The role of GFAP and vimentin in learning and memory
-
Ulrika Wilhelmsson


Abstract
Intermediate filaments (also termed nanofilaments) are involved in many cellular functions and play important roles in cellular responses to stress. The upregulation of glial fibrillary acidic protein (GFAP) and vimentin (Vim), intermediate filament proteins of astrocytes, is the hallmark of astrocyte activation and reactive gliosis in response to injury, ischemia or neurodegeneration. Reactive gliosis is essential for the protective role of astrocytes at acute stages of neurotrauma or ischemic stroke. However, GFAP and Vim were also linked to neural plasticity and regenerative responses in healthy and injured brain. Mice deficient for GFAP and vimentin (GFAP−/−Vim−/−) exhibit increased post-traumatic synaptic plasticity and increased basal and post-traumatic hippocampal neurogenesis. Here we assessed the locomotor and exploratory behavior of GFAP−/−Vim−/− mice, their learning, memory and memory extinction, by using the open field, object recognition and Morris water maze tests, trace fear conditioning, and by recording reversal learning in IntelliCages. While the locomotion, exploratory behavior and learning of GFAP−/−Vim−/− mice, as assessed by object recognition, the Morris water maze, and trace fear conditioning tests, were comparable to wildtype mice, GFAP−/−Vim−/− mice showed more pronounced memory extinction when tested in IntelliCages, a finding compatible with the scenario of an increased rate of reorganization of the hippocampal circuitry.
Introduction
Astrocytes in the central nervous system (CNS) participate in and control many aspects of CNS plasticity both in health and disease (Eroglu and Barres, 2010; Pekna et al., 2012; Pekny and Pekna, 2014; Pekny et al., 2016, 2019; Stogsdill et al., 2017; Verkhratsky and Nedergaard, 2018). Some of these functions seem to depend on astrocyte intermediate filament system (Pekny et al., 2014; Pekny and Pekna, 2016), a part of the cytoskeleton most often viewed as conferring resilience towards cellular stresses (Pekny and Lane, 2007). Indeed, mice carrying null mutations in the GFAP and vimentin genes (GFAP−/−Vim−/− mice) and consequently having their astrocytes completely devoid of cytoplasmic intermediate filaments (Eliasson et al., 1999; Pekny et al., 1999), show reduced ability to handle acute cellular stress (Wilhelmsson et al., 2004; Nakazawa et al., 2007; Li et al., 2008; de Pablo et al., 2013). However, the GFAP−/−Vim−/− mice also exhibit increased synaptic plasticity in the hippocampus following its partial de-afferentation by enthorhinal cortex lesions (Wilhelmsson et al., 2004) and increased basal (Larsson et al., 2004; Wilhelmsson et al., 2012), post-traumatic (Wilhelmsson et al., 2012) and post-ischemic (Jarlestedt et al., 2010) neurogenesis. The GFAP−/−Vim−/− mice also support better long-term integration of neural grafts (Kinouchi et al., 2003), increased neuronal differentiation of neural stem/progenitor cells grafted in the hippocampus (Widestrand et al., 2007) and better regeneration of severed optic nerve (Cho et al., 2005).
Adult hippocampal neurogenesis in mice and other mammals results in the reorganization of the hippocampal circuitry (Laplagne et al., 2006; Tashiro et al., 2006) and increased hippocampal neurogenesis was linked both to better learning and increased forgetting of already acquired memories (Deng et al., 2010; Sahay et al., 2011; Akers et al., 2014). Despite many reports on the GFAP−/−Vim−/− mice and a range of disease models these were exposed to (Lundkvist et al., 2004; Wilhelmsson et al., 2004; Nakazawa et al., 2007; Li et al., 2008; Verardo et al., 2008; Jarlestedt et al., 2010; Lu et al., 2011; Macauley et al., 2011; Berg et al., 2013; Kraft et al., 2013; Liu et al., 2014; Kamphuis et al., 2015; Wunderlich et al., 2015; Laterza et al., 2018), their behavior including learning and memory has not yet been systematically investigated. Here we applied a battery of tests to evaluate the potential effects of increased hippocampal neurogenesis in GFAP−/−Vim−/− mice on the exploratory behavior, learning and memory extinction.
Results
GFAP−/−Vim−/− mice exhibit normal locomotor and exploratory behavior
To assess the locomotor and exploratory behavior, we used the open field test. GFAP−/−Vim−/− and wildtype mice showed comparable speed of locomotion, distance travelled, activity, rearing, grooming and open space exploration (Table 1), suggesting normal locomotor and exploratory behavior.
Locomotor and exploratory behavior.
| Parameter | WT | GFAP−/−Vim−/− |
|---|---|---|
| Average speed (cm/s) | 5.34±0.19 | 5.95±0.26 |
| Total distance travelled (cm) | 3203±112 | 3569±155 |
| Activity (% of total time) | 48.2±1.1 | 50.7±1.4 |
| Number of rearings | 34.8±1.62 | 32.1±1.83 |
| Number of groomings | 5.53±0.42 | 5.87±0.65 |
| Distance travelled in the center zone (% of total distance travelled) | 11.72±1.38 | 9.56±0.82 |
| Time spent in the center zone (% of total time) | 6.16±0.87 | 4.62±0.47 |
Two-month-old male wildtype (WT; n=15) and GFAP−/−Vim−/− (n=15) mice were assessed in the open field test. Data are presented as a mean±SEM; there were no differences between the groups as assessed by two-tailed t test.
Normal short- and long-term memory retrieval of GFAP−/−Vim−/− mice in the object recognition test
Given the 74% increase in the hippocampal neurogenesis of GFAP−/−Vim−/− mice (Wilhelmsson et al., 2012) and the association between neurogenesis and cognitive functions (Deng et al., 2010; Sahay et al., 2011; Akers et al., 2014), we subjected the mice to three different learning and memory tests. To assess declarative memory, we used the object recognition test, a task based on the natural preference of mice for exploring novel rather than familiar objects. During the training session with two identical objects presented, GFAP−/−Vim−/− and wildtype mice showed comparable total object exploration time (Figure 1A). When an object was replaced by a novel one at 6 and 24 h after training session, both GFAP−/−Vim−/− and wildtype mice showed exploration preference towards the novel object, with no differences between the groups (Figure 1B). Thus, GFAP−/−Vim−/− mice show normal short- and long-term memory retrieval in the object recognition test.
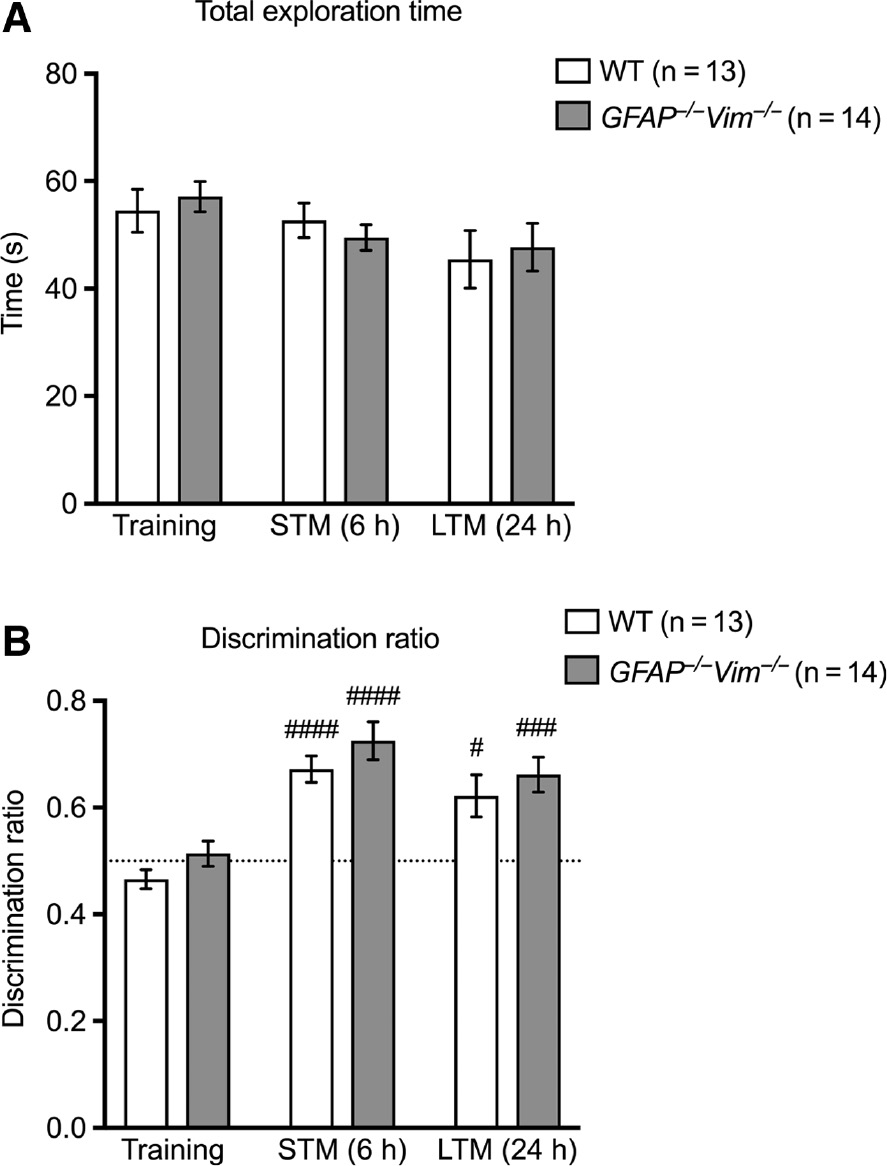
Normal long-term and short-term memory as assessed by the object recognition test.
The total time of exploration (A) was comparable between GFAP−/−Vim−/− and wildtype (WT) mice (2 months old) during the sessions of training and assessments of short-term memory (STM, 6 h) and long-term memory (LTM, 24 h). The discrimination ratio (B) shows that both groups of mice spent more time exploring the novel object during STM and LTM sessions compared to the training, while there were no differences in discrimination ratio between GFAP−/−Vim−/− and WT mice in STM and LTM as analyzed by analysis of variance (ANOVA) followed by the Holm Sidak post hoc test. n=13–14 mice per group. ####p<0.001; ###p<0.005; #p<0.05 for Training vs. STM and vs. LTM, for individual groups.
Normal spatial learning and memory retrieval of GFAP−/−Vim−/− mice in the Morris water maze test
We assessed spatial learning and memory using the Morris water maze test in which mice are trained to find a platform hidden below water surface in a pool. Both GFAP−/−Vim−/− and wildtype mice learned to find the platform over 8 days of training, as shown by the latency to find the platform and by the total distance swum, with no difference in learning curves between GFAP−/−Vim−/− and wildtype mice (Figure 2A–B). The average speed of swimming was also comparable between groups (Figure 2C). At day 9 and 15, the memory retention of the platform location was assessed in probe trials, in which the platform was removed. In both trials, GFAP−/−Vim−/− and wildtype mice showed comparable performance as assessed by time spent in platform quadrant, the latency to the first crossing of platform area, and the number of platform area crossings (Figure 2D). These results point to a normal spatial learning and memory retrieval of GFAP−/−Vim−/− mice.
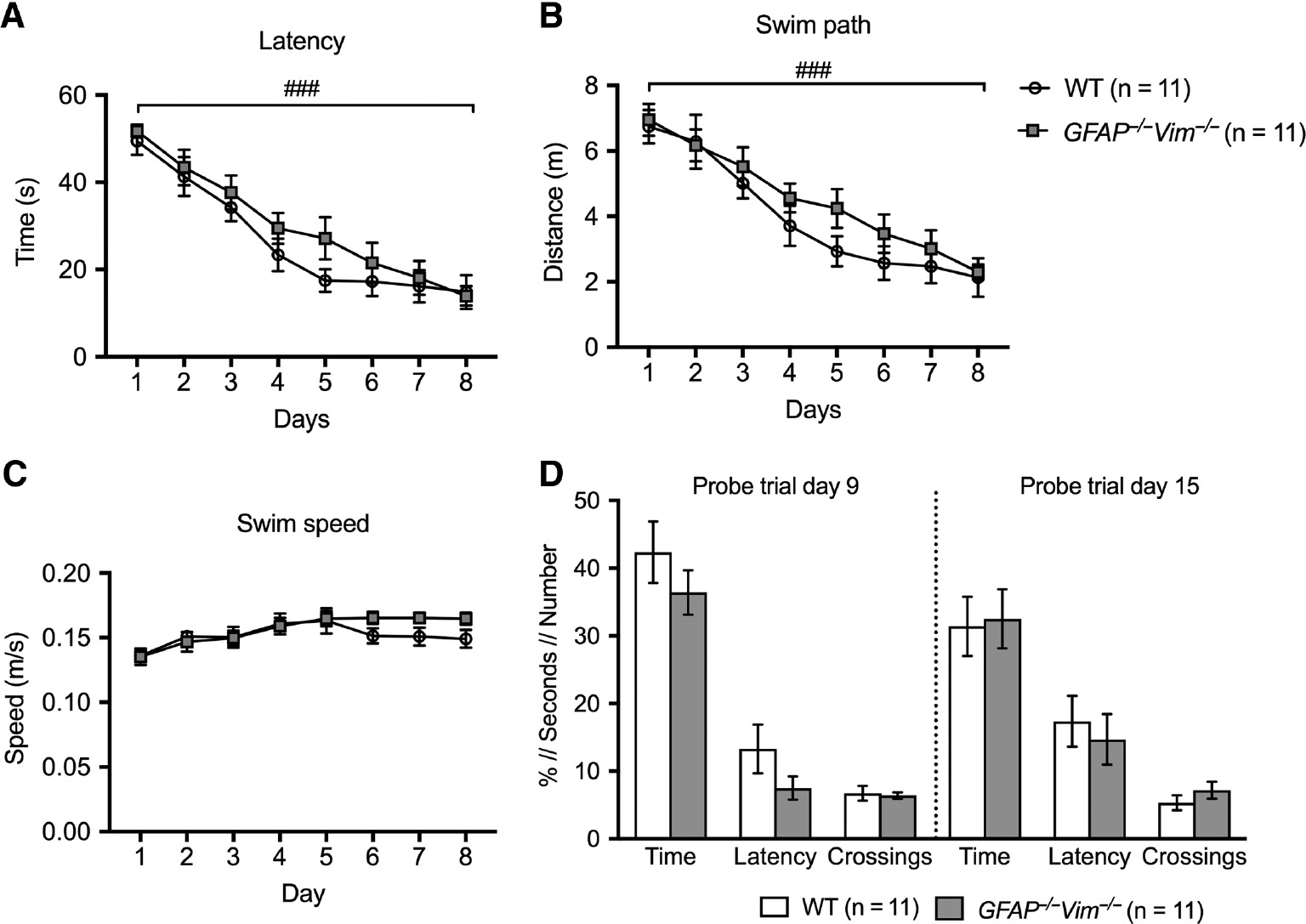
Comparable spatial learning and memory retrieval of GFAP−/−Vim−/− and wildtype mice in the Morris water maze test.
Three-month-old mice were trained and learned to find a hidden platform over 8 consecutive days with no difference between GFAP−/−Vim−/− and wildtype (WT) mice in the latency (A), the distance swum to locate the platform (B), and the swim speed (C). On day 9 and 15, probe trials were performed with the platform absent, with GFAP−/−Vim−/− and WT mice showing comparable performance as assessed by time spent in platform quadrant (Time), latency to first platform area crossing (Latency), and number of platform area crossings (Crossings) (D). The data were analyzed by two-way repeated-measures ANOVA (A–C), probe trial data in D by two-tailed t test. n=11 mice per group. ###p<0.001 for the time factor (A and B).
Normal short- and long-term memory retrieval of GFAP−/−Vim−/− mice in contextual and trace fear conditioning tests
Fear associated memory is dependent on amygdala and hippocampal functional circuits (Curzon et al., 2009) and was assessed by contextual and trace fear conditioning tests. GFAP−/−Vim−/− and wildtype mice were conditioned to an unpleasant electric impulse associated with an auditory cue after a time delay of 20 s. The next day, the natural fear response was assessed by the extent of freezing behavior in the same conditioned context but without the unpleasant stimulus or the auditory cue. GFAP−/−Vim−/− and wildtype mice showed comparable freezing response, indicating normal fear-associated memory of GFAP−/−Vim−/− mice (Figure 3A). Associative fear memory was also assessed by measuring the freezing response to the conditioned auditory cue in a novel context 1 day and 2 months after the initial fear conditioning. GFAP−/−Vim−/− and wildtype mice showed comparable initial freezing behavior when placed in the novel context before the presentation of the auditory cue (Figure 3B, pre CS), and a comparable increase in the freezing response after the auditory cue presentation (Figure 3B, CS-pre CS), both at 1 day and 2 months after the conditioning. This suggests that GFAP−/−Vim−/− mice have normal fear-associated memory and its retention.
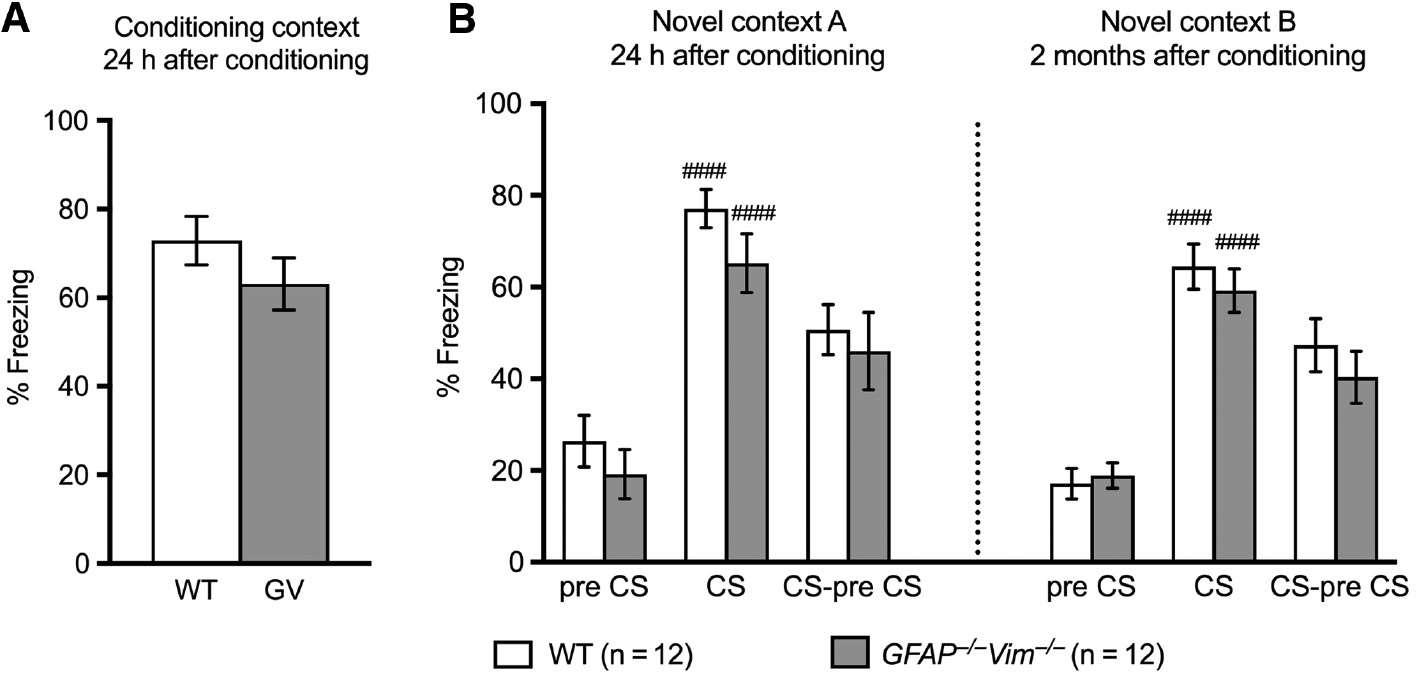
GFAP−/−Vim−/− and wildtype mice show comparable associative learning and memory in contextual and cued trace fear conditioning tests.
Freezing behavior of mice in response to the conditioning context assessed 24 h after conditioning (A), and in response to the conditioned stimulus (CS) in a new context (B) assessed 24 h and 2 months after conditioning showed no differences between GFAP−/−Vim−/− and wildtype (WT) mice. n=12 mice per group. Differences between groups were analyzed by two-tailed t test (A) or (B) two-way repeated-measures ANOVA followed by the post hoc Dunnett test (pre CS vs. CS; ####p<0.0001) or the Sidak test (WT vs. GFAP−/−Vim−/−, p>0.05).
GFAP−/−Vim−/− mice show increased memory extinction of place learning when recorded in the IntelliCage housing environment
To assess place learning and memory extinction over days, we used the IntelliCage system, which allows continuous automated monitoring and data collection (Karlsson et al., 2011). Groups of mice were housed over several days in cages in which water is accessed by a nose poke within chambers in all four corners of the cage. During the learning trial, the water access was limited to one of the four corners. After 5 days, the water access was changed to the opposite corner for additional 5 days to assess reversal learning and the place memory extinction for the previous drinking corner (Figure 4A). During the reversal learning and memory extinction period, GFAP−/−Vim−/− and wildtype mice showed normal learning as assessed by the increasing number of visits to the new drinking corner over time (Figure 4B, dotted lines) and comparable place memory extinction as assessed by reduced number of visits to the previous drinking corner over time (Figure 4B, continuous lines). Interestingly, GFAP−/−Vim−/− mice exhibited a reduced number of nose pokes per visit in the previous drinking corner compared to wildtype mice, showing that GFAP−/−Vim−/− mice were less persistent in trying to retrieve water in the previous drinking corner, a parameter that serves as a measure of memory extinction learning (Karlsson et al., 2011; Kalm et al., 2013). These results indicate that the GFAP−/−Vim−/− mice exhibit increased memory extinction.
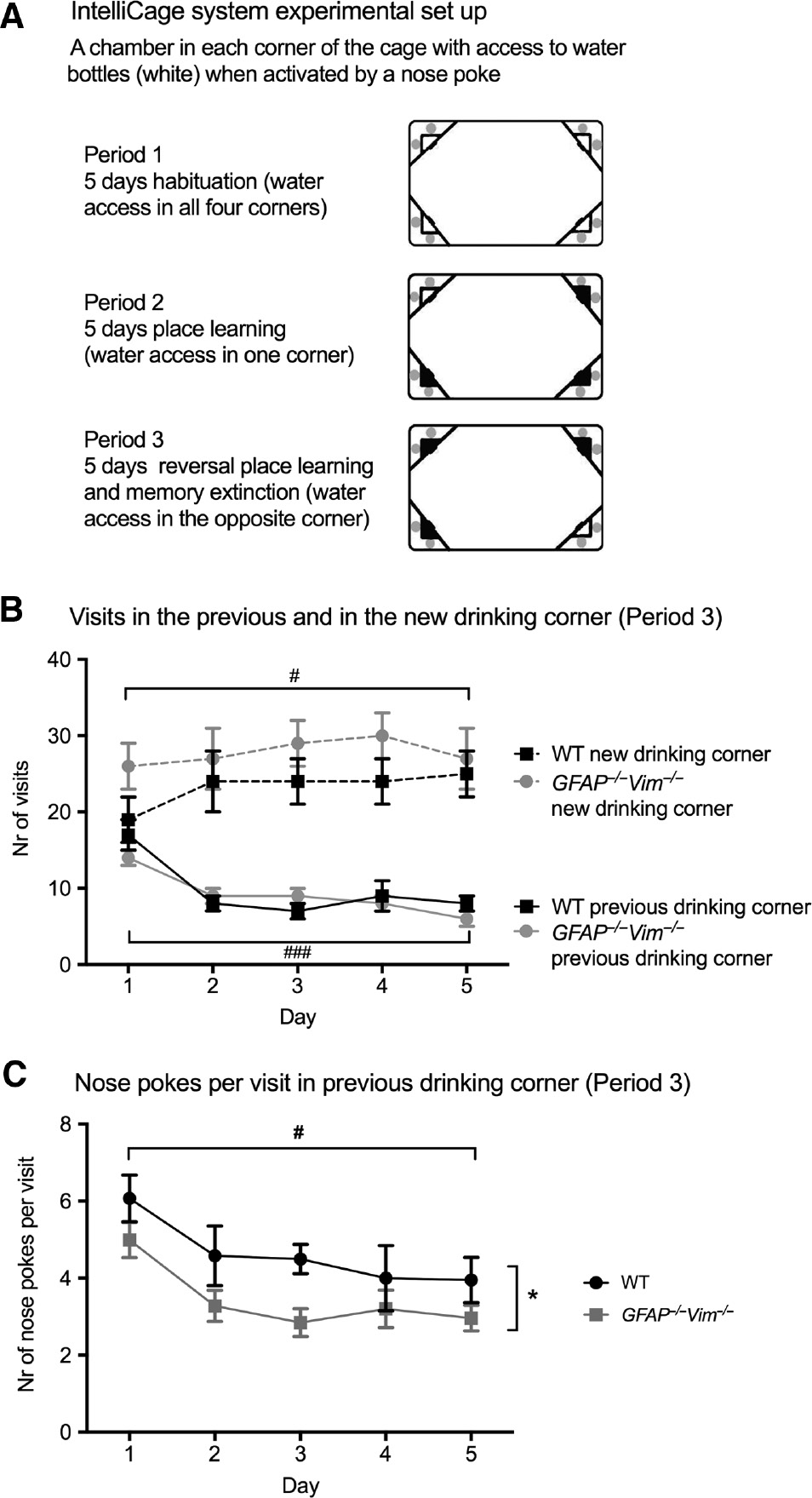
GFAP−/−Vim−/− mice show increased spatial memory extinction in IntelliCage housing.
Reversal place learning and memory extinction set up with IntelliCage system (A). Mice with microtransponders were housed in IntelliCages and habituated for 5 days (Period 1). Next, the water access after a nose poke was randomly limited to one of the four cage corners for 5 days (Period 2). For assessment of reversal place learning and memory extinction, each mouse was assigned a new drinking corner, and the number of visits and nose pokes in each corner was recorded for 5 days (Period 3). Wildtype (WT; n=12) and GFAP−/−Vim−/− mice (n=8) showed comparable drinking behavior assessed by the number of visits to the new drinking corner (B, dotted lines), and decreased number of visits to the previous drinking corner over time (B, continuous lines). The number of nose pokes per visit in the previous drinking corner was reduced for GFAP−/−Vim−/− compared to WT mice (C), indicating that GFAP−/−Vim−/− mice showed decreased persistence in trying to access water in the previous correct corner and thus exhibit more efficient extinction of place memory compared to WT mice. The generalized estimating equations statistical model was used to determine differences between groups and for the time factor. ###p<0.001; #p<0.05 for time factor (B and C); *p<0.05 for GFAP−/−Vim−/− vs. WT (C).
Discussion
Astrocytes regulate the micro-environment of the adult neurogenic niche (Song et al., 2002; Lu and Kipnis, 2010; Pekna et al., 2012; Pekny et al., 2016; Garber et al., 2018) and the synaptic integration of the newly formed neurons into existing hippocampal circuits (Tashiro et al., 2006; Toni et al., 2007) promotes the formation of new memories (Sahay et al., 2011; Anacker and Hen, 2017). The functional involvement of the newly generated neurons is suggested to lead to improved behavioral pattern separation, such as the ability to distinguish between different contexts or spatial cues, by modulating feedback inhibitory circuits in the hippocampus, so that fewer mature hippocampal neurons respond to incoming stimuli (Goncalves et al., 2016).
We report here that GFAP−/−Vim−/− mice which exhibit increased adult hippocampal neurogenesis (Larsson et al., 2004; Wilhelmsson et al., 2012) showed normal locomotor and exploratory behavior, which is in contrast to the previously reported increased locomotion of Vim−/− mice (Colucci-Guyon et al., 1999). GFAP−/−Vim−/− mice showed normal motor performance in the open field test and normal swimming speed in the Morris water maze. We did not find any alteration in exploratory behavior or anxiety in GFAP−/−Vim−/− mice, as evidenced by normal exploratory parameters assessed in the open field, or in the baseline freezing response in the novel context in the trace fear conditioning test. This supports the contention of an overall normal baseline behavior of GFAP−/−Vim−/− mice, which is a prerequisite for proper evaluation of their performance in other tests and in studies assessing functional recovery or disease progression.
We also report that GFAP−/−Vim−/− mice show normal short- and long-term memory retrieval in the object recognition test, spatial learning and memory retrieval in the Morris water maze test, and short- and long-term memory retrieval in contextual and trace fear conditioning tests. However, the GFAP−/−Vim−/− mice show increased memory extinction of place learning, indicating faster forgetting.
One possible explanation for the normal learning and memory retrieval in GFAP−/−Vim−/− mice, despite their increased hippocampal neurogenesis, could be that the tasks performed were not challenging enough with regards to requirements on pattern separation ability. Given that synaptic function is also critical for hippocampus-dependent cognitive functions (Dragoi et al., 2003; Nakazawa et al., 2003; Deng et al., 2010), altered synaptic plasticity and neurotransmission may present another explanation for the very subtle effect of GFAP−/−Vim−/− astrocytes on cognition. Notably, mice with reduced neurogenesis may exhibit enhanced place and reversal learning due to higher synaptic density (Perez-Alcazar et al., 2014; Shi et al., 2015). In addition, memory and forgetting processes are regulated at the level of NMDAR composition (Ge et al., 2019).
Recent studies show that astrocyte signaling on the synaptic as well as network level is linked to cognitive performance. For example, normal long-term memory formation is dependent on L-lactate shuttling from astrocytes to neurons (Suzuki et al., 2011), and mice with suppressed astrocyte exocytosis show impaired performance in the object recognition test (Lee et al., 2014). Indeed, astrocytes may contribute to normal cognitive function in several ways; through the regulation of neurogenesis (Wilhelmsson et al., 2012; Pekny and Pekna, 2016; Wilhelmsson et al., 2019) and the integration of new hippocampal neurons (Toni et al., 2007), and by affecting signal transmission, circuit formation, synaptic density and function (Santello et al., 2019). In support of this notion, astrocytes regulate synaptogenesis (Christopherson et al., 2005; Liauw et al., 2008; Eroglu et al., 2009; Kucukdereli et al., 2011; Stogsdill et al., 2017), synapse maturation (Allen et al., 2012) as well as synapse elimination (Chung et al., 2013). In addition, the release of D-serine by hippocampal astrocytes is required for NMDAR-dependent long-term potentiation (Henneberger et al., 2010), and supports the integration of new neurons into the hippocampal circuits (Sultan et al., 2015). Although increased levels of D-serine in mice did not affect the initial learning and memory retrieval in the Morris water maze task, they were associated with better reversal learning and faster memory extinction (Labrie et al., 2009). Similarly, our data support the contention that learning and memory duration are normal in GFAP−/−Vim−/− mice, but memory extinction is increased. Whereas synaptic density does not appear to be altered in GFAP−/−Vim−/− mice (Wilhelmsson et al., 2004), D-serine secretion by astrocytes in the hippocampus of these mice and the role of astrocytes in the developmental changes in the composition of NMDARs remain to be elucidated.
Interestingly, several studies indicate that the performance in spatial learning tasks is not impaired in mice in the absence of neurogenesis, but the same mice show impairment in reversal learning tasks (Kalm et al., 2013; Swan et al., 2014), suggesting that the newly born neurons might rather be involved in cognitive flexibility than spatial learning per se. This would fit with the model in which neurogenesis facilitates cognitive flexibility by allowing the formation of new distinct memory traces (Burghardt et al., 2012; Garthe et al., 2016; Anacker and Hen, 2017). Neurogenesis-mediated inhibition of hippocampal neurons may enable reversal learning by reducing memory interference through the extinction of previously learned associations (Epp et al., 2016). In this scenario, our data showing that GFAP−/−Vim−/− mice more readily forget previously acquired memories would be in line with the increased rate of hippocampal neurogenesis seen both in the adult and old GFAP−/−Vim−/− mice (Larsson et al., 2004; Wilhelmsson et al., 2012).
Materials and methods
Mice
Male mice 2–3 months old carrying a null mutation in GFAP (Pekny et al., 1995) and Vim (Colucci-Guyon et al., 1994) genes and wild-type control mice on the same genetic background (C57BL/6-129Sv-129Ola) were used for all the experiments and were housed in a barrier facility with a 12 h light/dark cycle with free access of food and water. A single colony of mice was used for all the experiments, heterozygotes were used for backcrosses, and the experimental groups were generated from littermates or within the next two generations. Experiments were conducted according to protocols approved by the Gothenburg Ethics Committee.
Open-field test
The activity of mice on the first day of habituation to the object recognition test arena was recorded as an open-field task to study locomotor and exploratory behavior as previously described (Wilhelmsson et al., 2019). A mouse was placed in the open-field and allowed to explore for 10 min and recorded. The Viewer video tracking system (Biobserve, Bonn, Germany) was used for the analysis.
Object recognition test
The object recognition test is used to assess declarative memory and is based on the preference of mice to spend more time with a novel object than a familiar one (Winters et al., 2008; Engelmann et al., 2011). The test was performed as previously described (Bevins and Besheer, 2006; Wilhelmsson et al., 2019). On each of 3 consecutive days, mice were habituated to the room for 60 min, then to the empty arena for 10 min. During the familiarization session, two identical objects (A and B) were placed 5 cm from the walls. Six hours later, short-term memory (STM) was assessed by replacing object B with a novel object C. Long-term memory (LTM) was assessed 24 h after by replacing the object C with a novel object D. Mice were allowed to explore the objects for 10 min per session and then returned to their cages. Exploration was defined as directing the nose to the object at a distance <2 cm or touching the object with the nose or forepaws. Mice that showed unequal exploration of the two identical objects, or failed to explore each object for at least 20 s during the familiarization session were excluded. Sessions were recorded and analyzed with a video tracking system (Viewer, Biobserve). Results are expressed as a discrimination ratio, calculated as the time spent exploring the novel object divided by the sum of the time spent exploring the novel and familiar object.
Morris water maze task
The Morris water maze task was done similarly as previously described (Dupret et al., 2008). The circular water maze had a diameter of 100 cm and a platform with a diameter of 10 cm was submerged 1 cm below the water surface in one of the quadrants. Temperature was constant at 18°C. The mice were trained for eight consecutive days with 5 daily trials (with 20 s rest between trials) starting from different positions, until they were able to find the platform within approximately 10–20 s. On the probe trial days 9 and 15, respectively, the platform was removed and all mice performed a single trial of 60 s. The latency to the first platform area crossing, the number of crossings and the time spent in the platform quadrant were recorded using a video-tracking system (2020 Plus tracking system, HVS Image, Hampton, UK).
Contextual and cued trace fear conditioning tests
Mice were tested for amygdala- and hippocampus-dependent associative cognition by contextual and cued trace fear conditioning as described (Curzon et al., 2009). The mice are trained to associate an aversive unconditioned stimulus (US; unpleasant electric impulses) with the conditioned stimulus (CS; the context or a cue). In trace fear conditioning, the CS and US are separated by stimulus-free “trace” interval. After training, the natural fear response (freezing of body movements) to the context or cue is measured.
Mice were habituated to the Automatic Reflex Conditioner (Ugo Basile, Gemonio, Italy), a rectangular plexiglass chamber with a metal grid floor in a sound-attenuating box to which a digital camera is attached to record the sessions. The mice were acclimatized to the testing room for 1 h, placed in the conditioning chamber, and allowed to explore it for 10 min. The next day, mice were placed in the same chamber and subjected to the conditioning cue (a 70 dB, 670 Hz tone) for 20 s, followed by a trace period of 18 s and a foot shock (0.5 mA, 2 s). This sequence was delivered 8 times at 60 s intervals. The mice were returned to their home cages, and the chamber was cleaned with 50% ethanol.
Contextual and cued fear responses were assessed 24 h after the conditioning session. During the contextual test, mice were placed in the conditioning chamber, their freezing behavior was recorded for 3 min, and the mice were returned to their home-cages for 1–3 h. The cued fear response was then assessed by placing the mice in a novel context with different properties (a plastic cylinder with striped walls and plastic floor, and novel odor of vanilla extract). After 3 min of exploration (pre-CS), the auditory cue was presented 4 times for 20 s at 60 s intervals (CS), and fear responses were recorded. The cued fear response was assessed again 2 months later in another novel context. Freezing behavior was defined as the absence of all body movements except those associated with respiration and were scored manually every 5th second throughout the contextual and cue tests. Freezing time was calculated as the percent of total time scored.
Place memory extinction in the IntelliCage test
Mice were tested for hippocampal-dependent reversal place learning and memory extinction using the IntelliCages (New Behavior, Zurich, Switzerland), a system for unbiased monitoring of mouse behavior in a home cage setting (Galsworthy et al., 2005; Karlsson et al., 2011). Two weeks prior to testing, mice were anesthetized with isoflurane (Abbott Laboratories, North Chicago, IL, USA) and implanted subcutaneously with microtransponders (DATAMARS, PetLink, Lamone, Switzerland) to allow individual animal identification in the IntelliCages. Mice were housed in the IntelliCages in groups of up to five animals per cage. The GFAP−/−Vim−/− (n=8) and WT (n=12) mice had a habituation period of 5 days in the IntelliCages during which the animals were acclimatized to performing nose pokes to gain access to the water bottles in all the four corners of the cage. During the 3 first days of the habituation period, all doors were open. During the last 2 days of the habituation the doors were closed, hence the mice had to practice doing a nose poke to open the door to access water. This was followed by a place learning period for which each animal was randomized to one drinking corner (the most visited corner during the habituation was excluded), in which a nose poke activates the door giving the mouse access to two water bottles. In the incorrect corners, the doors to the water bottles did not open in response to nose pokes. After 5 days, the animals were randomized to a new corner for the reversal learning and the number of visits and nose pokes for each corner were recorded. Animals were observed daily to ensure that the system registered drinking and visits. Food was provided ad libitum during the experiments in the IntelliCage and plastic houses were provided as shelters. Data from the IntelliCages were analyzed using the IntelliCage software (IntelliCage Plus, 2.4, New Behavior AG, Zurich, Switzerland) and SPSS 23.0 (SPSS, Chicago, IL, USA). Only the active (dark) period (16.00–08.00) was analyzed, visits not leading to a nose poke or visits lasting longer than 180 s were excluded from the analysis.
Statistical analysis
Statistical analyses were done with Microsoft Excel, SPSS 23.0 or GraphPad Prism 7.0 (GraphPad Software Inc., San Diego, CA, USA). Differences were considered significant at p<0.05. Values are presented as mean±standard error of the mean (SEM). Generalized estimating equations (GEE) in SPSS software were used to analyze the different variables measured in the IntelliCage system; this method has previously been described (Karlsson et al., 2011).
Acknowledgments
This work was supported by Swedish Medical Research Council (2017-02255), ALF Gothenburg (146051), AFA Research Foundation, Söderbergs Foundations, Sten A. Olsson Foundation for Research and Culture, Hjärnfonden, Hagströmer’s Foundation Millennium, Amlöv’s Foundation, E. Jacobson’s Donation Fund, VINNOVA, the Swedish Stroke Foundation, NanoNet COST Action (BM1002), EuroCellNet COST Action, EU FP 7 Program EduGlia (237956), and EU FP 7 Program TargetBraIn (279017). We thank Dr. Leke for teaching Dr. Pozo-Rodrigalvarez the open field and object recognition tests.
References
Akers, K.G., Martinez-Canabal, A., Restivo, L., Yiu, A.P., De Cristofaro, A., Hsiang, H.L., Wheeler, A.L., Guskjolen, A., Niibori, Y., Shoji, H., et al. (2014). Hippocampal neurogenesis regulates forgetting during adulthood and infancy. Science 344, 598–602.10.1126/science.1248903Search in Google Scholar
Allen, N.J., Bennett, M.L., Foo, L.C., Wang, G.X., Chakraborty, C., Smith, S.J., and Barres, B.A. (2012). Astrocyte glypicans 4 and 6 promote formation of excitatory synapses via GluA1 AMPA receptors. Nature 486, 410–414.10.1038/nature11059Search in Google Scholar
Anacker, C. and Hen, R. (2017). Adult hippocampal neurogenesis and cognitive flexibility – linking memory and mood. Nat. Rev. Neurosci. 18, 335–346.10.1038/nrn.2017.45Search in Google Scholar
Berg, A., Zelano, J., Pekna, M., Wilhelmsson, U., Pekny, M., and Cullheim, S. (2013). Axonal regeneration after sciatic nerve lesion is delayed but complete in GFAP- and vimentin-deficient mice. PLoS One 8, e79395.10.1371/journal.pone.0079395Search in Google Scholar
Bevins, R.A. and Besheer, J. (2006). Object recognition in rats and mice: a one-trial non-matching-to-sample learning task to study ‘recognition memory’. Nat. Protoc. 1, 1306–1311.10.1038/nprot.2006.205Search in Google Scholar
Burghardt, N.S., Park, E.H., Hen, R., and Fenton, A.A. (2012). Adult-born hippocampal neurons promote cognitive flexibility in mice. Hippocampus 22, 1795–1808.10.1002/hipo.22013Search in Google Scholar
Cho, K.S., Yang, L., Lu, B., Feng, M.H., Huang, X., Pekny, M., and Chen, D.F. (2005). Re-establishing the regenerative potential of central nervous system axons in postnatal mice. J. Cell Sci. 118, 863–872.10.1242/jcs.01658Search in Google Scholar
Christopherson, K.S., Ullian, E.M., Stokes, C.C., Mullowney, C.E., Hell, J.W., Agah, A., Lawler, J., Mosher, D.F., Bornstein, P., and Barres, B.A. (2005). Thrombospondins are astrocyte-secreted proteins that promote CNS synaptogenesis. Cell 120, 421–433.10.1016/j.cell.2004.12.020Search in Google Scholar
Chung, W.S., Clarke, L.E., Wang, G.X., Stafford, B.K., Sher, A., Chakraborty, C., Joung, J., Foo, L.C., Thompson, A., Chen, C., et al. (2013). Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 504, 394–400.10.1038/nature12776Search in Google Scholar
Colucci-Guyon, E., Portier, M.M., Dunia, I., Paulin, D., Pournin, S., and Babinet, C. (1994). Mice lacking vimentin develop and reproduce without an obvious phenotype. Cell 79, 679–694.10.1016/0092-8674(94)90553-3Search in Google Scholar
Colucci-Guyon, E., Gimenez, Y.R.M., Maurice, T., Babinet, C., and Privat, A. (1999). Cerebellar defect and impaired motor coordination in mice lacking vimentin. Glia 25, 33–43.10.1002/(SICI)1098-1136(19990101)25:1<33::AID-GLIA4>3.0.CO;2-JSearch in Google Scholar
Curzon, P., Rustay, N.R., and Browman, K.E. (2009). Cued and contextual fear conditioning for rodents. Methods Behav Anal Neurosci. JJ Buccafusco. Boca Raton (FL).10.1201/NOE1420052343.ch2Search in Google Scholar
de Pablo, Y., Nilsson, M., Pekna, M., and Pekny, M. (2013). Intermediate filaments are important for astrocyte response to oxidative stress induced by oxygen-glucose deprivation and reperfusion. Histochem. Cell Biol. 140, 81–91.10.1007/s00418-013-1110-0Search in Google Scholar
Deng, W., Aimone, J.B., and Gage, F.H. (2010). New neurons and new memories: how does adult hippocampal neurogenesis affect learning and memory? Nat. Rev. Neurosci. 11, 339–350.10.1038/nrn2822Search in Google Scholar
Dragoi, G., Harris, K.D., and Buzsaki, G. (2003). Place representation within hippocampal networks is modified by long-term potentiation. Neuron 39, 843–853.10.1016/S0896-6273(03)00465-3Search in Google Scholar
Dupret, D., Revest, J.M., Koehl, M., Ichas, F., De Giorgi, F., Costet, P., Abrous, D.N., and Piazza, P.V. (2008). Spatial relational memory requires hippocampal adult neurogenesis. PLoS One 3, e1959.10.1371/journal.pone.0001959Search in Google Scholar
Eliasson, C., Sahlgren, C., Berthold, C.H., Stakeberg, J., Celis, J.E., Betsholtz, C., Eriksson, J.E., and Pekny, M. (1999). Intermediate filament protein partnership in astrocytes. J. Biol. Chem. 274, 23996–24006.10.1074/jbc.274.34.23996Search in Google Scholar
Engelmann, M., Hadicke, J., and Noack, J. (2011). Testing declarative memory in laboratory rats and mice using the nonconditioned social discrimination procedure. Nat. Protoc. 6, 1152–1162.10.1038/nprot.2011.353Search in Google Scholar
Epp, J.R., Silva Mera, R., Kohler, S., Josselyn, S.A., and Frankland, P.W. (2016). Neurogenesis-mediated forgetting minimizes proactive interference. Nat. Commun. 7, 10838.10.1038/ncomms10838Search in Google Scholar
Eroglu, C. and Barres, B.A. (2010). Regulation of synaptic connectivity by glia. Nature 468, 223–231.10.1038/nature09612Search in Google Scholar
Eroglu, C., Allen, N.J., Susman, M.W., O’Rourke, N.A., Park, C.Y., Ozkan, E., Chakraborty, C., Mulinyawe, S.B., Annis, D.S., Huberman, A.D., et al. (2009). Gabapentin receptor a2d-1 is a neuronal thrombospondin receptor responsible for excitatory CNS synaptogenesis. Cell 139, 380–392.10.1016/j.cell.2009.09.025Search in Google Scholar
Galsworthy, M.J., Amrein, I., Kuptsov, P.A., Poletaeva, II, Zinn, P., Rau, A., Vyssotski, A., and Lipp, H.P. (2005). A comparison of wild-caught wood mice and bank voles in the Intellicage: assessing exploration, daily activity patterns and place learning paradigms. Behav. Brain Res. 157, 211–217.10.1016/j.bbr.2004.06.021Search in Google Scholar PubMed
Garber, C., Vasek, M.J., Vollmer, L.L., Sun, T., Jiang, X., and Klein, R.S. (2018). Astrocytes decrease adult neurogenesis during virus-induced memory dysfunction via IL-1. Nat. Immunol. 19, 151–161.10.1038/s41590-017-0021-ySearch in Google Scholar PubMed PubMed Central
Garthe, A., Roeder, I., and Kempermann, G. (2016). Mice in an enriched environment learn more flexibly because of adult hippocampal neurogenesis. Hippocampus 26, 261–271.10.1002/hipo.22520Search in Google Scholar PubMed PubMed Central
Ge, M., Song, H., Li, H., Li, R., Tao, X., Zhan, X., Yu, N., Sun, N., Lu, Y., and Mu, Y. (2019). Memory susceptibility to retroactive interference is developmentally regulated by NMDA receptors. Cell Rep. 26, 2052–2063 e2054.10.1016/j.celrep.2019.01.098Search in Google Scholar PubMed
Goncalves, J.T., Schafer, S.T., and Gage, F.H. (2016). Adult neurogenesis in the hippocampus: from stem cells to behavior. Cell 167, 897–914.10.1016/j.cell.2016.10.021Search in Google Scholar PubMed
Henneberger, C., Papouin, T., Oliet, S.H., and Rusakov, D.A. (2010). Long-term potentiation depends on release of D-serine from astrocytes. Nature 463, 232–236.10.1038/nature08673Search in Google Scholar PubMed PubMed Central
Jarlestedt, K., Rousset, C.I., Faiz, M., Wilhelmsson, U., Stahlberg, A., Sourkova, H., Pekna, M., Mallard, C., Hagberg, H., and Pekny, M. (2010). Attenuation of reactive gliosis does not affect infarct volume in neonatal hypoxic-ischemic brain injury in mice. PLoS One 5, e10397.10.1371/journal.pone.0010397Search in Google Scholar PubMed PubMed Central
Kalm, M., Karlsson, N., Nilsson, M.K., and Blomgren, K. (2013). Loss of hippocampal neurogenesis, increased novelty-induced activity, decreased home cage activity, and impaired reversal learning one year after irradiation of the young mouse brain. Exp. Neurol. 247, 402–409.10.1016/j.expneurol.2013.01.006Search in Google Scholar PubMed
Kamphuis, W., Kooijman, L., Orre, M., Stassen, O., Pekny, M., and Hol, E.M. (2015). GFAP and vimentin deficiency alters gene expression in astrocytes and microglia in wild-type mice and changes the transcriptional response of reactive glia in mouse model for Alzheimer’s disease. Glia 63, 1036–1056.10.1002/glia.22800Search in Google Scholar PubMed
Karlsson, N., Kalm, M., Nilsson, M.K., Mallard, C., Bjork-Eriksson, T., and Blomgren, K. (2011). Learning and activity after irradiation of the young mouse brain analyzed in adulthood using unbiased monitoring in a home cage environment. Radiat. Res. 175, 336–346.10.1667/RR2231.1Search in Google Scholar PubMed
Kinouchi, R., Takeda, M., Yang, L., Wilhelmsson, U., Lundkvist, A., Pekny, M., and Chen, D.F. (2003). Robust neural integration from retinal transplants in mice deficient in GFAP and vimentin. Nat. Neurosci. 6, 863–868.10.1038/nn1088Search in Google Scholar PubMed
Kraft, A.W., Hu, X., Yoon, H., Yan, P., Xiao, Q., Wang, Y., Gil, S.C., Brown, J., Wilhelmsson, U., Restivo, J.L., et al. (2013). Attenuating astrocyte activation accelerates plaque pathogenesis in APP/PS1 mice. FASEB J. 27, 187–198.10.1096/fj.12-208660Search in Google Scholar PubMed PubMed Central
Kucukdereli, H., Allen, N.J., Lee, A.T., Feng, A., Ozlu, M.I., Conatser, L.M., Chakraborty, C., Workman, G., Weaver, M., Sage, E.H., et al. (2011). Control of excitatory CNS synaptogenesis by astrocyte-secreted proteins Hevin and SPARC. Proc. Natl. Acad. Sci. USA 108, E440–449.10.1073/pnas.1104977108Search in Google Scholar PubMed PubMed Central
Labrie, V., Duffy, S., Wang, W., Barger, S.W., Baker, G.B., and Roder, J.C. (2009). Genetic inactivation of D-amino acid oxidase enhances extinction and reversal learning in mice. Learn. Mem. 16, 28–37.10.1101/lm.1112209Search in Google Scholar PubMed PubMed Central
Laplagne, D.A., Esposito, M.S., Piatti, V.C., Morgenstern, N.A., Zhao, C., van Praag, H., Gage, F.H., and Schinder, A.F. (2006). Functional convergence of neurons generated in the developing and adult hippocampus. PLoS Biol. 4, e409.10.1371/journal.pbio.0040409Search in Google Scholar PubMed PubMed Central
Larsson, A., Wilhelmsson, U., Pekna, M., and Pekny, M. (2004). Increased cell proliferation and neurogenesis in the hippocampal dentate gyrus of old GFAP−/−Vim−/− mice. Neurochem. Res. 29, 2069–2073.10.1007/s11064-004-6880-2Search in Google Scholar PubMed
Laterza, C., Uoshima, N., Tornero, D., Wilhelmsson, U., Stokowska, A., Ge, R., Pekny, M., Lindvall, O., and Kokaia, Z. (2018). Attenuation of reactive gliosis in stroke-injured mouse brain does not affect neurogenesis from grafted human iPSC-derived neural progenitors. PLoS One 13, e0192118.10.1371/journal.pone.0192118Search in Google Scholar PubMed PubMed Central
Lee, H.S., Ghetti, A., Pinto-Duarte, A., Wang, X., Dziewczapolski, G., Galimi, F., Huitron-Resendiz, S., Pina-Crespo, J.C., Roberts, A.J., Verma, I.M., et al. (2014). Astrocytes contribute to gamma oscillations and recognition memory. Proc. Natl. Acad. Sci. USA 111, E3343–3352.10.1073/pnas.1410893111Search in Google Scholar PubMed PubMed Central
Li, L., Lundkvist, A., Andersson, D., Wilhelmsson, U., Nagai, N., Pardo, A.C., Nodin, C., Stahlberg, A., Aprico, K., Larsson, K., et al. (2008). Protective role of reactive astrocytes in brain ischemia. J. Cereb. Blood Flow Metab. 28, 468–481.10.1038/sj.jcbfm.9600546Search in Google Scholar PubMed
Liauw, J., Hoang, S., Choi, M., Eroglu, C., Choi, M., Sun, G.H., Percy, M., Wildman-Tobriner, B., Bliss, T., Guzman, R.G., et al. (2008). Thrombospondins 1 and 2 are necessary for synaptic plasticity and functional recovery after stroke. J. Cereb. Blood Flow Metab. 28, 1722–1732.10.1038/jcbfm.2008.65Search in Google Scholar PubMed
Liu, Z., Li, Y., Cui, Y., Roberts, C., Lu, M., Wilhelmsson, U., Pekny, M., and Chopp, M. (2014). Beneficial effects of gfap/vimentin reactive astrocytes for axonal remodeling and motor behavioral recovery in mice after stroke. Glia 62, 2022–2033.10.1002/glia.22723Search in Google Scholar
Lu, Z. and Kipnis, J. (2010). Thrombospondin 1--a key astrocyte-derived neurogenic factor. FASEB J. 24, 1925–1934.10.1096/fj.09-150573Search in Google Scholar
Lu, Y.B., Iandiev, I., Hollborn, M., Korber, N., Ulbricht, E., Hirrlinger, P.G., Pannicke, T., Wie, E.Q., Bringmann, A., Wolburg, H., et al. (2011). Reactive glial cells: increased stiffness correlates with increased intermediate filament expression. FASEB J. 25, 624–631.10.1096/fj.10-163790Search in Google Scholar
Lundkvist, A., Reichenbach, A., Betsholtz, C., Carmeliet, P., Wolburg, H., and Pekny, M. (2004). Under stress, the absence of intermediate filaments from Muller cells in the retina has structural and functional consequences. J. Cell Sci. 117, 3481–3488.10.1242/jcs.01221Search in Google Scholar
Macauley, S.L., Pekny, M., and Sands, M.S. (2011). The role of attenuated astrocyte activation in infantile neuronal ceroid lipofuscinosis. J. Neurosci. 31, 15575–15585.10.1523/JNEUROSCI.3579-11.2011Search in Google Scholar
Nakazawa, K., Sun, L.D., Quirk, M.C., Rondi-Reig, L., Wilson, M.A., and Tonegawa, S. (2003). Hippocampal CA3 NMDA receptors are crucial for memory acquisition of one-time experience. Neuron 38, 305–315.10.1016/S0896-6273(03)00165-XSearch in Google Scholar
Nakazawa, T., Takeda, M., Lewis, G.P., Cho, K.S., Jiao, J., Wilhelmsson, U., Fisher, S.K., Pekny, M., Chen, D.F., and Miller, J.W. (2007). Attenuated glial reactions and photoreceptor degeneration after retinal detachment in mice deficient in glial fibrillary acidic protein and vimentin. Invest. Ophthalmol. Vis. Sci. 48, 2760–2768.10.1167/iovs.06-1398Search in Google Scholar PubMed PubMed Central
Pekna, M., Pekny, M., and Nilsson, M. (2012). Modulation of neural plasticity as a basis for stroke rehabilitation. Stroke 43, 2819–2828.10.1161/STROKEAHA.112.654228Search in Google Scholar PubMed
Pekny, M. and Lane, E.B. (2007). Intermediate filaments and stress. Exp. Cell Res. 313, 2244–2254.10.1016/j.yexcr.2007.04.023Search in Google Scholar PubMed
Pekny, M. and Pekna, M. (2014). Astrocyte reactivity and reactive astrogliosis: costs and benefits. Physiol. Rev. 94, 1077–1098.10.1152/physrev.00041.2013Search in Google Scholar PubMed
Pekny, M. and Pekna, M. (2016). Reactive gliosis in the pathogenesis of CNS diseases. Biochim. Biophys. Acta 1862, 483–491.10.1016/j.bbadis.2015.11.014Search in Google Scholar PubMed
Pekny, M., Leveen, P., Pekna, M., Eliasson, C., Berthold, C.H., Westermark, B., and Betsholtz, C. (1995). Mice lacking glial fibrillary acidic protein display astrocytes devoid of intermediate filaments but develop and reproduce normally. EMBO J. 14, 1590–1598.10.1002/j.1460-2075.1995.tb07147.xSearch in Google Scholar
Pekny, M., Johansson, C.B., Eliasson, C., Stakeberg, J., Wallen, A., Perlmann, T., Lendahl, U., Betsholtz, C., Berthold, C.H., and Frisen, J. (1999). Abnormal reaction to central nervous system injury in mice lacking glial fibrillary acidic protein and vimentin. J. Cell Biol. 145, 503–514.10.1083/jcb.145.3.503Search in Google Scholar PubMed PubMed Central
Pekny, M., Wilhelmsson, U., and Pekna, M. (2014). The dual role of astrocyte activation and reactive gliosis. Neurosci. Lett. 565, 30–38.10.1016/j.neulet.2013.12.071Search in Google Scholar PubMed
Pekny, M., Pekna, M., Messing, A., Steinhauser, C., Lee, J.M., Parpura, V., Hol, E.M., Sofroniew, M.V., and Verkhratsky, A. (2016). Astrocytes: a central element in neurological diseases. Acta Neuropathol. 131, 323–345.10.1007/s00401-015-1513-1Search in Google Scholar PubMed
Pekny, M., Wilhelmsson, U., Tatlisumak, T., and Pekna, M. (2019). Astrocyte activation and reactive gliosis-a new target in stroke? Neurosci. Lett. 689, 45–55.10.1016/j.neulet.2018.07.021Search in Google Scholar PubMed
Perez-Alcazar, M., Daborg, J., Stokowska, A., Wasling, P., Bjorefeldt, A., Kalm, M., Zetterberg, H., Carlstrom, K.E., Blomgren, K., Ekdahl, C.T., et al. (2014). Altered cognitive performance and synaptic function in the hippocampus of mice lacking C3. Exp. Neurol. 253, 154–164.10.1016/j.expneurol.2013.12.013Search in Google Scholar PubMed
Sahay, A., Scobie, K.N., Hill, A.S., O’Carroll, C.M., Kheirbek, M.A., Burghardt, N.S., Fenton, A.A., Dranovsky, A., and Hen, R. (2011). Increasing adult hippocampal neurogenesis is sufficient to improve pattern separation. Nature 472, 466–470.10.1038/nature09817Search in Google Scholar PubMed PubMed Central
Santello, M., Toni, N., and Volterra, A. (2019). Astrocyte function from information processing to cognition and cognitive impairment. Nat. Neurosci. 22, 154–166.10.1038/s41593-018-0325-8Search in Google Scholar PubMed
Shi, Q., Colodner, K.J., Matousek, S.B., Merry, K., Hong, S., Kenison, J.E., Frost, J.L., Le, K.X., Li, S., Dodart, J.C., et al. (2015). Complement C3-deficient mice fail to display age-related hippocampal decline. J. Neurosci. 35, 13029–13042.10.1523/JNEUROSCI.1698-15.2015Search in Google Scholar PubMed PubMed Central
Song, H., Stevens, C.F., and Gage, F.H. (2002). Astroglia induce neurogenesis from adult neural stem cells. Nature 417, 39–44.10.1038/417039aSearch in Google Scholar PubMed
Stogsdill, J.A., Ramirez, J., Liu, D., Kim, Y.H., Baldwin, K.T., Enustun, E., Ejikeme, T., Ji, R.R., and Eroglu, C. (2017). Astrocytic neuroligins control astrocyte morphogenesis and synaptogenesis. Nature 551, 192–197.10.1038/nature24638Search in Google Scholar PubMed PubMed Central
Sultan, S., Li, L., Moss, J., Petrelli, F., Casse, F., Gebara, E., Lopatar, J., Pfrieger, F.W., Bezzi, P., Bischofberger, J., et al. (2015). Synaptic integration of adult-born hippocampal neurons is locally controlled by astrocytes. Neuron 88, 957–972.10.1016/j.neuron.2015.10.037Search in Google Scholar PubMed
Suzuki, A., Stern, S.A., Bozdagi, O., Huntley, G.W., Walker, R.H., Magistretti, P.J., and Alberini, C.M. (2011). Astrocyte-neuron lactate transport is required for long-term memory formation. Cell 144, 810–823.10.1016/j.cell.2011.02.018Search in Google Scholar PubMed PubMed Central
Swan, A.A., Clutton, J.E., Chary, P.K., Cook, S.G., Liu, G.G., and Drew, M.R. (2014). Characterization of the role of adult neurogenesis in touch-screen discrimination learning. Hippocampus 24, 1581–1591.10.1002/hipo.22337Search in Google Scholar PubMed PubMed Central
Tashiro, A., Sandler, V.M., Toni, N., Zhao, C., and Gage, F.H. (2006). NMDA-receptor-mediated, cell-specific integration of new neurons in adult dentate gyrus. Nature 442, 929–933.10.1038/nature05028Search in Google Scholar PubMed
Toni, N., Teng, E.M., Bushong, E.A., Aimone, J.B., Zhao, C., Consiglio, A., van Praag, H., Martone, M.E., Ellisman, M.H., and Gage, F.H. (2007). Synapse formation on neurons born in the adult hippocampus. Nat. Neurosci. 10, 727–734.10.1038/nn1908Search in Google Scholar PubMed
Verardo, M.R., Lewis, G.P., Takeda, M., Linberg, K.A., Byun, J., Luna, G., Wilhelmsson, U., Pekny, M., Chen, D.F., and Fisher, S.K. (2008). Abnormal reactivity of muller cells after retinal detachment in mice deficient in GFAP and vimentin. Invest. Ophthalmol. Vis. Sci. 49, 3659–3665.10.1167/iovs.07-1474Search in Google Scholar PubMed PubMed Central
Verkhratsky, A. and Nedergaard, M. (2018). Physiology of astroglia. Physiol. Rev. 98, 239–389.10.1007/978-981-13-9913-8_3Search in Google Scholar PubMed PubMed Central
Widestrand, A., Faijerson, J., Wilhelmsson, U., Smith, P.L., Li, L., Sihlbom, C., Eriksson, P.S., and Pekny, M. (2007). Increased neurogenesis and astrogenesis from neural progenitor cells grafted in the hippocampus of GFAP−/− Vim−/− mice. Stem Cells 25, 2619–2627.10.1634/stemcells.2007-0122Search in Google Scholar PubMed
Wilhelmsson, U., Li, L., Pekna, M., Berthold, C.H., Blom, S., Eliasson, C., Renner, O., Bushong, E., Ellisman, M., Morgan, T.E., et al. (2004). Absence of glial fibrillary acidic protein and vimentin prevents hypertrophy of astrocytic processes and improves post-traumatic regeneration. J. Neurosci. 24, 5016–5021.10.1523/JNEUROSCI.0820-04.2004Search in Google Scholar PubMed PubMed Central
Wilhelmsson, U., Faiz, M., de Pablo, Y., Sjoqvist, M., Andersson, D., Widestrand, A., Potokar, M., Stenovec, M., Smith, P.L., Shinjyo, N., et al. (2012). Astrocytes negatively regulate neurogenesis through the Jagged1-mediated Notch pathway. Stem Cells 30, 2320–2329.10.1002/stem.1196Search in Google Scholar PubMed
Wilhelmsson, U., Lebkuechner, I., Leke, R., Marasek, P., Yang, X., Antfolk, A., Chen, M., Mohseni, P., Lasic, E., Bobnar, S., et al. (2019). Nestin regulates neurogenesis in mice through Notch signaling from astrocytes to neural stem cells. Cereb. Cortex, doi: 10.1093/cercor/bhy284.10.1093/cercor/bhy284Search in Google Scholar PubMed
Winters, B.D., Saksida, L.M., and Bussey, T.J. (2008). Object recognition memory: neurobiological mechanisms of encoding, consolidation and retrieval. Neurosci. Biobehav. Rev. 32, 1055–1070.10.1016/j.neubiorev.2008.04.004Search in Google Scholar PubMed
Wunderlich, K.A., Tanimoto, N., Grosche, A., Zrenner, E., Pekny, M., Reichenbach, A., Seeliger, M.W., Pannicke, T., and Perez, M.T. (2015). Retinal functional alterations in mice lacking intermediate filament proteins glial fibrillary acidic protein and vimentin. FASEB J. 29, 4815–4828.10.1096/fj.15-272963Search in Google Scholar PubMed
©2019 Ulrika Wilhelmsson et al., published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 Public License.
Articles in the same Issue
- Frontmatter
- Highlight: dynamics of the nervous system in health and disease
- Actin(g) on mitochondria – a role for cofilin1 in neuronal cell death pathways
- Neuronal microtubules and proteins linked to Parkinson’s disease: a relevant interaction?
- Contribution of astrocytes to metabolic dysfunction in the Alzheimer’s disease brain
- Chemical LTD, but not LTP, induces transient accumulation of gelsolin in dendritic spines
- Sub-membranous actin rings in the axon initial segment are resistant to the action of latrunculin
- The role of GFAP and vimentin in learning and memory
- Vimentin is required for normal accumulation of body fat
- The microtubule skeleton and the evolution of neuronal complexity in vertebrates
- Amyloidosis causes downregulation of SorLA, SorCS1 and SorCS3 expression in mice
- A fluorescent protein-readout for transcriptional activity reveals regulation of APP nuclear signaling by phosphorylation sites
- Kavalactones from Kava (Piper methysticum) root extract as modulators of recombinant human glycine receptors
- Research Articles/Short Communications
- Cell Biology and Signaling
- LncRNA HOTAIR targets miR-126-5p to promote the progression of Parkinson’s disease through RAB3IP
- Novel Techniques
- In vitro import experiments with semi-intact cells suggest a role of the Sec61 paralog Ssh1 in mitochondrial biogenesis
- Corrigendum
- Corrigendum to: SIAH1/ZEB1/IL-6 axis is involved in doxorubicin (Dox) resistance of osteosarcoma cells
Articles in the same Issue
- Frontmatter
- Highlight: dynamics of the nervous system in health and disease
- Actin(g) on mitochondria – a role for cofilin1 in neuronal cell death pathways
- Neuronal microtubules and proteins linked to Parkinson’s disease: a relevant interaction?
- Contribution of astrocytes to metabolic dysfunction in the Alzheimer’s disease brain
- Chemical LTD, but not LTP, induces transient accumulation of gelsolin in dendritic spines
- Sub-membranous actin rings in the axon initial segment are resistant to the action of latrunculin
- The role of GFAP and vimentin in learning and memory
- Vimentin is required for normal accumulation of body fat
- The microtubule skeleton and the evolution of neuronal complexity in vertebrates
- Amyloidosis causes downregulation of SorLA, SorCS1 and SorCS3 expression in mice
- A fluorescent protein-readout for transcriptional activity reveals regulation of APP nuclear signaling by phosphorylation sites
- Kavalactones from Kava (Piper methysticum) root extract as modulators of recombinant human glycine receptors
- Research Articles/Short Communications
- Cell Biology and Signaling
- LncRNA HOTAIR targets miR-126-5p to promote the progression of Parkinson’s disease through RAB3IP
- Novel Techniques
- In vitro import experiments with semi-intact cells suggest a role of the Sec61 paralog Ssh1 in mitochondrial biogenesis
- Corrigendum
- Corrigendum to: SIAH1/ZEB1/IL-6 axis is involved in doxorubicin (Dox) resistance of osteosarcoma cells