Mitophagy could fight Parkinson’s disease through antioxidant action
-
Anthea Di Rita
Anthea Di Rita graduated with a degree in Cellular and Molecular Biology in 2014 at the University of Rome Tor Vergata (Italy). During her PhD, she acquired a solid knowledge in the fields of autophagy, mitophagy, and cell death. She is currently a post-doctoral fellow working under the supervision of Dr. Flavie Strappazzon at the IRCCS Fondazione Santa Lucia in Rome (Italy).
Flavie Strappazzon is a junior head of a laboratory at the IRCCS Fondazione Santa Lucia in Rome (Italy) and an external teacher at the University of Rome "Tor Vergata". She graduated with a degree in Cellular Biology in 2007 at the University of Grenoble (France). During this period, she acquired a solid knowledge in Neurosciences. Next, she moved to Italy to perform two post-doctoral trainings in the field of Autophagy. In 2018, she started her research group working on selective autophagy and, in particular, on mitophagy in the context of multiple sclerosis and Huntington’s disease (IRCCS Santa Lucia, Rome).


Abstract
During aging, the process of mitophagy, a system that allows the removal of dysfunctional mitochondria through lysosomal degradation, starts to malfunction. Because of this defect, damaged mitochondria are not removed correctly, and their decomposing components accumulate inside the cells. Dysfunctional mitochondria that are not removed by mitophagy produce high amounts of reactive oxygen species (ROS) and, thus, cause oxidative stress. Oxidative stress, in turn, is very harmful for the cells, neuronal cells, in particular. Consequently, the process of mitophagy plays a crucial role in mitochondria-related disease. Mitochondrial dysfunctions and oxidative stress are well-established factors contributing to Parkinson’s disease (PD), one of the most common neurodegenerative disorders. In this review, we report various known antioxidants for PD treatments and describe the stimulation of mitophagy process as a novel and exciting method for reducing oxidative stress in PD patients. We describe the different mechanisms responsible for mitochondria removal through the mitophagy process. In addition, we review the functional connection between mitophagy induction and reduction of oxidative stress in several in vitro models of PD and also agents (drugs and natural compounds) already known to be antioxidants and to be able to activate mitophagy. Finally, we propose that there is an urgent need to test the use of mitophagy-inducing antioxidants in order to fight PD.
Parkinson’s disease and mitochondrial oxidative stress
Parkinson’s disease (PD) is a severe and widespread neurodegenerative disorder characterized by a progressive reduction of neurons in the substantia nigra pars compacta leading to dopamine deficiency and α-synuclein aggregate formation. This pathology is associated with motor and non-motor symptoms, including bradykinesia, rigidity, dystonia, anxiety, dementia, and sexual dysfunction. The disease stems from the impairment of several molecular pathways, including α-synuclein proteostasis, mitochondrial function, oxidative stress, calcium homeostasis, axonal transport, and neuroinflammation (Poewe et al., 2017).
Although the etiology of PD is still elusive, mitochondrial dysfunctions and related oxidative stress are crucial contributing factors (Figure 1). Several findings suggest that mitochondria are the primary source of reactive oxygen species (ROS) that are possibly contributing to intracellular oxidative stress (Starkov, 2008; Murphy, 2009). Various groups observed a reduction in mitochondrial complex I protein level or activity in the nigral neurons of PD patients (Schapira et al., 1990; Bose and Beal 2016). Inclusions of α-synuclein protein aggregates in the mitochondria induce an impairment in the mitochondrial complex I activity leading to oxidative stress (Devi et al., 2008). Moreover, it was demonstrated that the downregulation in dopaminergic neurons of the mitochondrial transcription factor A (TFAM), essential for mitochondrial DNA expression, leads to a defect in the electron transport chain, thus, promoting neuronal degeneration in adulthood (Ekstrand et al., 2007). In addition, some mitochondria-related genes, i.e. phosphatase and tensin homolog-induced kinase 1 (PINK1), parkin RING-in-between-RING E3 ubiquitin protein ligase (PARK2/PRKN), Parkinsonism-associated deglycase (DJ-1/PARK7), and F-box protein 7 (Fbxo7), prove to be mutated in the familial form of PD. These variations compromise mitochondria functioning and contribute to oxidative stress production. Furthermore, PD patients show high levels (40–60%) of mtDNA deletions, associated with a cytochrome c oxidase deficiency in the substantia nigra (Bender et al., 2006; Kraytsberg et al., 2006). Isobe and colleagues showed that the levels of oxidized coenzyme Q-10 (CoQ10) in PD patients’ cerebrospinal fluid were significantly increased (Isobe et al., 2010). Other pathologies, such as Alzheimer’s disease (AD), multiple sclerosis (MS), or amyotrophic lateral sclerosis (ALS) are related to damaged mitochondria and oxidative stress production (Dutta et al., 2006; Sau et al., 2007; Campbell et al., 2011; Swerdlow et al., 2014). To date, pharmacological substitution of striatal dopamine, and more recently, therapies for cellular transport of α-synuclein remain the principal treatment for PD.
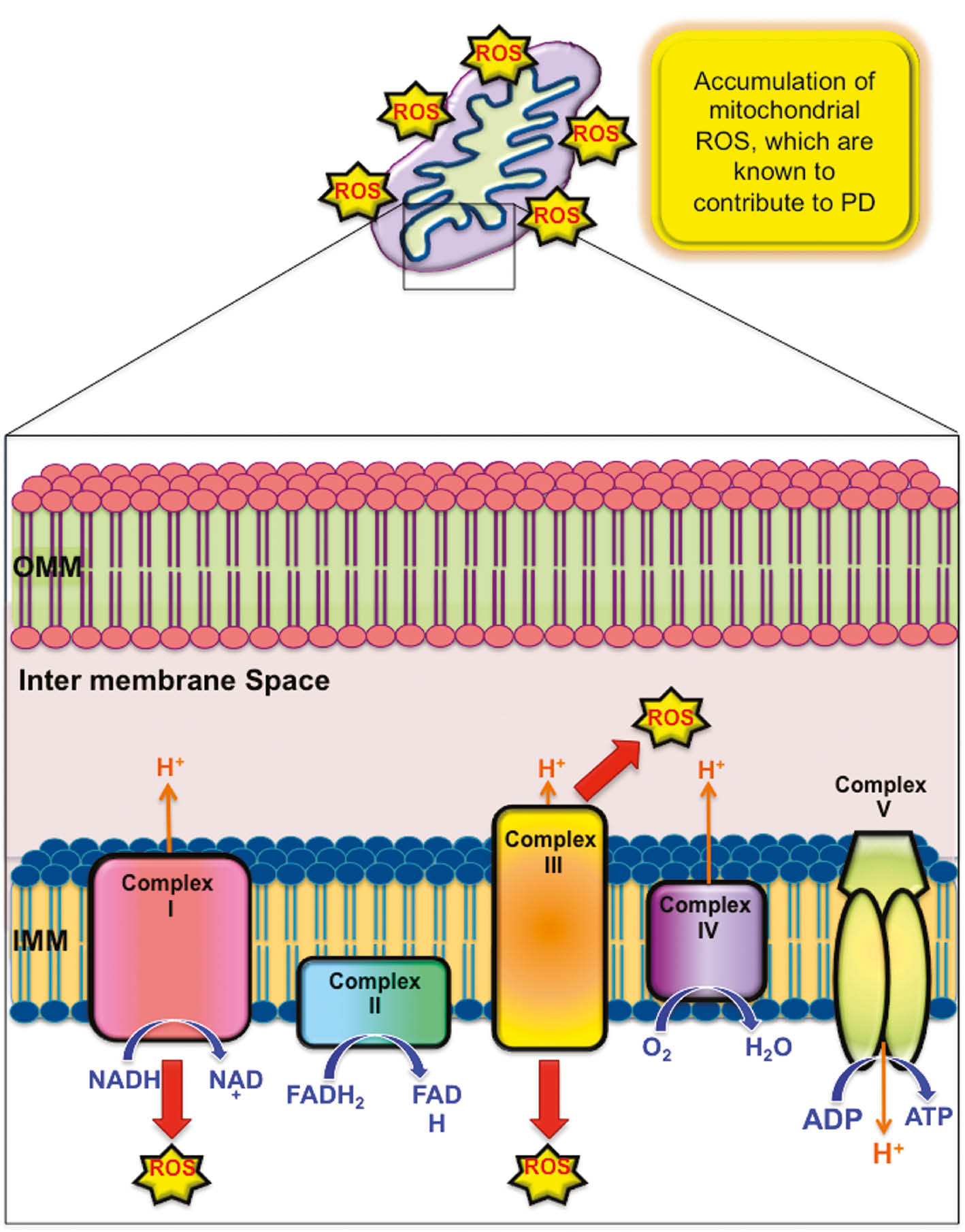
Generation of mitochondrial ROS contributes to PD.
A defect in the complex I or complex III of mitochondrial respiratory chain promotes the accumulation of reactive radicals, leading to oxidative stress.
Given the link between mitochondrial dysfunctions and ROS production in PD, clinical approaches that counteract the oxidative stress may be beneficial for patients. We review below some of the main promising antioxidants for improving PD patient health. In addition, we propose, as a novel and more effective strategy for combating PD, the adoption of a combined approach through the activation of both endogenous or exogenous antioxidants and the clearance system that eliminates ROS-produced damaged mitochondria (known as mitophagy).
Antioxidants: preclinical and clinical studies in PD
As nigrostriatal dopaminergic neuron degeneration in PD patients is related to oxidative stress and mitochondrial dysfunctions, general antioxidants or mitochondrial-targeted antioxidants (MTAs) may be a fruitful therapy for counteracting PD. Unlike the ubiquitous antioxidant, a common feature of MTAs is the presence of a positive charge in the lipophilic cation, known as triphenylphosphonium (TPP), necessary for crossing the mitochondrial membranes (Murphy and Smith, 2007). In this section, we list the most promising MTAs and general antioxidants in treating PD, their structures being reported in Figures 2 and 3.
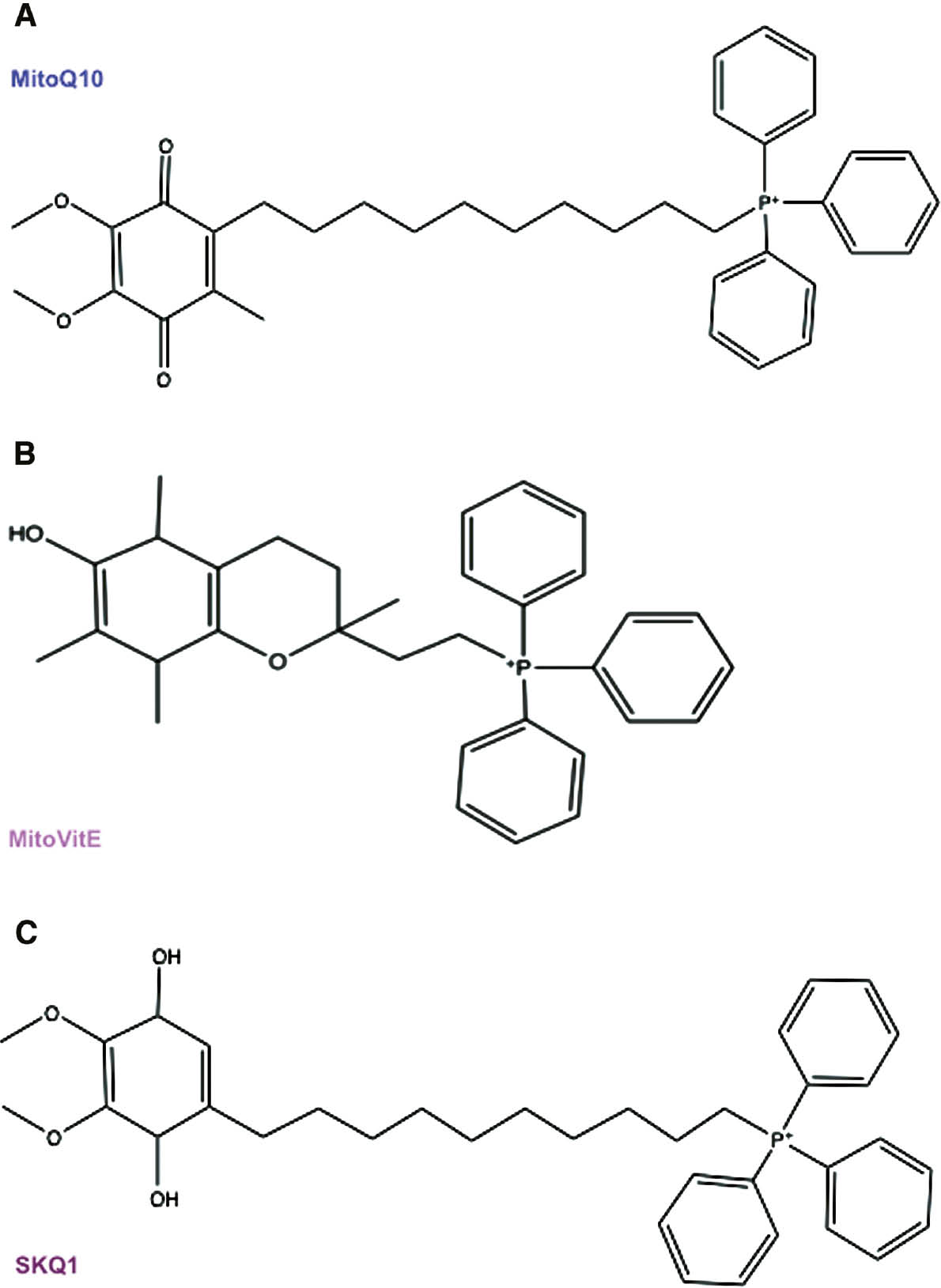
MTAs.
Structures of TPP+-containing molecules that seem to be able to fight PD, such as MitoQ10 (A), MitoVitE (B), and SkQ1 (C).
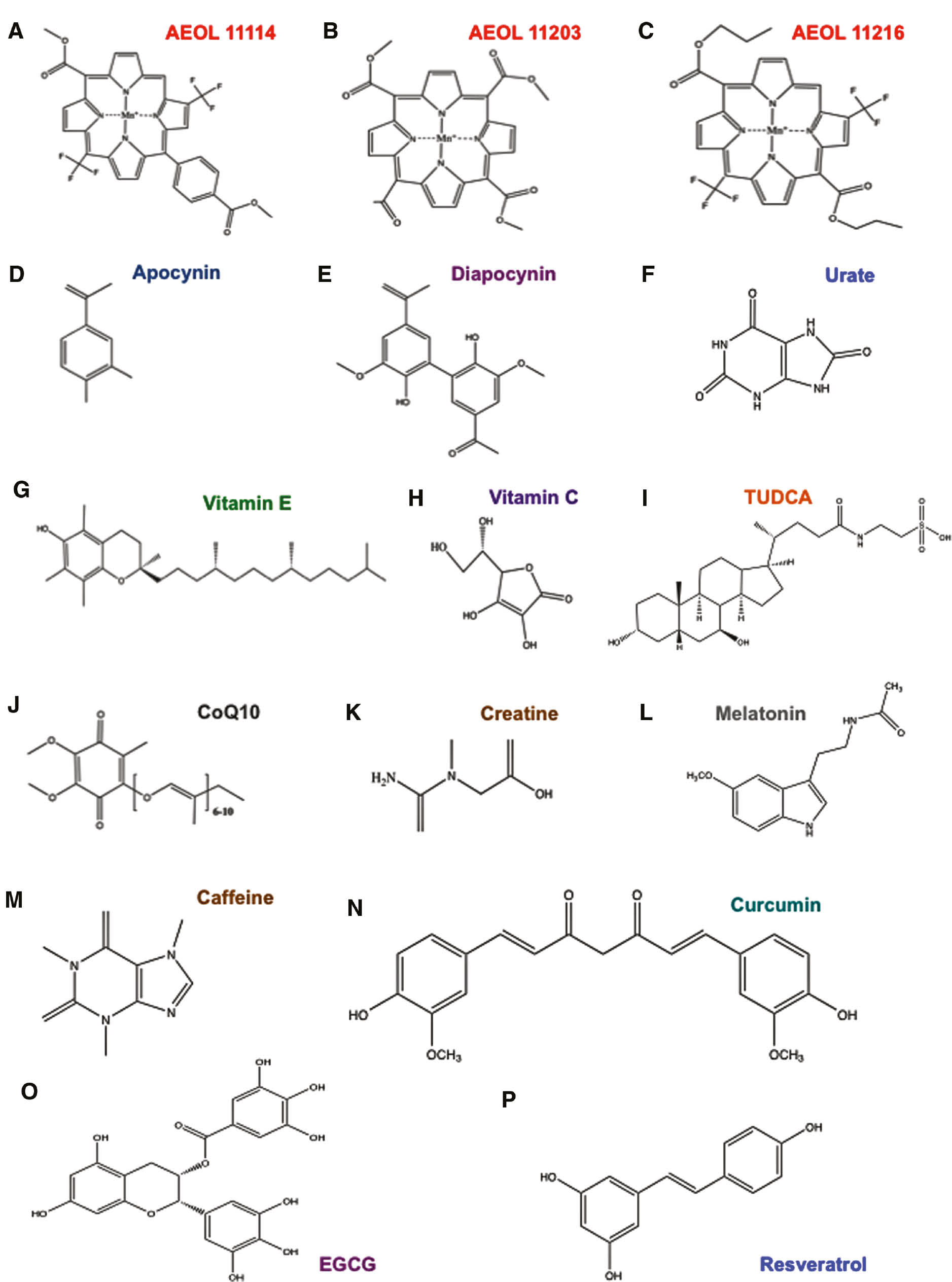
Main general antioxidants.
Structures of a series of antioxidants that counteract Parkinsonism, such as AEOL11114 (A), AEOL11203 (B), AEOL11216 (C), apocynin (D), diapocynin (E), urate (F), vitamin E (G), vitamin C (H), TUDCA (I), CoQ10 (J), creatine (K) and melatonin (L), caffeine (M), curcumin (N), EGCG (O), resveratrol (N).
MTAs
MitoQ
Mitoquinone (MitoQ) constitutes a 10-carbon aliphatic carbon chain necessary for TPP covalent binding to the endogenous antioxidant CoQ10 (Figure 2A). MitoQ is able to scavenge peroxyl, peroxynitrite, and superoxide and, after completing its function, is reduced to the active form ubiquinol by the complex II of the respiratory chain (Smith and Murphy, 2011). As in vitro and in vivo studies demonstrated the beneficial effects of MitoQ, it underwent phase I and then phase II clinical trials (Gane et al., 2010; Snow et al., 2010). Despite its promising role, no neuroprotective benefits were detected in PD patients after MitoQ oral intake (Snow et al., 2010). Further studies are, thus, necessary to better analyze the therapeutic effect of MitoQ in PD patients.
MitoVitE
MitoVitE is the first MTA to be discovered and consists of TPP linked to the α-tocopherol of vitamin E through a two-carbon chain (Figure 2B). Despite its fast intake in the mitochondria of cell culture or mice tissue subjected to oxidative damage, the use of MitoVitE in patients is still being debated. In fact, although MitoVitE has a protective effect in both cellular and animal models (Jauslin et al., 2003; Zhang et al., 2012), no beneficial effects were observed in vivo in striatal neurons. Moreover, high doses of MitoVitE are neurotoxic (Convey et al., 2006).
SkQ1
SkQ1 constitutes the TPP attached to plastoquinone (Figure 2C). This structure allows the incorporation of SkQ1 into the inner mitochondrial membrane (IMM) where it protects from ROS-induced oxidation (Skulachev et al., 2011). It was observed that SkQ1 treatment reduces degeneration of dopaminergic neurons of the substantia nigra and ventral tegmental area (VTA) in PD mouse models (Pavshintsev et al., 2017).
General antioxidants
Metalloporphyrins
Synthetic catalytic antioxidants known as AEOL 11114, AEOL 11203, and AEOL 11216 metalloporphyrins show promising effects for PD treatment due to their ability to penetrate the blood brain barrier together with their oral bioavailability and longer plasma elimination half-lives (Liang et al., 2017) (Figure 3A–C).
Apocynin and diapocynin
Apocynin (4-hydroxy-3-methoxyacetophenone) and its derivate diapocynin are plant antioxidants that function as NADPH oxidase inhibitors. Their protective role in PD model systems was evaluated in vivo (Vejrazka et al., 2005; Luchtefeld et al., 2008) (Figure 3D, E).
Urate
Acid uric or urate is a scavenger of superoxide, hydroxyl radical, single oxygen, and also an iron chelator (Cohen et al., 1984) (Figure 3F). Although over the year it was shown to improve the condition of PD patients, the treatment is limited as urate in high levels causes gout (Choi et al., 2004).
Vitamins
The antioxidant function of vitamins in counteracting oxidative stress was discovered in 1992 (Sies et al., 1992). In particular, vitamin E efficacy to prevent PD progression was demonstrated by several groups (Zhang et al., 2002). By contrast, the role of vitamin C is still controversial (Zhang et al., 2002; Etminan et al., 2005) (Figure 3G, H).
TUDCA
Tauroursodeoxycholic acid (TUDCA) is a derivate from tauroine conjugated to the endogenous bile acid ursodeoxycholic acid (UDCA) (Figure 3I). It was observed that TUDCA improves mitochondrial functions in PD patient-derived fibroblasts (Mortiboys et al., 2015) and has a neuroprotective role in the nematode-PD model (Ved et al., 2005). Moreover TUDCA acts as an antioxidant in several neurodegenerative diseases, including ALS, AD, and HD (Keene et al., 2002; Nunes et al., 2012; Elia et al., 2016). Recently, Rosa and co-workers demonstrated that TUDCA has a neuroprotective effect by stimulating mitophagy in in vitro and in vivo models of PD (see the “Pharmacological inducers of mitophagy” section below) (Zheng et al., 2017).
CoQ10
In vitro and in vivo studies demonstrated the beneficial role of coenzyme Q10 (CoQ10) in PD (Beal et al., 1998; Horvath et al., 2003; Moon et al., 2005). At high doses (1200 mg daily), CoQ10 shows beneficial results in PD rating (Shults et al., 2002) and is currently in the clinical trial phase III (Figure 3J).
Creatine
Creatine is a nitrogenous derivative of the guanidinium cation necessary for supplying energy in vertebrate muscle and nerve cells (Figure 3K). In addition, its antioxidant function was well established in PD mice models (Matthews et al., 1999). Clinical studies demonstrate that creatine has a neuroprotective role in PD patients. At present, creatine is still in the phase III clinical trial, in a long-term study on 1720 patients with PD (Bloom, 2007; Couzin, 2007).
Melatonin
Melatonin is a pineal hormone with an antioxidant function that was demonstrated in different animal models of PD (Saravanan et al., 2007; Borah and Mohanakumar, 2009). Melatonin was shown to block the pro-oxidant effect of both dopamine and l-dopa in an in vitro study (Miller et al., 1996). Moreover, it preserves the activity of tyrosine hydroxylase (TH) in MPP+-treated rats (Jin et al., 1998). Additionally, melatonin is used to reduce REM sleep behavior disorder (RBD), which is a typical feature of parkinsonism and other forms of dementia (Aurora et al., 2010) (Figure 3L).
Caffeine
As caffeine is in coffee and other beverages, it is the most widely consumed psychoactive drug. Different groups found that caffeine has a neuroprotective effect, attenuating the reduction in striatal dopamine, 3,4-dihydroxyphenylacetic acid (DOPAC) and DAT-binding sites (Chen et al., 2001; Joghataie et al., 2004). More recently, Negida and colleagues revealed that caffeine reduces oxidative stress in PD through its antioxidant function, at the mitochondria. In addition, they found that caffeine binds α-synuclein proteins preventing their aggregation in Lewy’s bodies and enhances the number of dopamine receptors (Negida et al., 2017) (Figure 3M).
Curcumin
Curcumin is a plant diarylheptanoid with a bright yellow color. It is widely used in cooking for its anti-inflammatory and antioxidant properties. Curcumin was shown to alleviate motor deficits and neuronal damage in 6-OHDA (6-hydroxydopamine)-treated rats. In fact, it inhibits the conversion of MPTP to MPP+ toxic metabolite (Rajeswari and Sabesan, 2008) (Figure 3N).
Epigallocatechin gallate
Epigallocatechin gallate (EGCG) is a plant catechin typical of the green tea. EGCG has a neuroprotective effect in the MPTP (1-metil 4-fenil 1,2,3,6-tetraidro-piridina) in vivo model of PD. In particular, EGCG controls ferroportin, which is the iron-export protein in the substantia nigra, and alleviates oxidative stress, thus, conferring a rescue effect against MPTP toxicity (Xu et al., 2017) (Figure 3O).
Resveratrol
Resveratrol, which is a polyphenol of red grapes, was shown to reduce mitochondria dysfunction of 6-OHDA dopaminergic neurons of rat (Jin et al., 2008). Moreover, a diet rich in resveratrol combats MPTP neurotoxicity (Blanchet et al., 2008). This polyphenol stimulates SIRTUIN-1 activity, thus, controlling various transcription factors and co-activators, including PGC1-α that regulates mitochondria biogenesis (Peng et al., 2016) (Figure 3P).
Iron chelators
As iron accumulation was found in the nervous system of patients, different iron chelators were used in preclinical studies of PD (Weinreb et al., 2013). Among them, desferrioxamine (DFO) preserves striatal dopamine degeneration induced by 6-OHDA treatment, thus, partially restoring the normal behavior (Ben-Shachar et al., 1992). Despite its functional role, DFO is a large molecule, and its size limits the blood brain barrier crossing (Zheng et al., 2005). Novel iron chelators were tested in the in vitro model of PD (Shachar et al., 2004).
Besides these general antioxidants, the combination of R-α-lipoic acid and acetyl-l-carnitine has also beneficial effects in the in vitro model of PD, stimulating mitochondria biogenesis and reducing oxidative stress (Zhang et al., 2010).
Mitophagy
Antioxidant administration in combination with new approaches focused on the reduction of ROS could be more effective in halting neurodegeneration in PD.
Mitochondria are the main source of ROS production during oxidative phosphorylation for ATP production (Lenaz, 2001). Functional mitochondria maintenance is fundamental for cellular behavior, in particular, for neuronal cells. Despite its low mass, the brain consumes 20% of the body’s oxygen for the production of ATP (Attwell and Laughlin, 2011). The majority of ATP production comes from the mitochondrial respiration chain, although glycolysis contributes to ATP generation (Rangaraju et al., 2014). Correct mitochondria functioning is, thus, crucial in order to regulate, in each region of the neuron, the balance between energy supply and energy demand. Therefore, some mitochondria remain in the soma, while others are carried along axons and dendrites in order to form energy sites in the presynaptic terminals and in proximity of the nodes of Ranvier (Berthold et al., 1993; Shepherd and Harris, 1998).
A selective elimination of damaged mitochondria through mitophagy and the maintenance of healthy mitochondria are, thus, both mandatory for cellular viability (Palikaras and Tavernarakis, 2014). Mitophagy is the most characterized selective autophagy pathway, and it is regulated by the outer mitochondrial membrane (OMM), IMM, or cytosolic proteins named mitophagy receptors (Hamacher-Brady and Brady, 2016).
Mitophagy receptors
Upon mitophagy induction, mitophagy receptors are recruited to damaged mitochondria. The engulfment of undesired mitochondria into the autophagy machinery needs a direct interaction between mitophagy receptors (see Table 1) and the autophagosome markers microtubule-associated proteins 1 light chain 3 (LC3s) or GABA type A receptor-associated proteins (GABARAPs). The binding is mediated by a particular sequence on mitophagy receptors, known as the LC3-interacting region (LIR) motif. Moreover, two crucial posttranslational modifications occur during mitophagy: phosphorylation and ubiquitylation. The former can be a positive or negative change that allows or inhibits the mitophagy receptor-autophagosome marker binding (Hamacher-Brady and Brady, 2016). The latter, takes place on the OMM proteins and is a signal for cytosolic mitophagy receptor recruitment and, therefore, for mitochondria elimination (Yamano et al., 2016). This pathway is able to selectively eliminate damaged mitochondria in order to maintain cellular homeostasis and to preserve mitochondria vitality.
Overview of the known mitophagy pathways.
| PINK1/PARK2-dependent pathway |
| PINK1 is a mitochondria kinase that functions as a sensor of mitochondria status. Upon mitochondria stress, PINK1 translocates to the OMM, thus, mediating a phospho-ubiquitylation signal on s65 of both ubiquitin and PARK2. In particular, the cytosolic E3 ligase PARK2 is recruited to the mitochondria in order to amplify the ubiquitilation, thus, leading to the recruitment of cytosolic mitophagy receptors OPTN, NDP52, P62, NBR1, and TAX1BP1 (Lazarou et al., 2015). Lazarou and colleagues observed that OPTN and NDP52 are the primary receptors, and the others are dispensable for mitophagy. In addition, the IMM PHB2 regulates mitophagy during paternal mtDNA degradation cooperating with PARK2 (Wei et al., 2017). |
| FUNDC1: is an OMM protein whose role in mitophagy was clarified since 2012 (Wu et al., 2014). It possesses an LIR motif that is regulated posttranslationally by phosphorylation. Wu and colleagues demonstrated that FUNDC1 mediates mitophagy upon drug induction and also following physiological stimuli (e.g., hypoxia), and its action depends on phosphorylation of Ser/Tyr sites near the LIR motif. |
| PINK1/PARK2-independent pathway |
| BNIP3L: this OMM protein is the mammalian homolog of NIX, and it regulates mitophagy upon reticulocyte differentiation and after hypoxic stress (Novak et al., 2010). In analogy with FUNDC1, also the BNIP3L activity in mitophagy is regulated by phosphorylation. |
| BCL2-L13: the mammalian homolog of Atg32 interacts with LC3B and regulates mitochondria-selective removal (Murakawa et al., 2015). |
| FKBP8/FKBP38: is a member of the FK 506-binding protein family, and it interacts preferentially with LC3A in order to induce mitophagy. Interestingly, although FKBP8 is an OMM adaptor protein and promotes mitophagy, it is able to escape from the acidified mitochondria preserving itself (Bhujalab et al., 2017). |
| PINK1/PARK2-dependent/independent pathway |
| AMBRA1: is a cytosolic protein localized also on the OMM (Strappazzon et al., 2011) that interacts with the autophagosome marker LC3B during the mitophagic process (Strappazzon et al., 2015). It is able to mediate mitophagy interacting with PARK2 and amplifying the PINK1-dependent digestion pathway (Van Humbeeck et al., 2011; Strappazzon 2015) or promoting mitophagy by itself, independently of PINK1/PARK2 pathway, once expressed at the mitochondria (Strappazzon et al., 2015; Di Rita et al., 2018). |
Cardiolipin
Cardiolipin or 1,3-bis(sn-3′-phosphatidyl)-sn-glycerol is a lipid of the IMM that functions as a sensor of mitochondrial stress. In detail, upon mitophagy induction, cardiolipin is externalized on the mitochondria where it interacts with the autophagosome marker LC3-II in order to mediate the engulfment into the autophagosome machinery (Chu et al., 2013).
Mitophagy receptors as therapeutic targets for PD
Neurodegenerative diseases, including PD, AD, and HD, are characterized by the accumulation of damaged mitochondria that produces critical effects on neuron functionality (Johri and Beal, 2012). Recent evidence shows that the selective mitochondria autophagy contributes to improving mitochondrial status and cellular homeostasis of neurons. In particular, two mitophagy receptors, Nix and AMBRA1, show a central role in delaying cell death in PD models through an antioxidant action. In particular, Koentjoro and colleagues demonstrated that when the main mitophagy process (PINK1/PARK2-mediated mitophagy) is abrogated in PD dopaminergic neurons, Nix is able to stimulate a damaged mitochondria selective removal that preserves dopaminergic neurons. Moreover, phorbol 12-myristate 13-acetate (PMA) was reported to induce Nix expression, leading to a reduction in ROS production in PD dopaminergic neurons (Koentjoro et al., 2017).
In a similar manner, AMBRA1-mediated mitophagy is able to rescue mitophagy in fibroblasts of PD patients in which PINK1 or PARK2 are mutated (Strappazzon et al., 2015). In addition, a particular form of AMBRA1, localized at the OMM (AMBRA1ActA) is able to stimulate mitophagy in an in vitro model of PD and reduces cell death induced by rotenone or 6-OHDA treatments, by limiting selectively the source of oxidative stress (Di Rita et al., 2018). In line with such evidence, both Nix and AMBRA1 could be considered as attractive therapeutic targets in drug development and human application.
Moreover, Chu and colleagues demonstrated that cardiolipin drives mitophagy in response to low doses of rotenone or 6-OHDA through a direct interaction with LC3. Despite the fact that, at present, cardiolipin has no antioxidant functions, in line with AMBRA1 and NIX, it possesses phosphorylation sites in the offset of its LIR motif that could control mitophagy in a fine-tuned system. These analogies could suggest that these three mitophagy regulators could control mitochondria degradation through the autophagy machinery in PD and that their synergic action could be useful in reducing PD progression by limiting the accumulation of damaged mitochondria in neurons.
To date, scientists are able to induce mitophagy in vitro and in vivo, mainly using pharmacological compounds, i.e. antimycin A, or oligomycin or carbonyl cyanide-4-(trifluoromethoxy) phenylhydrazone (FCCP), and simulating hypoxic/ischemic conditions. However, in the near future, the generation of a drug that could specifically stimulate both AMBRA1- and NIX-mediated mitophagy presents an attractive challenge for counteracting PD by strongly reducing oxidative stress and preserving mitochondria health in dopaminergic neurons.
Inducers of mitophagy
As maintaining mitophagy is crucial in age-related diseases, several inducers of mitophagy were developed and can be classified in pharmacological or natural compounds.
Pharmacological inducers of mitophagy
NRF2 induction through PMI
NRF2 (nuclear factor erythroid 2-related factor 2) is a transcription factor that controls the expression of several genes, such as PINK1, nuclear dot protein 52 (NDP52) and sequestosome 1 (P62) by linking the antioxidant response element (ARE) in their promoter regions. It is, thus, considered the master regulator of the cellular redox homeostasis. NRF2 activity is able to defend cells against mitochondrial toxins; indeed, its function is suppressed in mitochondrial disorders such as PD (Georgakopoulos et al., 2017). PMI/HB229 is an inducer of NRF2 and stimulates P62 production by reversibly inhibiting the activity of Kelch-like ECH-associated protein 1 (KEAP1), which is important for NRF2 degradation by the proteasome.
Mechanistically, PMI interferes protein-protein interaction (PPI) between NRF2 and KEAP1, blocking the NRF2 ubiquitylation and favoring its nuclear accumulation. Consequently, NRF2 activates gene production; among the genes activated, P62 becomes upregulated, and mitophagy is activated. Of note, this effect of NRF2 is independent of the PINK1/PARK2 pathway.
Interestingly, the mitophagic activity of PMI is related to the redox status of mitochondria as selective scavenging of mitochondrial superoxides (i.e. NAC, Mito-TEMPOL) inhibits the PMI-induced mitophagy. In this context, the inhibition of mitophagy by Mito-TEMPOL is intriguing, suggesting that ROS is necessary for inducing PMI-dependent mitophagy.
As PMI has an antioxidant action and induces mitophagy independent of PINK1/PARK2, its action has considerable potential in the case of PD patients, and as such, merits further study.
MitoQ
Mitoquinone was originally used to suppress ROS production and lipid peroxidation as explained above in the “Antioxidants: preclinical and clinical studies in PD” section. Interestingly, MitoQ was shown to reduce oxidative stress through mitophagy induction in an in vivo model of diabetic kidney disease (DKD). In fact, Xiao and collaborators demonstrated that MitoQ activates mitochondria-selective removal controlling the NRF2 expression, activity, and translocation, which, in turn, cooperates with PINK1 mitophagy induction (Xiao et al., 2017). Moreover, MitoQ was shown to support mitophagy by increasing mitochondrial membrane depolarization (ΔΨm) in aggressive cancer cells (Biel and Rao, 2018).
Phorbol 12-myristate 13-acetate (PMA)
Phorbol 12-myristate 13-acetate (PMA) was shown to favor Nix transcriptional activation in vitro through the stimulation of protein kinase C α (PRKCA) activity and by promoting the binding of the transcription factor SP1 to GC-rich elements in the Nix promoter in cultured neonatal rat cardiac myocytes (Galvez et al., 2006). More recently, PMA was found to stimulate mitophagy in PD models by increasing Nix expression (Koentjoro et al., 2017). This pharmacological induction of Nix promotes CCCP-induced mitophagy in the PARK2-and PINK1-related PD patient cell lines. Drugs targeting Nix are, thus, expected to provide neuroprotective action in PD patients.
TUDCA
TUDCA strongly prevents CCCP-induced cell death, ROS generation, and mitochondrial damage in neuroblastoma cell line. The neuroprotective role of TUDCA in this cell model is mediated by PARK2-dependent mitophagy. The demonstration that pharmacological upregulation of mitophagy by TUDCA prevents neurodegeneration provides new insights into the use of TUDCA in order to favor mitochondrial quality control in PD context.
Natural compound inducers of mitophagy
Melanoidin
Melanoidin derives from aged vinegar or pomegranate extract (PE) and is a well-known antioxidant (Delgado-Andrade et al., 2005). Yang and colleagues found that melanoidins from Shanxi aged vinegar-induced mitophagy (Yang et al., 2014). In particular, they reported that melanoidins reduce ROS in normal human liver cells and mouse livers through a mitophagy-dependent pathway. These results support the fact that melanoidins are able to reduce ROS levels through mitophagy activation, by degrading damaged mitochondria, the source of oxidative stress. In addition, to be direct free radical scavengers, melanoidins are, thus, mitophagy inducers in the context of injury-induced hepatocytes and cardiomyocites (Yang et al., 2016).
Melanoidins appears, thus, as new therapeutic target to be tested in the context of PD.
Of note, during the roasting process of coffee, melanoidins are formed (Moreira et al., 2012). As coffee could be one of the main sources of melanoidins in the human diet, their health implications are of great interest. Interestingly, epidemiologic links exist between caffeine and lower PD risk (Ross et al., 2000; Hernán et al., 2002). However, the potential of coffee diet on health of PD patients were tested, but caffeine did not show improvement of PD symptomatic effects (Postuma et al., 2017). It should be interesting to test the effect of melanoidins directly on PD patients instead of caffeine.
Urolithin A
Urolithin A is a microflora human metabolite originating from the digestion by intestinal bacteria. Its precursor is the ellagic acid (ellagitannin family). When this acid is consumed by diet, the bacterial microflora is able to transform it into urolithin A. Some foods are rich in ellagic acid (e.g., blackberries, cranberries, grapes, pecans raspberries, etc.); however, among these foods, it appears that pomegranates have the highest potential for producing the urolithin A.
This metabolite was shown to prevent accumulation of damaged mitochondria and improve lifespan in nematode and exercise capacity in mice (Ryu et al., 2016).
Again, it is expected that urolithin A may favor health of PD patients by causing mitophagy and reducing oxidative stress.
Spermidine
Spermidine is a small organic molecule of the polyamine family and controls several physiological processes such as cell growth, proliferation, and survival (Gevrekci, 2017). Interestingly, sperimidine levels decline with age in some tissues such as the heart and kidneys (Nishimura et al., 2006). A diet enriched with spermidine was found to improve lifespans of yeast, nematodes flies, and mice (Eisenberg et al., 2009). Moreover, spermidine activates mitophagy in aged cardiomyocytes, thus, repairing mitochondrial activity (Eisenberg et al., 2016). Qi and colleagues demonstrated that spermidine-induced mitophagy is dependent on ATM (ataxia-telangiectasia-mutated kinase) (Qi et al., 2016). Administration of spermidine delays PD-related degenerative processes in the fruit fly Drosophila melanogaster and in the nematode Caenorhabditis elegans, two established model systems for PD pathology (Büttner et al., 2014). Spermidine-dietary supplementation, thus, has the potential for improving the health of PD patients.
Resveratrol and other GSE components
Different studies demonstrated that resveratrol alleviates symptoms and is beneficial against cancers, heart disorders, and Alzheimer’s disease. Recently, Wu and colleagues reported that daily assumption of GSE (grape skin extracts) alleviates mitochondrial dysfunctions in a D. melanogaster model of PD. In particular, besides resveratrol, GSE contains quercetin, myricetin, catechins, tannins, anthocyanidins, and ferulic acid. This work reported that there is a link between mitophagy stimulation and resveratrol beneficial effects necessary for improving lifespan (Wu et al., 2018).
Conclusion/perspectives
PMA, MitoQ, TUDCA, PMI, or GSE administration in mice models of PD, and eventually in PD patients, could, thus, offer an effect method of combating PD stimulating both antioxidant responses and mitophagy.
Another exciting issue is the proposal to increase the mitophagy process in PD patients through melanoidin, urolithin A, or spermidine consumption. Thus, there is a need to generate and test specialized diet to fit the PD context (see the proposed model in Figure 4).
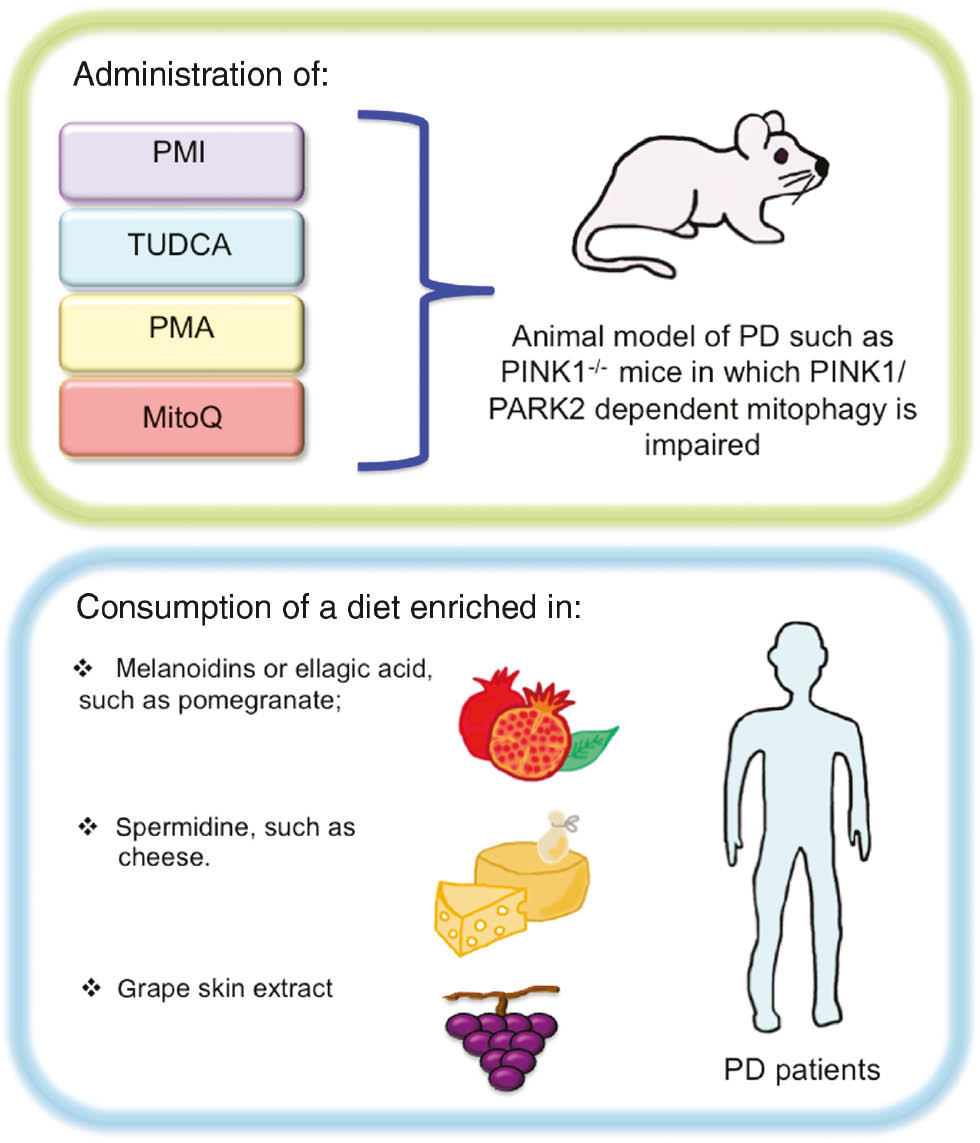
Antioxidants and/or mitophagy inducer administration in order to counteract PD.
The use of pharmacological compounds able to activate both mitophagy and antioxidant responses could be a promising strategy in PD mice models in order to delay neurodegeneration. In addition, it could be worth testing a diet enriched in natural mitophagy inducers and/or antioxidants, such as pomegranate or cheese, as a method of limiting oxidative stress and, thus, neurodegeneration in PD patients.
About the authors

Anthea Di Rita graduated with a degree in Cellular and Molecular Biology in 2014 at the University of Rome Tor Vergata (Italy). During her PhD, she acquired a solid knowledge in the fields of autophagy, mitophagy, and cell death. She is currently a post-doctoral fellow working under the supervision of Dr. Flavie Strappazzon at the IRCCS Fondazione Santa Lucia in Rome (Italy).

Flavie Strappazzon is a junior head of a laboratory at the IRCCS Fondazione Santa Lucia in Rome (Italy) and an external teacher at the University of Rome "Tor Vergata". She graduated with a degree in Cellular Biology in 2007 at the University of Grenoble (France). During this period, she acquired a solid knowledge in Neurosciences. Next, she moved to Italy to perform two post-doctoral trainings in the field of Autophagy. In 2018, she started her research group working on selective autophagy and, in particular, on mitophagy in the context of multiple sclerosis and Huntington’s disease (IRCCS Santa Lucia, Rome).
Acknowledgments
We wish to thank Roche Company who supports F.S. (Grant “Roche per la ricerca 2017”). We thank Dr. M. Bennett for proofreading the work.
References
Attwell, D. and Laughlin, S.B. (2011). An energy budget for signaling in the grey matter of the brain. J. Cereb. Blood Flow Metab. 21, 1133–1145.10.1097/00004647-200110000-00001Suche in Google Scholar
Aurora, R.N., Zak, R.S., Maganti, R.K., Auerbach, S.H., Casey, K.R., Chowdhuri, S., Karippot, A., Ramar, K., Kristo, D.A., and Morgenthaler, T.I. (2010). Best practice guide for the treatment of REM sleep behavior disorder (RBD). J. Clin. Sleep Med. 6,85–95.10.5664/jcsm.27717Suche in Google Scholar
Beal, M.F., Matthews, R.T., Tieleman, A., and Shults, C.W. (1998). Coenzyme Q10 attenuates the 1-methyl-4-phenyl-1,2,3,tetrahydropyridine (MPTP) induced loss of striatal dopamine and dopaminergic axons in aged mice. Brain Res. 783, 109–114.10.1016/S0006-8993(97)01192-XSuche in Google Scholar
Bender, A., Krishnan, K.J., Morris, C.M., Taylor, G.A., Reeve, A.K., Perry, R.H., Jaros, E., Hersheson, J.S., Betts, J., Klopstock, T., et al. (2006). High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 38, 515–517.10.1038/ng1769Suche in Google Scholar PubMed
Ben-Shachar, D., Eshel, G., Riederer, P., and Youdim, M.B. (1992). Role of iron and iron chelation in dopaminergic-induced neurodegeneration: implication for Parkinson’s disease. Ann. Neurol. 1992, 32.10.1002/ana.410320718Suche in Google Scholar PubMed
Berthold, C.H., Fabricius, C., Rydmark, M., and Andersén, B. (1993). Axoplasmic organelles at nodes of Ranvier. I. Occurrence and distribution in large myelinated spinal root axons of the adult cat. J. Neurocytol. 22, 925–940.10.1007/BF01218351Suche in Google Scholar PubMed
Biel, T.G. and Rao, V.A. (2018). Mitochondrial dysfunction activates lysosomal-dependent mitophagy selectively in cancer cells. Oncotarget 9, 995–1011.10.18632/oncotarget.23171Suche in Google Scholar PubMed PubMed Central
Blanchet, J., Longpré, F., Bureau, G., Morissette, M., DiPaolo, T., Bronchti, G., and Martinoli, M.G. (2008). Resveratrol, a red wine polyphenol, protects dopaminergic neurons in MPTP-treated mice. Prog. Neuropsychopharmacol. Biol. Psychiatry 32, 1243–1250.10.1016/j.pnpbp.2008.03.024Suche in Google Scholar PubMed
Bloom, M.Z. (2007). NIH announces phase III clinical trial of creatine for Parkinson’s disease. Consult. Pharm. 22, 378.Suche in Google Scholar
Bose, A. and Beal, M.F. (2016). Mitochondrial dysfunction in Parkinson’s disease. J. Neurochem. 139, 216–231.10.1111/jnc.13731Suche in Google Scholar PubMed
Bhujabal, Z., Birgisdottir, Å.B., Sjøttem, E., Brenne, H.B., Øvervatn, A., Habisov, S., Kirkin, V., Lamark, T., and Johansen, T. (2017). FKBP8 recruits LC3A to mediate Parkin-independent mitophagy. EMBO Rep. 18, 947–961.10.15252/embr.201643147Suche in Google Scholar PubMed PubMed Central
Borah, A. and Mohanakumar, K.P. (2009). Melatonin inhibits 6-hydroxydopamine production in the brain to protect against experimental Parkinsonism in rodents. J. Pineal Res. 47, 293–300.10.1111/j.1600-079X.2009.00713.xSuche in Google Scholar
Büttner, S., Broeskamp, F., Sommer, C., Markaki, M., Habernig, L., Alavian-Ghavanini, A., Carmona-Gutierrez, D., Eisenberg, T., Michael, E., Kroemer, G., et al. (2014). Spermidine protects against α-synuclein neurotoxicity. Cell Cycle 13, 3903–3908.10.4161/15384101.2014.973309Suche in Google Scholar
Campbell, G.R., Ziabreva, I., Reeve, A.K., Krishnan, K.J., Reynolds, R., Howell, O., Lassmann, H., Turnbull, D.M., and Mahad, D.J. (2011). Mitochondrial DNA deletions and neurodegeneration in multiple sclerosis. Ann. Neurol. 69, 481–492.10.1002/ana.22109Suche in Google Scholar
Chen, J.F., Xu, K., Petzer, J.P., Staal, R., Xu, Y.J., Beilstein, M., Sonsalla, P.K., Castagnoli, K., Castagnoli, N., and Schwarzschild, M.A. (2001). Neuroprotection by caffeine and A(2A) adenosine receptor inactivation in a model of Parkinson’s disease. J. Neurosci. 21, RC143.10.1523/JNEUROSCI.21-10-j0001.2001Suche in Google Scholar
Choi, H.K., Atkinson, K., Karlson, E.W., Willett, W., and Curhan, G. (2004). Alcohol intake and risk of incident gout in men: a prospective study. Lancet 363, 1277–1281.10.1016/S0140-6736(04)16000-5Suche in Google Scholar
Cohen, A.M., Aberdroth, R.E., and Hochstein, P. (1984). Inhibition of free radical-induced DNA damage by uric acid. FEBS Lett. 174, 147–150.10.1016/0014-5793(84)81094-7Suche in Google Scholar
Couzin, J. (2007). Clinical research. Testing a novel strategy against Parkinson’s disease. Science 315, 1778.10.1126/science.315.5820.1778Suche in Google Scholar PubMed
Covey, M.V., Murphy, M.P., Hobbs, C.E., Smith, R.A., and Oorschot, D.E. (2006). Effect of the mitochondrial antioxidant, Mito Vitamin E, on hypoxic-ischemic striatal injury in neonatal rats: a dose-response and stereological study. Exp. Neurol. 199, 513–519.10.1016/j.expneurol.2005.12.026Suche in Google Scholar PubMed
Chu, C.T., Ji, J., Dagda, R.K., Jiang, J.F., Tyurina, Y.Y., Kapralov, A.A., Tyurin, V.A., Yanamala, N., Shrivastava, I.H., Mohammadyani, D., et al. (2013). Cardiolipin externalization to the outer mitochondrial membrane acts as an elimination signal for mitophagy in neuronal cells. Nat. Cell Biol. 15, 1197–1205.10.1038/ncb2837Suche in Google Scholar PubMed PubMed Central
Delgado-Andrade, C., Rufián-Henares, J.A., and Morales, F.J. (2005). Assessing the antioxidant activity of melanoidins from coffee brews by different antioxidant methods. J. Agric. Food Chem. 53, 7832–7836.10.1021/jf0512353Suche in Google Scholar PubMed
Devi, L., Raghavendran, V., Prabhu, B.M., Avadhani, N.G., and Anandatheerthavarada, H.K. (2008). Mitochondrial import and accumulation of alpha-synuclein impair complex I in human dopaminergic neuronal cultures and Parkinson disease brain. J. Biol. Chem. 283, 9089–9100.10.1074/jbc.M710012200Suche in Google Scholar
Di Rita, A., D’Acunzo, P., Simula, L., Campello, S., Strappazzon, F., and Cecconi, F. (2018). AMBRA1-mediated mitophagy counteracts oxidative stress and apoptosis onduced by neurotoxicity in human neuroblastoma SH-SY5Y cells. Front. Cell Neurosci. 12, 92.10.3389/fncel.2018.00092Suche in Google Scholar
Dutta, R., McDonough, J., Yin, X., Peterson, J., Chang, A., Torres, T., Gudz, T., Macklin, W.B., Lewis, D.A., Fox, R.J., et al. (2006). Mitochondrial dysfunction as a cause of axonal degeneration in multiple sclerosis patients. Ann. Neurol. 59, 478–489.10.1002/ana.20736Suche in Google Scholar
Eisenberg, T., Knauer, H., Schauer, A., Büttner, S., Ruckenstuhl, C., Carmona-Gutierrez, D., Ring, J., Schroeder, S., Magnes, C., Antonacci, L., et al. (2009). Induction of autophagy by spermidine promotes longevity. Nat. Cell Biol. 11, 1305–1314.10.1038/ncb1975Suche in Google Scholar
Eisenberg, T., Abdellatif, M., Schroeder, S., Primessnig, U., Stekovic, S., Pendl, T., Harger, A., Schipke, J., Zimmermann, A., Schmidt, A., et al. (2016). Cardioprotection and lifespan extension by the natural polyamine spermidine. Nat. Med. 22, 1428–1438.10.1038/nm.4222Suche in Google Scholar
Ekstrand, M.I., Terzioglu, M., Galter, D., Zhu, S., Hofstetter, C., Lindqvist, E., Thams, S., Bergstrand, A., Hansson, F.S., Trifunovic, A., et al. (2007). Progressive parkinsonism in mice with respiratory-chain-deficient dopamine neurons. Proc. Natl. Acad. Sci. USA 104, 1325–1330.10.1073/pnas.0605208103Suche in Google Scholar
Elia, A.E., Lalli, S., Monsurrò, M.R., Sagnelli, A., Taiello, A.C., Reggiori, B., La Bella, V., Tedeschi, G., and Albanese, A. (2016). Tauroursodeoxycholic acid in the treatment of patients with amyotrophic lateral sclerosis. Eur. J. Neurol. 23, 45–52.10.1111/ene.12664Suche in Google Scholar
Etminan, M., Gill, S.S., and Samii, A. (2005). Intake of vitamin E, vitamin C, and carotenoids and the risk of Parkinson’s disease: a meta-analysis. Lancet Neurol. 4, 362–365.10.1016/S1474-4422(05)70097-1Suche in Google Scholar
Gálvez, A.S., Brunskill, E.W., Marreez, Y., Benner, B.J., Regula, K.M., Kirschenbaum, L.A., and Dorn, G.W. (2006). Distinct pathways regulate proapoptotic Nix and BNip3 in cardiac stress. J. Biol. Chem. 281, 1442–1448.10.1074/jbc.M509056200Suche in Google Scholar PubMed
Gane, E.J., Weilert, F., Orr, D.W., Keogh, G.F., Gibson, M., Lockhart, M.M., Frampton, C.M., Taylor, K.M., Smith, R.A., and Murphy, M.P. (2010). The mitochondria-targeted anti-oxidant mitoquinone decreases liver damage in a phase II study of hepatitis C patients. Liver Int. 30, 1019–1026.10.1111/j.1478-3231.2010.02250.xSuche in Google Scholar PubMed
Georgakopoulos, N.D., Frison, M., Alvarez, M.S., Bertrand, H., Wells, G., and Campanella, M. (2017). Reversible Keap1 inhibitors are preferential pharmacological tools to modulate cellular mitophagy. Sci. Rep. 7, 10303.10.1038/s41598-017-07679-7Suche in Google Scholar
Gevrekci, A.Ö. (2017). The roles of polyamines in microorganisms. World J. Microbiol. Biotechnol. 33, 204.10.1007/s11274-017-2370-ySuche in Google Scholar
Hamacher-Brady, A. and Brady, N.R. (2016). Mitophagy programs: mechanisms and physiological implications of mitochondrial targeting by autophagy. Cell Mol. Life Sci. 73, 775–795.10.1007/s00018-015-2087-8Suche in Google Scholar
Hernán, M.A., Takkouche, B., Caamaño-Isorna, F., and Gestal-Otero, J.J. A meta-analysis of coffee drinking, cigarette smoking, and the risk of Parkinson’s disease. Ann. Neurol. 52, 276–284.10.1002/ana.10277Suche in Google Scholar
Horvath, T.L., Diano, S., Leranth, C., Garcia-Segura, L.M., Cowley, M.A., Shanabrough, M., Elsworth, J.D., Sotonyi, P., Roth, R.H., Dietrich, E.H., et al. (2003). Coenzyme Q induces nigral mitochondrial uncoupling and prevents dopamine cell loss in a primate model of Parkinson’s disease. Endocrinology 144, 2757–2760.10.1210/en.2003-0163Suche in Google Scholar
Isobe, C., Abe, T., and Terayama, Y. (2010). Levels of reduced and oxidized coenzyme Q-10 and 8-hydroxy-2′-deoxyguanosine in the cerebrospinal fluid of patients with living Parkinson’s disease demonstrate that mitochondrial oxidative damage and/or oxidative DNA damage contributes to the neurodegenerative process. Neurosci. Lett. 469, 159–163.10.1016/j.neulet.2009.11.065Suche in Google Scholar
Jauslin, M.L., Meier, T., Smith, R.A., and Murphy, M.P. (2003). Mitochondria-targeted antioxidants protect Friedreich ataxia fibroblasts from endogenous oxidative stress more effectively than untargeted antioxidants. FASEB J. 17, 1972–1974.10.1096/fj.03-0240fjeSuche in Google Scholar
Jin, B.K., Shin, D.Y., Jeong, M.Y., Gwag, M.R., Baik, H.W., Yoon, K.S., Cho, Y.H., Joo, W.S., Kim, Y.S., and Baik, H.H. (1998). Melatonin protects nigral dopaminergic neurons from 1-methyl-4-phenylpyridinium (MPP+) neurotoxicity in rats. Neurosci. Lett. 245, 61–64.10.1016/S0304-3940(98)00170-0Suche in Google Scholar
Jin, F., Wu, Q., Lu, Y.F., Gong, Q.H., and Shi, J.S. (2008). Neuroprotective effect of resveratrol on 6-OHDA-induced Parkinson’s disease in rats. Eur. J. Pharmacol. 600, 78–82.10.1016/j.ejphar.2008.10.005Suche in Google Scholar PubMed
Joghataie, M.T., Roghani, M., Negahdar, F., and Hashemi, L. (2004). Protective effect of caffeine against neurodegeneration in a model of Parkinson’s disease in rat: behavioral and histochemical evidence. Parkinsonism Relat. Disord. 10, 657–661.10.1016/j.parkreldis.2004.06.004Suche in Google Scholar PubMed
Johri, A. and Beal, M.F. (2012). Mitochondrial dysfunction in neurodegenerative diseases. J. Pharmacol. Exp. Ther. 342, 619–630.10.1124/jpet.112.192138Suche in Google Scholar
Keene, C.D., Rodrigues, C.M.P., Eich, T., Chhabra, M.S., Steer, C.J., and Low, W.C. (2002). Tauroursodeoxycholic acid, a bile acid, is neuroprotective in a transgenic animal model of Huntington’s disease. Proc. Natl. Acad. Sci. USA 99, 10671–10676.10.1073/pnas.162362299Suche in Google Scholar
Koentjoro, B., Park, J.S., and Suea, C.M. (2017). Nix restores mitophagy and mitochondrial function to protect against PINK1/Parkin-related Parkinson’s disease. Sci. Rep. 7, 44373.10.1038/srep44373Suche in Google Scholar
Kraytsberg, Y., Kudryavtseva, E., McKee, A.C., Geula, C., Kowall, N.W., and Khrapko, K. (2006). Mitochondrial DNA deletions are abundant and cause functional impairment in aged human substantia nigra neurons. Nat. Genet. 38, 518–520.10.1038/ng1778Suche in Google Scholar
Lazarou, M., Sliter, D.A., Kane, L.A., Sarraf, S.A., Wang, C., Burman, J.L., Sideris, D.P., Fogel, A.I., and Youle, R.J. (2015). The ubiquitin kinase PINK1 recruits autophagy receptors to induce mitophagy. Nature 524, 309–314.10.1038/nature14893Suche in Google Scholar
Lenaz, G. (2001). The mitochondrial production of reactive oxygen species: mechanisms and implications in human pathology. IUBMB Life 52, 159–164.10.1080/15216540152845957Suche in Google Scholar
Liang, L.P., Huang, J., Fulton, R., Pearson-Smith, J.N., Day, B.J., and Patel, M. (2017). Pre-clinical therapeutic development of a series of metalloporphyrins for Parkinson’s disease. Toxicol. Appl. Pharmacol. 326, 34–42.10.1016/j.taap.2017.04.004Suche in Google Scholar
Luchtefeld, R., Luo, R., Stine, K., Alt, M.L., Chernovitz, P.A., and Smith, R.E. (2008). Dose formulation and analysis of diapocynin. J. Agric. Food Chem. 56, 301–306.10.1021/jf072792nSuche in Google Scholar
Matthews, R.T., Ferrante, R.J., Klivenyi, P., Yang, L., Klein, A.M., Mueller, G., Kaddurah-Daouk, R., and Beal, M.F. (1999). Creatine and cyclocreatine attenuate MPTP neurotoxicity. Exp. Neurol. 157, 142–149.10.1006/exnr.1999.7049Suche in Google Scholar
Miller, J.W., Selhub, J., and Joseph, J.A. (1996). Oxidative damage caused by free radicals produced during catecholamine autoxidation: protective effects of O-methylation and melatonin. Free Radic. Biol. Med. 21, 241–249.10.1016/0891-5849(96)00033-0Suche in Google Scholar
Moon, Y., Lee, K.H., Park, J.H., Geum, D., and Kim, K. (2005). Mitochondrial membrane depolarization and the selective death of dopaminergic neurons by rotenone: protective effect of coenzyme Q10. J. Neurochem. 93, 1199–1208.10.1111/j.1471-4159.2005.03112.xSuche in Google Scholar PubMed
Moreira, A.S., Nunes, F.M., Domingues, M.R., and Coimbra, M.A. (2012). Coffee melanoidins: structures, mechanisms of formation and potential health impacts. Food Funct. 3, 903–915.10.1039/c2fo30048fSuche in Google Scholar PubMed
Mortiboys, H., Furmston, R., Bronstad, G., Aasly, J., Elliott, C., and Bandmann, O. (2015). UDCA exerts beneficial effect on mitochondrial dysfunction in LRRK2 (G2019S) carriers and in vivo. Neurology 85, 846–852.10.1212/WNL.0000000000001905Suche in Google Scholar PubMed PubMed Central
Murakawa, T., Yamaguchi, O., Hashimoto, A., Hikoso, S., Takeda, T., Oka, T., Yasui, H., Ueda, H., Akazawa, Y., Nakayama, H., et al. (2015). Bcl-2-like protein 13 is a mammalian Atg32 homologue that mediates mitophagy and mitochondrial fragmentation. Nat. Commun. 6, 7527.10.1038/ncomms8527Suche in Google Scholar PubMed PubMed Central
Murphy, M.P. (2009). How mitochondria produce reactive oxygen species. Biochem. J. 417, 1–13.10.1042/BJ20081386Suche in Google Scholar PubMed PubMed Central
Murphy, M.P. and Smith, R.A. (2007). Targeting antioxidants to mitochondria by conjugation to lipophilic cations. Annu. Rev. Pharmacol. Toxicol. 47, 629–656.10.1002/9780470372531.ch25Suche in Google Scholar
Negida, A., Elfil, M., Attia, Farahat, E., Gabr, M., Essam, A., Attia, D., and Ahmed, H. (2017). Caffeine; the forgotten potential for Parkinson’s disease. CNS Neurol. Disord. Drug Targets 16, 652–657.10.2174/1871527315666161107091149Suche in Google Scholar PubMed
Nishimura, K., Shiina, R., Kashiwagi, K., and Igarashi, K. (2006). Decrease in polyamines with aging and their ingestion from food and drink. J. Biochem. 139, 81–90.10.1093/jb/mvj003Suche in Google Scholar PubMed
Novak, I., Kirkin, V., McEwan, D.G., Zhang, J., Wild, P., Rozenknop, A., Rogov, V., Lohr, F., Popovic, D., Occhipinti, A., et al. (2010). Nix is a selective autophagy receptor for mitochondrial clearance. EMBO Rep. 11, 45–51.10.1038/embor.2009.256Suche in Google Scholar PubMed PubMed Central
Nunes, A.F., Amaral, J.D., Lo, A.C., Fonseca, M.B., Viana, R.S.J., Callaerts-Vegh, Z., D’Hooge, R., and Rodrigues, C.M.P. (2012). TUDCA, a bile acid, attenuates amyloid precursor protein processing and amyloid beta deposition in APP/PS1 mice, Mol. Neurobiol. 45, 440–454.10.1007/s12035-012-8256-ySuche in Google Scholar
Palikaras, K. and Tavernarakis, N. (2014). Mitochondrial homeostasis: the interplay between mitophagy and mitochondrial biogenesis. Exp. Gerontol. 56, 182–188.10.1016/j.exger.2014.01.021Suche in Google Scholar PubMed
Pavshintsev, V.V., Podshivalova, L.S., Frolova, O.Y., Belopolskaya, M.V., Averina, O.A., Kushnir, E.A., Marmiy, N.V., and Lovat, M.L. (2017). Effects of mitochondrial antioxidant SkQ1 on biochemical and behavioral parameters in a parkinsonism model in mice. Biochemistry (Mosc). 82, 1513–1520.10.1134/S0006297917120100Suche in Google Scholar PubMed
Peng, K., Tao, Y., Zhang, J., Wang, J., Ye, F., Dan, G., Zhao, Y., Cai, Y., Zhao, J., Wu, Q., et al. (2016). Resveratrol regulates mitochondrial biogenesis and fission/fusion to attenuate rotenone-induced neurotoxicity. Oxid. Med. Cell Longev. 2016, 6705621.10.1155/2016/6705621Suche in Google Scholar PubMed PubMed Central
Poewe, W., Seppi, K., Tanner, C.M., Halliday, G.M., Brundin, P., Volkmann, J., Schrag, A.E., and Lang, A.E. (2017). Parkinson disease. Nat. Rev. Dis. Primers 3, 17013.10.1038/nrdp.2017.13Suche in Google Scholar PubMed
Postuma, R.B., Anang, J., Pelletier, A., Joseph, L., Moscovich, M., Grimes, D., Furtado, S., Munhoz, R.P., Appel-Cresswell, S., Moro, A., et al. (2017). Caffeine as symptomatic treatment for Parkinson disease (Café-PD): A randomized trial. Neurology 89, 1795–1803.10.1212/WNL.0000000000004568Suche in Google Scholar PubMed PubMed Central
Qi, Y., Qiu, Q., Gu, X., Tian, Y. and Zhanga, Y. (2016). ATM mediates spermidine-induced mitophagy via PINK1 and Parkin regulation in human fibroblasts. Sci Rep. 6, 24700.10.1038/srep24700Suche in Google Scholar PubMed PubMed Central
Rajeswari, A. and Sabesan, M. (2008). Inhibition of monamine oxidase-B by the polyphenolic compound, curcumin and its metabolite tetrahydrocurcumine, in a model of Parkinson’s disease induced by MPTP neurodegeneration in mice. Inflammopharmacology 16, 96–99.10.1007/s10787-007-1614-0Suche in Google Scholar PubMed
Rangaraju, V., Calloway, N., and Ryan, T.A. (2014). Activity-driven local ATP synthesis is required for synaptic function. Cell 156, 825–835.10.1016/j.cell.2013.12.042Suche in Google Scholar PubMed PubMed Central
Ross, G.W., Abbott, R.D., Petrovitch, H., Morens, D.M., Grandinetti, A., Tung, K.H., Tanner, C.M., Masaki, K.H., Blanchette, P.L., Curb, J.D., et al. (2000). Association of coffee and caffeine intake with the risk of Parkinson disease. JAMA 283, 2674–2679.10.1001/jama.283.20.2674Suche in Google Scholar PubMed
Ryu, D., Mouchiroud, L., Andreux, P.A., Katsyuba, E., Moullan, N., Nicolet-Dit-Félix, A.A., Williams, E.G., Jha, P., Lo Sasso, G., Huzard, D., et al. (2016). Urolithin A induces mitophagy and prolongs lifespan in C. elegans and increases muscle function in rodents. Nat. Med. 22, 879–888.10.1038/nm.4132Suche in Google Scholar PubMed
Saravanan, K.S., Sindhu, K.M., and Mohanakumar, K.P. (2007). Melatonin protects against rotenone-induced oxidative stress in a hemiparkinsonian rat model, J. Pineal Res. 42, 247–253.10.1111/j.1600-079X.2006.00412.xSuche in Google Scholar PubMed
Sau, D., De Biasi, S., Vitellaro-Zuccarello, L., Riso, P., Guarnieri, S., Porrini, M., Simeoni, S., Crippa, V., Onesto, E., Palazzolo, I., et al. (2007). Mutation of SOD1 in ALS: a gain of a loss of function. Hum. Mol. Genet. 16, 1604–1618.10.1093/hmg/ddm110Suche in Google Scholar PubMed
Shachar, D.B., Kahana, N., Kampel, V., Warshawsky, A., and Youdim, M.B. (2004). Neuroprotection by a novel brain permeable iron chelator, VK-28, against 6-hydroxydopamine lesion in rats. Neuropharmacology 46, 254–263.10.1016/j.neuropharm.2003.09.005Suche in Google Scholar PubMed
Schapira, A.H., Cooper, J.M., Dexter, D., Clark, J.B., Jenner, P., and Marsden, C.D. (1990). Mitochondrial complex I deficiency in Parkinson’s disease. J. Neurochem. 54, 823–827.10.1111/j.1471-4159.1990.tb02325.xSuche in Google Scholar PubMed
Shepherd, G.M. and Harris, K.M. (1998). Three-dimensional structure and composition of CA3-CA1 axons in rat hippocampal slices: implications for presynaptic connectivity and compartmentalization. J. Neurosci. 18, 8300–8310.10.1523/JNEUROSCI.18-20-08300.1998Suche in Google Scholar
Shults, C.W., Oakes, D., Kieburtz, K., Beal, M.F., Haas, R., Plumb, S., Juncos, J.L., Nutt, J., Shoulson, I., Carter, J., et al. (2002). Effects of coenzyme Q10 in early Parkinson disease: evidence of slowing of the functional decline. Arch. Neurol. 59, 1541–1550.10.1001/archneur.59.10.1541Suche in Google Scholar PubMed
Sies, H., Stahl, W., and Sundquist, A.R. (1992). Antioxidant functions of vitamins. Vitamins E and C, beta-carotene, and other carotenoids. Ann. NY Acad. Sci. 669, 7–20.10.1111/j.1749-6632.1992.tb17085.xSuche in Google Scholar PubMed
Skulachev, M.V., Antonenko, Y.N., Anisimov, V.N., Chernyak, B.V., Cherepanov, D.A., Chistyakov, V.A., Egorov, M.V., Kolosova, N.G., Korshunova, G.A., Lyamzaev, K.G., et al. (2011). Mitochondrial-targeted plastoquinone derivatives. Effect on senescence and acute age-related pathologies. Curr. Drug Targets 12, 800–826.10.2174/138945011795528859Suche in Google Scholar PubMed
Smith, R.A. and Murphy, M.P. (2011). Mitochondria-targeted antioxidants as therapies. Discov. Med. 11, 106–114.10.1201/b12308-8Suche in Google Scholar
Snow, B.J., Rolfe, F.L., Lockhart, M.M., Frampton, C.M., O’Sullivan, J.D., Fung, V., Smith, R.A., Murphy, M.P., and Taylor, K.M. (2010). A double-blind, placebo-controlled study to assess the mitochondria-targeted antioxidant MitoQ as a disease-modifying therapy in Parkinson’s disease. Mov. Disord. 25, 1670–1674.10.1002/mds.23148Suche in Google Scholar PubMed
Starkov, A.A. (2008). The role of mitochondria in reactive oxygen species metabolism and signaling. Ann. NY Acad. Sci. 1147, 37–52.10.1196/annals.1427.015Suche in Google Scholar PubMed PubMed Central
Strappazzon, F., Vietri-Rudan, M., Campello, S., Nazio, F., Florenzano, F., Fimia, G.M., Piacentini, M., Levine, B., and Cecconi, F. (2011). Mitochondrial BCL-2 inhibits AMBRA1-induced autophagy. EMBO J. 30, 1195–1208.10.1038/emboj.2011.49Suche in Google Scholar PubMed PubMed Central
Strappazzon, F., Nazio, F., Corrado, M., Cianfanelli, V., Romagnoli, A., Fimia, G.M., Campello, S., Nardacci, R., Piacentini, M.,Campanella, M., et al. (2015). AMBRA1 is able to induce mitophagy via LC3 binding, regardless of PARKIN and p62/SQSTM1. Cell Death Differ. 22, 517.10.1038/cdd.2014.190Suche in Google Scholar PubMed PubMed Central
Swerdlow, R.H., Burns, J.M., and Khan, S.M. (2014). The Alzheimer’s disease mitochondrial cascade hypothesis: progress and perspectives. Biochim. Biophys. Acta 1842, 1219–1231.10.1016/j.bbadis.2013.09.010Suche in Google Scholar PubMed PubMed Central
Van Humbeeck, C., Cornelissen, T., Hofkens, H., Mandemakers, W., Gevaert, K., De Strooper, B., and Vandenberghe, W. (2011). Parkin interacts with Ambra1 to induce mitophagy. J. Neurosci. 31, 10249–10261.10.1523/JNEUROSCI.1917-11.2011Suche in Google Scholar PubMed PubMed Central
Ved, R., Saha, S., Westlund, B., Perier, C., Burnam, L., Sluder, A., Hoener, M., Rodrigues, C.M., Alfonso, A., Steer, C., et al. (2005). Similar patterns of mitochondrial vulnerability and rescue induced by genetic modification of α-synuclein, parkin, and DJ-1 in Caenorhabditis elegans. J. Biol. Chem. 280, 42655–42668.10.1074/jbc.M505910200Suche in Google Scholar PubMed PubMed Central
Vejrazka, M., Micek, R., and Stipek, S. (2005). Apocynin inhibits NADPH oxidase in phagocytes but stimulates ROS production in non-phagocytic cells. Biochim. Biophys. Acta 1722, 143–147.10.1016/j.bbagen.2004.12.008Suche in Google Scholar PubMed
Wei, Y., Chiang, W.C., Sumpter, R., Jr., Mishra, P., and Levine, B. (2017). Prohibitin 2 is an inner mitochondrial membrane mitophagy receptor. Cell 168, 224–238 e210.10.1016/j.cell.2016.11.042Suche in Google Scholar PubMed PubMed Central
Weinreb, O., Mandel, S., Youdim, M.B.H., and Amit, T. (2013). Targeting dysregulation of brain iron homeostasis in Parkinson’s disease by iron chelators. Free Radic. Biol. Med. 62, 52–64.10.1016/j.freeradbiomed.2013.01.017Suche in Google Scholar PubMed
Wu, W., Tian, W., Hu, Z., Chen, G., Huang, L., Li, W., Zhang, X., Xue, P., Zhou, C., Liu, L., et al. (2014). ULK1 translocates to mitochondria and phosphorylates FUNDC1 to regulate mitophagy. EMBO Rep. 15, 566–575.10.1002/embr.201438501Suche in Google Scholar PubMed PubMed Central
Wu, Z., Wu, A., Dong, J., Sigears, A., and Lu, B. (2018). Grape skin extract improves muscle function and extends lifespan of a Drosophila model of Parkinson’s disease through activation of mitophagy. Exp. Gerontol. 113, 10–17.10.1016/j.exger.2018.09.014Suche in Google Scholar PubMed
Xiao, L., Xu, X., Zhang, F., Wang, M., Xu, Y., Tang, D., Wang, J., Qin, Y., Liu, Y., Tang, C., et al. (2017). The mitochondria-targeted antioxidant MitoQ ameliorated tubular injury mediated by mitophagy in diabetic kidney disease via Nrf2/PINK1. Redox Biol. 11, 297–311.10.1016/j.redox.2016.12.022Suche in Google Scholar PubMed PubMed Central
Xu, Q., Langley, M., Kanthasamy, A.G., and Reddy, M.B. (2017). Epigallocatechin gallate has a neurorescue effect in a mouse model of Parkinson disease. J. Nutr. 147, 1926–1931.10.3945/jn.117.255034Suche in Google Scholar PubMed PubMed Central
Yamano, K., Matsuda, N., and Tanaka, K. (2016). The ubiquitin signal and autophagy: an orchestrated dance leading to mitochondrial degradation. EMBO Rep. 17, 300–316.10.15252/embr.201541486Suche in Google Scholar PubMed PubMed Central
Yang, L., Wang, X., and Yang, X. (2014). Possible antioxidant mechanism of melanoidins extract from Shanxi aged vinegar in mitophagy-dependent and mitophagy-independent pathways. J. Agric. Food Chem. 62, 8616–8622.10.1021/jf501690eSuche in Google Scholar PubMed
Yang, Q., Zheng, C., Cao, J., Cao, G., Shou, P., Lin, L., Velletri, T., Jiang, M., Chen, Q., Han, Y., et al. (2016). Spermidine alleviates experimental autoimmune encephalomyelitis through inducing inhibitory macrophages. Cell Death Differ. 23, 1850–1861.10.1038/cdd.2016.71Suche in Google Scholar PubMed PubMed Central
Zang, Q.S., Sadek, H., Maass, D.L., Martinez, B., Ma, L., Kilgore, J.A., Williams, N.S., Frantz, D.E., Wigginton, J.G., Nwariaku, F.E., et al. (2012). Specific inhibition of mitochondrial oxidative stress suppresses inflammation and improves cardiac function in a rat pneumonia-related sepsis model. Am. J. Physiol. Heart Circ. Physiol. 302, 1847–1859.10.1152/ajpheart.00203.2011Suche in Google Scholar PubMed
Zhang, S.M., Hernan, M.A., Chen, H., Spiegelman, D., Willett, W.C., and Ascherio, A. (2002). Intakes of vitamins E and C, carotenoids, vitamin supplements, and PD risk. Neurology 59, 1161–1169.10.1212/01.WNL.0000028688.75881.12Suche in Google Scholar
Zhang, H., Jia, H., Liu, J., Ao, N., Yan, B., Shen, W., Wang, X., Li, X., Luo, C., and Liu, J. (2010). Combined R-alpha-lipoic acid and acetyl-L-carnitine exerts efficient preventative effects in a cellular model of Parkinson’s disease. J. Cell Mol. Med. 14, 215–225.10.1111/j.1582-4934.2008.00390.xSuche in Google Scholar PubMed PubMed Central
Zheng, H., Weiner, L.M., Bar-Am, O., Epsztejn, S., Cabantchik, Z.I., Warshawsky, A., Youdim, M.B., and Fridkin, M. (2005). Design, synthesis, and evaluation of novel bifunctional iron-chelators as potential agents for neuroprotection in Alzheimer’s, Parkinson’s, and other neurodegenerative diseases. Bioorg. Med. Chem. 13, 773–783.10.1016/j.bmc.2004.10.037Suche in Google Scholar PubMed
Zheng, L., Bernard-Marissal, N., Moullan, N., D’Amico, D., Auwerx, J., Moore, D.J., Knott, G., Aebischer, P., and Schneider, B.L. (2017). Parkin functionally interacts with PGC-1a to preserve mitochondria and protect dopaminergic neurons. Hum. Mol. Genet. 26, 582–598.10.1093/hmg/ddw418Suche in Google Scholar
©2019 Walter de Gruyter GmbH, Berlin/Boston
Artikel in diesem Heft
- Frontmatter
- Evidence for nucleolar dysfunction in Alzheimer’s disease
- National Institutes of Health Stroke Scale, modified Rankin Scale, and modified Thrombolysis in Cerebral Infarction as autonomy predictive tools for stroke patients
- Immune system and new avenues in Parkinson’s disease research and treatment
- Mitophagy could fight Parkinson’s disease through antioxidant action
- Metabolic pattern analysis of 18F-FDG PET as a marker for Parkinson’s disease: a systematic review and meta-analysis
- Pathological and cognitive changes in patients with type 2 diabetes mellitus and comorbid MCI and protective hypoglycemic therapies: a narrative review
- Pain in cervical dystonia and the antinociceptive effects of botulinum toxin: what is currently known?
- N2 amplitude modulation across the antisocial spectrum: a meta-analysis
Artikel in diesem Heft
- Frontmatter
- Evidence for nucleolar dysfunction in Alzheimer’s disease
- National Institutes of Health Stroke Scale, modified Rankin Scale, and modified Thrombolysis in Cerebral Infarction as autonomy predictive tools for stroke patients
- Immune system and new avenues in Parkinson’s disease research and treatment
- Mitophagy could fight Parkinson’s disease through antioxidant action
- Metabolic pattern analysis of 18F-FDG PET as a marker for Parkinson’s disease: a systematic review and meta-analysis
- Pathological and cognitive changes in patients with type 2 diabetes mellitus and comorbid MCI and protective hypoglycemic therapies: a narrative review
- Pain in cervical dystonia and the antinociceptive effects of botulinum toxin: what is currently known?
- N2 amplitude modulation across the antisocial spectrum: a meta-analysis