2009.00712.xSearch in Google Scholar PubMed
Tamura, K. and M. Nei. 1993. Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Mol. Biol. Evol. 10: 512–526.Search in Google Scholar
Tang, Y.Z., T.A. Egerton, L. Kong and H.G. Marshall. 2008. Morphological variation and phylogenetic analysis of the dinoflagellate Gymnodinium aureolum from a tributary of Chesapeake Bay. J. Eukaryot. Microbiol. 55: 91–99.10.1111/j.1550-7408.2008.00305.xSearch in Google Scholar PubMed
Thompson, J.D., D.G. Higgins and T.J. Gibson. 1994. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22: 4673–4680.10.1093/nar/22.22.4673Search in Google Scholar PubMed PubMed Central
Zarei Darki, B. 2009. Algal flora of rivers in Iran. Int. J. Algae 11: 310–320.10.1615/InterJAlgae.v11.i2.70Search in Google Scholar
©2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and ecology
- Hydrodynamics in Indo-Pacific seagrasses with a focus on short canopies
- Thermal adaptation in a marine-derived tropical strain of Fusarium equiseti and polar strains of Pseudogymnoascus spp. under different nutrient sources
- Taxonomy/phylogeny and biogeography
- Expanding known dinoflagellate distributions: investigations of slurry cultures from Caspian Sea sediment
- The Chlorophytes of Curaçao (Caribbean): a revised checklist for the south-west coast
- Taxonomic reassessment of the Indo-Pacific Scytosiphonaceae (Phaeophyceae): Hydroclathrus rapanuii sp. nov. and Chnoospora minima from Easter Island, with proposal of Dactylosiphon gen. nov. and Pseudochnoospora gen. nov.
- Labyrinthulomycota from Brazilian mangrove swamps and coastal waters
- Phylogenetic survey and antimicrobial activity of cultivable fungi associated with five scleractinian coral species in the South China Sea
- First report of the invasive macroalga Acrothamnion preissii (Rhodophyta, Ceramiales) in the Atlantic Ocean
- Chemistry and applications
- Monthly variation in the chemical composition and biological activity of Sargassum horridum
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and ecology
- Hydrodynamics in Indo-Pacific seagrasses with a focus on short canopies
- Thermal adaptation in a marine-derived tropical strain of Fusarium equiseti and polar strains of Pseudogymnoascus spp. under different nutrient sources
- Taxonomy/phylogeny and biogeography
- Expanding known dinoflagellate distributions: investigations of slurry cultures from Caspian Sea sediment
- The Chlorophytes of Curaçao (Caribbean): a revised checklist for the south-west coast
- Taxonomic reassessment of the Indo-Pacific Scytosiphonaceae (Phaeophyceae): Hydroclathrus rapanuii sp. nov. and Chnoospora minima from Easter Island, with proposal of Dactylosiphon gen. nov. and Pseudochnoospora gen. nov.
- Labyrinthulomycota from Brazilian mangrove swamps and coastal waters
- Phylogenetic survey and antimicrobial activity of cultivable fungi associated with five scleractinian coral species in the South China Sea
- First report of the invasive macroalga Acrothamnion preissii (Rhodophyta, Ceramiales) in the Atlantic Ocean
- Chemistry and applications
- Monthly variation in the chemical composition and biological activity of Sargassum horridum