Chemical and biochemical thermodynamics reunification (IUPAC Technical Report)
-
Antonio Sabatini
 ,
Gerard P. Moss
,
Gerard P. Moss


Abstract
According to the 1994 IUBMB-IUPAC Joint Commission on Biochemical Nomenclature (JCBN) on chemical and biochemical reactions, two categories of thermodynamics, based on different concepts and different formalisms, are established: (i) chemical thermodynamics, which employ conventional thermodynamic potentials to deal with chemical reactions [1], [2], [3]; and (ii) biochemical thermodynamics, which employ transformed thermodynamic quantities to deal with biochemical reactions based on the formalism proposed by Alberty [4], [5], [6], [7]. We showed that the two worlds of chemical and biochemical thermodynamics, which so far have been treated separately, can be reunified within the same thermodynamic framework. The thermodynamics of chemical reactions, in which all species are explicitly considered with their atoms and charge balanced, are compared with the transformed thermodynamics generally used to treat biochemical reactions where atoms and charges are not balanced. The transformed thermodynamic quantities suggested by Alberty are obtained by a mathematical transformation of the usual thermodynamic quantities. The present analysis demonstrates that the transformed values for Δr G′0 and Δr H′0 can be obtained directly, without performing any transformation, by simply writing the chemical reactions with all the pseudoisomers explicitly included and the elements and charges balanced. The appropriate procedures for computing the stoichiometric coefficients for the pseudoisomers are fully explained by means of an example calculation for the biochemical ATP hydrolysis reaction. It is concluded that the analysis reunifies the “two separate worlds” of conventional thermodynamics and transformed thermodynamics.
1 Introduction
In a chemical reaction, all species involved are explicitly considered and their atoms and charges balanced. At equilibrium, the thermodynamic (or standard) equilibrium constant, K, for the chemical reaction is
where
A biochemical reaction involves many species and simultaneous ancillary reactions. Therefore, the determination of the equilibrium composition requires many equilibrium expressions and many conservation equations. A different approach has thus far proved more suitable.
Networks of biochemical reactions take place under steady-state conditions in living organisms. Steady state is the typical condition of the living systems that allows work to be done by chemical reactions at maximal efficiency, or, in other words, to increase entropy at the minimal rate [8]. Another typical feature of living systems is homeostasis, which has the effect of keeping the conditions at which biochemical reactions occur within a narrow range. For example, in a biochemical reaction, the concentration of certain ancillary chemical species, such as H+ and Mg2+ ions, remains essentially constant.
The polyanionic nature of the molecules involved in biochemical reactions implies that these reagents may act as Lewis bases B and react with Lewis acids, such as H+ and Mg2+, to form adducts such as BH+, BMg2+, etc. Consequently, the biochemical reagents are often a mixture of different chemical species. Therefore, biochemical reactions require the use of an equilibrium constant, usually named the apparent equilibrium constant or conditional equilibrium constant. This is the equivalent of an equilibrium constant for a biochemical reaction, where reagents and products are written in terms of the sum of species instead of specific species [4], [5], [6], [7]. It is valid at a given pH, temperature, and ionic strength.
Given the biochemical reaction a A + b B = c C + d D, where A, B, C, and D are biochemical reagents (“sum of species”), the conditional equilibrium constant is as follows (activity coefficients γ i are assumed equal to 1 and are therefore omitted for sake of clarity, see below):
where c
0 is the standard concentration [4], [5], [6], [7] considered equal for all species (1 mol dm−3, according to the “chemical convention”). The value of the conditional equilibrium constant K′ depends, besides T, p, and I, also on pH and pMg (according to IUPAC convention, in this paper pH and pMg are defined as
The amount of H+ and Mg2+ bound to the chemical species of the reactants A and B is different from the amount of these ions bound to the chemical species of the products C and D. Ions H+ and Mg2+ are thus produced or consumed during the course of the reaction. For this reason, chemical and biochemical reactions require a different thermodynamic formulation, since chemical equations are written in terms of specific ionic and elemental species and balance elements and charge, whereas biochemical equations are written in terms of biochemical reactants that consist of species in equilibrium with each other and do not balance elements that are assumed to be fixed, such as H+ and Mg2+. Therefore, when pH and pMg are specified, the conditional equilibrium constant K′ for a biochemical reaction is written in terms of sums of species and can be used to calculate a standard Gibbs energy of reaction Δr G′0, where the prime symbol (′) has been used to indicate that H+ and Mg2+ ions concentrations are constant, but not at the standard value.
For example, in the cytosolic solution, the biochemical reactant ATP (adenosine 5′-triphosphate) is composed of the chemical species ATP4−, HATP3−, MgATP2−, MgHATP−, H2ATP2−, and Mg2ATP. The biochemical reaction of the hydrolysis of ATP is formally written as:
where ADP (adenosine 5′-diphosphate) is understood to be composed of the chemical species ADP3−, HADP2−, H2ADP−, MgADP−, and MgHADP; and Pi (inorganic phosphate) is similarly understood to be composed of the chemical species
Alberty defined pseudoisomers as all the chemical species that form a biochemical reagent [4], [5], [6], [7]. Referring to what is written above, the number of pseudoisomers of ATP are six, while those of ADP are five and those of Pi are four. The complex species are the adducts of these Lewis bases with the Lewis acids H+ and Mg2+. The equilibria between the pseudoisomers of ATP, ADP, and Pi are shown in Fig. 1.
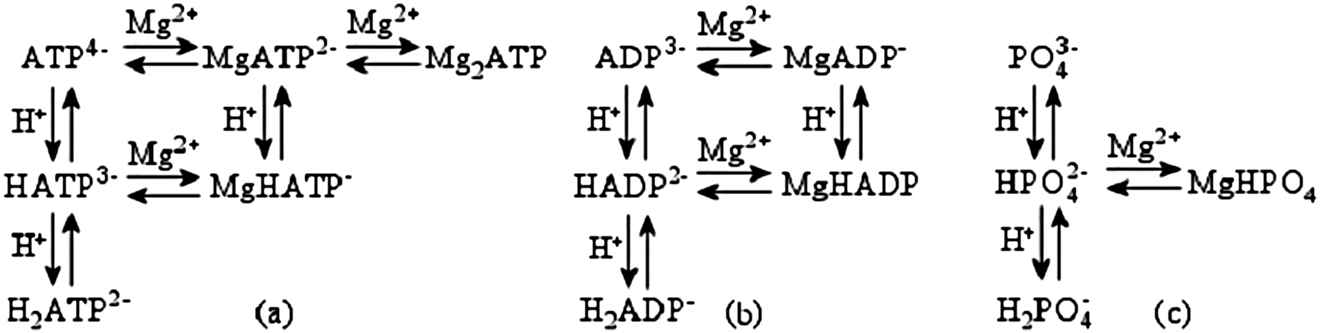
Equilibria between the pseudoisomers of ATP (a), ADP (b) and Pi (c) (from. ref. 16).
However, MgATP2− is the active species in enzyme binding in the cellular active transport and the form responsible for the energy production and muscular contraction [9]. As a consequence, the biologically relevant ATP pseudoisomer is MgATP2− and the chemical reaction related to ATP hydrolysis is:
Taking into account the equilibrium H2PO4 − ⇆ H+ + HPO4 2−, the equilibrium constant for the chemical reaction (3) can be written as:
The value of K depends on temperature T, pressure p, and ionic strength I of the solution. The conditional equilibrium constant for the biochemical reaction (2) is
Besides T, p, and I, its value depends also on pH and pMg. The corresponding values of the standard Gibbs energy of reaction can be obtained from the values of the equilibrium constants:
The Δr G′0, named the standard “conditional” Gibbs energy of reaction, is the reference state Gibbs energy of reaction for the biochemical reaction under the specific experimental condition of pH and pMg.
In the “Recommendations for nomenclature and tables in biochemical thermodynamics”, the IUPAC-IUBMB Joint Commission on Biochemical Nomenclature (JCBN) states [7], “When pH and pMg are specified, a whole new set of transformed thermodynamic properties come into play. These properties are different from the usual Gibbs energy G, enthalpy H, and entropy S and they are referred to as the transformed Gibbs energy G′, transformed enthalpy H′, transformed entropy S’.” As a consequence, two categories of thermodynamics based on different concepts and different formulations were established: (i) chemical thermodynamics, which employ conventional thermodynamic quantities to deal with chemical reactions; and (ii) biochemical thermodynamics, which employ transformed thermodynamic quantities to deal with biochemical reactions.
In his works, Alberty has shown how to obtain the transformed thermodynamic quantities Δr G′0, Δr H′0, and Δr S′0 from Δf G 0, Δf H 0, and Δf S 0 of the specific chemical species [10], [11]. According to Alberty, Δr G and Δr G′ provide the Gibbs energy of the chemical and biochemical reactions, respectively. However, it has been shown that Δr G = Δr G′ regardless of the reaction involved [12]. This fact has not been sufficiently divulged, and it is still not generally well known by the scientific community. It is worth underlining that this equality is restricted to Gibbs energy changes and does not apply to enthalpy or entropy changes.
In addition, it has been subsequently demonstrated that the ‘transformed’ thermodynamic quantities can be obtained without carrying out any ‘transformation’, but simply by considering the chemical potential of the H+ and Mg2+ ions constant in the course of the reaction [13].
This means that the thermodynamic quantities G′, H′, and S′ can also be obtained without any “transformation” and the classification into chemical thermodynamics and biochemical thermodynamics can be misleading. Mathematical clarification of the methods allows the two worlds of chemical and biochemical thermodynamics, which are often treated separately, to be reunified within the same thermodynamic framework. In the next sections, the two different methods will be described in detail.
2 Alberty’s method
According to Alberty, the species formed by a given biochemical reagent constitute a pseudoisomer group [4], [5], [6], [7]. Starting from this consideration, Alberty develops a procedure based on transformed Gibbs energies [4], [7], [10], [11]. Since the chemical species constituting the pseudoisomer group are at equilibrium with H+ and Mg2+ ions (Δμ reaction = 0), i.e.
the chemical potential μ
i
of the ith complex species
where
The chemical potentials,
At a given pH and pMg, the chemical potentials of H+ and Mg2+ ions have a defined value that can be subtracted from the standard potential:
Substituting in Eq. (10), one obtains
The standard transformed chemical potential,
where N
iso is the number of pseudoisomers in the jth isomer group [4], [5], [6], [7]. As a consequence,
3 Thermodynamics of biochemical reactions
An alternative procedure to the one proposed by Alberty for the study of the thermodynamics of a biochemical reaction has been developed and can be used without having to apply transformations of the Gibbs energy [12], [13], [15], [16].
The fundamental equation for the Gibbs energy is given by
where the differential of the amount of species i is dn
i
and N is the number of different kinds of species in the system. When making calculations in chemical thermodynamics, μ
i
can be replaced with Δf
G
i
, the Gibbs energy of formation of species i. At constant T and p, substituting μ
i
with
In a biochemical reaction, however, pH and pMg remain constant and, consequently, their chemical potential remains constant. For this general approach, the considered equilibria are for a “dilute ideal” solution, in which the activity coefficients of all reactants and products are considered equal to one [13], [16]. Although these conditions cannot be expected to hold rigorously in a biological system, this approximation simplifies the following formal development and calculation of the thermodynamic quantities without compromising their validity. The problem of calculating and using the activity coefficients has been extensively discussed elsewhere [12], [17], [18]. Equation (16) can, therefore, be rewritten as follows:
where N − 2 are the chemical species that make up the reactants and products of the biochemical reaction (N = 17 for reaction (2)). The prime symbol (′) indicates that the chemical potential of the H+ and Mg2+ ions has a determined and constant value. By analogy with the terminology used for the equilibrium constant K′ of the biochemical reactions, the Gibbs energy G′ is called “conditional” Gibbs energy.
As reported above, a biochemical reagent is a kind of “sum of chemical species”. Indicating the biochemical reagents with the letter j and the individual chemical species of these biochemical reagents with the letter i, the intrinsic mole fraction f i,j of the chemical species i related to the biochemical reagent j is given by:
where n i,j is the amount of the chemical species i of the biochemical reagent j and n j is the total amount of the biochemical reagent j. For example, referring to the equilibrium (a) in Fig. 1: biochemical reagent: ATP; individual chemical species: ATP4−, HATP3−, H2ATP2−, MgATP2−, MgHATP− and Mg2ATP; n i,j refers to n ATP4− ,ATP , n HATP3− ,ATP , n H 2 ATP2− ,ATP , n MgATP2− ,ATP, n MgHATP− ,ATP , n Mg2ATP,ATP ; n j = n ATP4− ,ATP + n HATP3− ,ATP + n H2ATP2− ,ATP + n MgATP2− ,ATP + n MgHATP− ,ATP + n Mg 2 ATP,ATP. The detailed procedure for the calculation of n i,j is reported in [13]. Placing ν i,j = f i,j or ν i,j = ‒f i,j when the chemical species i is a product or a reagent of the reaction, Δr G′ of a biochemical reaction, when the extent of reaction is 1 mol at constant pH and pMg, is given by
where NB is the number of biochemical reactants and products involved in the biochemical reaction, N(j) is the number of chemical species of the biochemical reagent j,
The standard chemical potential
When the concentrations c j of the biochemical reagents is equal to c 0, Eq. (20) provides the conditional standard Gibbs energy value for the biochemical reaction:
The intrinsic mole fractions f i,j are calculated with a procedure previously described [15]. In the following, X stands for the free species (ATP4−, ADP3−, etc.) of the biochemical reactant X j (ATP, ADP, etc.) and [XH h Mg m ](h+2m)+ for a generic complex species of X (where h and m indicate, respectively, the number of H+ and Mg2+ ions bound to X species; ionic charges from now on are omitted for simplicity). Remembering Eq. (9), one can write the following equality:
Rearranging this equation, one obtains
Substituting
one obtains
For a 1 M solution of the biochemical reactant X, one has:
Dividing both members by c X and taking the reciprocal one obtains:
Once
The concentrations of the chemical species have been calculated for a 1 M solution of the biochemical reagent and their numerical value corresponds to their intrinsic mole fraction.
The stoichiometric coefficients
If the value of
Once the stoichiometric coefficients of all the chemical species participating in the reaction are obtained, the standard enthalpy of reaction can be easily calculated:
4 Thermodynamics of ATP hydrolysis
In this paragraph we report, as a numerical example of application of Eq. (21), the calculation of Δr G′0 for ATP hydrolysis (reaction (2)) at pH = 7 and pMg = 3.
The first column of Table 1 shows the list of the chemical species (reactants and products) involved in the hydrolysis reaction. In the second column the values of Δf
G
0 (T = 298.15 K, I = 0.25 m) of all chemical species implied in the biochemical reaction are reported [7], [11], [13]. At first, we calculate the standard conditional Gibbs energies of formation
The chemical species (reactants and products) involved in the hydrolysis reaction are reported in col. 1. The values reported in the other columns are calculated according to the corresponding equations reported in the text.
| Species | Δf G 0 (kJ mol−1) | N(H+) | N(Mg2+) | Δf G′0 (kJ mol−1) | exp | c = fc 0 (mol dm−3) | Δf G (kJ mol−1) | ν | νΔf G (kJ mol−1) |
|---|---|---|---|---|---|---|---|---|---|
| ATP4− | −2781.06 | 0 | 0 | −2781.06 | 1 | 0.102 48 | −2786.70 | −0.102 48 | 285.57 |
| HATP3− | −2818.77 | 1 | 0 | −2778.00 | 0.291 74 | 0.029 90 | −2827.47 | −0.029 90 | 84.53 |
| H2ATP2− | −2841.42 | 2 | 0 | −2759.89 | 0.000 20 | 0.000 02 | −2868.23 | −0.000 02 | 0.06 |
| MgATP2− | −3261.92 | 0 | 1 | −2786.26 | 8.149 65 | 0.835 14 | −3262.37 | −0.835 14 | 2724.54 |
| MgHATP− | −3288.31 | 1 | 1 | −2771.88 | 0.024 70 | 0.002 53 | −3303.13 | −0.002 53 | 8.36 |
| Mg2ATP | −3729.33 | 0 | 2 | −2778.00 | 0.292 09 | 0.029 93 | −3738.03 | −0.029 93 | 111.89 |
| ADP3− | −1913.42 | 0 | 0 | −1913.42 | 1 | 0.475 51 | −1915.26 | 0.475 51 | −910.73 |
| HADP2− | −1950.34 | 1 | 0 | −1909.57 | 0.212 07 | 0.100 84 | −1956.03 | 0.100 84 | −197.25 |
| H2ADP− | −1972.79 | 2 | 0 | −1891.26 | 0.000 13 | 0.000 06 | −1996.79 | 0.000 06 | −0.12 |
| MgADP− | −2388.78 | 0 | 1 | −1913.12 | 0.885 87 | 0.421 24 | −2390.92 | 0.421 24 | −1007.16 |
| MgHADP | −2416.67 | 1 | 1 | −1900.24 | 0.004 92 | 0.002 34 | −2431.69 | 0.002 34 | −5.69 |
| PO4 3− | −1031.80 | 0 | 0 | −1031.80 | 1 | 0.000 01 | −1059.55 | 0.000 01 | −0.01 |
| HPO4 2− | −1099.34 | 1 | 0 | −1058.57 | 48977.9 | 0.673 48 | −1100.32 | 0.673 48 | −741.05 |
| H2PO4 − | −1138.11 | 2 | 0 | −1056.58 | 21902.0 | 0.301 17 | −1141.08 | 0.301 17 | −343.66 |
| MgHPO4 | −1566.87 | 1 | 1 | −1050.44 | 1842.49 | 0.025 34 | −1575.98 | 0.025 34 | −39.93 |
| H2O | −237.19 | 2 | 0 | −1 | 237.19 | ||||
| H+ | −0.81 | 1 | 0 | 1.00E-07 | −40.77 | 0.62801 | −25.60 | ||
| Mg2+ | −458.54 | 0 | 1 | 1.00E-03 | −475.66 | 0.44862 | −213.39 |
-
Standard state: T = 298.15 K, p = 100 kPa, I = 0.25 M; exp stands for
These values, reported in the last two rows of the table’s column 8, must then be subtracted from the standard Gibbs energies of formation
The stoichiometric numbers,
This chemical equation is written in terms of specific ionic species and balanced elements and charge and it has been named the balanced biochemical reaction [13]. Using the balanced biochemical reaction approach, the calculations needed to obtain Δr
N (H+) =
As already shown [11], at constant pH and pMg, when the chemical species of each biochemical reagent are in equilibrium with each other, the Δr G of any chemical reaction between various chemical species has the same value of the Δr G′ of the biochemical reaction. It would be difficult to measure the concentration of the single chemical species involved in the energy-producing chemical reaction, while it is relatively easy to measure the total concentration of the biochemical reactants.
In the case of ATP hydrolysis, MgATP2− is the active species [9] and, as a consequence, the actual reaction of ATP hydrolysis, in vivo, is the chemical reaction (3). In the muscle cytosol, we observe [9]:
Using the stoichiometric numbers, ν, reported in Table 1 and the procedure reported in detail in the reference publication [12], we obtain:
The Δr G of the chemical reaction (3) at T = 298.15 K, p = 105 Pa, I = 0.25 M, pH = 7 and pMg = 3 is:
At the same conditions of T, p, I, pH, and pMg, the Δr G′ of the biochemical reaction (2) is
The assertion that the Δr G of any chemical reaction between various chemical species has the same value of the Δr G′ of the biochemical reaction is verified. It is worth underlining that it has also been demonstrated that Δr G can be calculated either in terms of activities or concentrations, giving the same value, under the assumption that all activity coefficients are constant [12].
5 Conclusions
The presence of two categories of thermodynamics based on different concepts and different formalisms has often been presumed in the literature: (i) chemical thermodynamics, which make use of the conventional thermodynamic properties and is only suitable to deal with chemical reactions; and (ii) biochemical thermodynamics, which make use of transformed thermodynamic properties and is only suitable to deal with biochemical reactions.
The complexity of biochemical reactions entails a conceptual and experimental problem to determine experimentally the energy released by a specific enzymatic reaction and to assess the associated thermodynamic properties.
The analysis above demonstrates that biochemical reactions, which occur at constant pH and pMg, can be conveniently described by conventional thermodynamics [16]. The two worlds of chemical and biochemical thermodynamics should be viewed as being within the same consistent thermodynamic framework.
Biochemists and analytical chemists are used to dealing with chemical equilibria occurring in a solution at constant pH and having reagents that are the “sum of species”. The corresponding equilibrium constant, K′, is referred to as conditional [19], [20] whereas ΔrG′, ΔrH′, and ΔrS′ have been so far termed, according to Alberty, transformed thermodynamics properties. We suggest the unification of the terminology using the appellation conditional for both K′ and ΔrG′, ΔrH′, and ΔrS′.
The prime symbol is used to indicate that the equilibrium constant and the thermodynamic properties refer to reactions that occur, not only at constant T, p, and I, but also at certain fixed pH and pMg values. The term transformed thermodynamics should be abolished and conditional thermodynamics used instead.
Membership of the sponsoring body
Membership of the IUPAC Physical and Biophysical Chemistry Division for the period 2018–2019 was as follows:
President: Ronald Weir (Canada); Past President: Angela K. Wilson (USA); Vice President: Tim Wallington (USA); Secretary: Attila Császár (Hungary); Titular Members: Frances Separovic (Australia); Roberto Marquardt (France); Pierangelo Metrangolo (Italy); Hiroko Tokoro (Japan); Bert Weckhuysen (Netherlands); Jeremy G. Frey (UK); Associate Members: Zhigang Shuai (China); Vladislav Tomišic (Croatia); Theo Christian Kurtén (Finland); Modou Fall (Senegal); Michal Korenko (Slovakia); National Representatives: Ioanna Zoi (USA); Chia-Chun Jay Chen (China/Taipei); Joaquim Luís Bernades Martins de Faria (Portugal); Leticia Gonzalez (Austria); Keith C. Gordon (New Zealand); Seung-Joon Jeon (South Korea); Gamini Rajapakse (Sri Lanka); Vessela Tsakova (Bulgaria); Ilya Vorotyntsev (Russia); Malgorzata Witko (Poland).
Acknowledgments
We would like to remember Antonio Sabatini, who passed away unexpectedly during the final preparation of the manuscript. Antonio had been working on this project until the last minute. He was very enthusiastic about the project and very active in its completion. He was really a great scientist and a great man.
-
Research funding: This work was prepared under project 2017-021-2-100 of IUPAC (Funder ID: 10.13039/100006987).
References
[1] M. B. Ewing, T. H. Lilley, G. M. Olofsson, M. T. Ratzsch, G. Somsen. Pure Appl. Chem. 66, 533–552 (1994). https://doi.org/10.1351/pac199466030533.Search in Google Scholar
[2] IUPAC Green Book-Physical Chemistry Division. Quantities, Units and Symbols in Physical Chemistry, Blackwell Scientific Publications, Oxford (1988), 2nd ed. (1993).Search in Google Scholar
[3] IUPAC-Physical Chemistry Division. Manual of symbols and terminology for physicochemical quantities and units. Appendix IV, Pure Appl. Chem. 54, 1239–1250 (1982).Search in Google Scholar
[4] R. A. Alberty. Pure Appl. Chem. 66, 1641–1666 (1994). Revised by IUBMB in R. A. Alberty, A. Cornish-Bowden, R. N. Goldberg, G. G. Hammes, K. Tipton, H. V. Westerhoff. Biophys. Chem. 155, 89–203 (2011). https://doi.org/10.1351/pac199466081641.Search in Google Scholar
[5] R. A. Alberty. Biophys. J. 65, 1243–1254 (1993). https://doi.org/10.1016/s0006-3495(93)81146-9.Search in Google Scholar PubMed PubMed Central
[6] K. C. Vinnakota, F. Wu, M. J. Kushmerick, D. A. Beard. Methods Enzymol. 454, 29–68 (2009). https://doi.org/10.1016/s0076-6879(08)03802-0.Search in Google Scholar
[7] G. P. Moss (1994) IUBMB-IUPAC Joint commission on Biochemical Nomenclature (JCBN), recommendations for nomenclature and tables in biochemical thermodynamics. Available: http://www.chem.qmul.ac.uk/iubmb/thermod/.Search in Google Scholar
[8] A. Katchalsky, P. F. Curran. Non Equilibrium Thermodynamics in Biophysics, Harvard University Press, Cambridge, Massachusetts (1965).10.4159/harvard.9780674494121Search in Google Scholar
[9] S. Iotti, C. Frassineti, A. Sabatini, A. Vacca, B. Barbiroli. Biochim. Biophys. Acta 1708, 164–177 (2005). https://doi.org/10.1016/j.bbabio.2005.01.008.Search in Google Scholar PubMed
[10] R. A. Alberty, Biophys. Chem. 42, 117–131 (1992). https://doi.org/10.1016/0301-4622(92)85002-l.Search in Google Scholar PubMed
[11] R. A. Alberty, Biophys. Chem. 43, 239–254 (1992). https://doi.org/10.1016/0301-4622(92)85024-x.Search in Google Scholar
[12] S. Iotti, A. Sabatini, A. Vacca. J. Phys. Chem. B 114, 1985–1993 (2010). https://doi.org/10.1021/jp903990j.Search in Google Scholar PubMed
[13] A. Sabatini, A. Vacca, S. Iotti. PLoS One 7, e29529 (2012). https://doi.org/10.1371/journal.pone.0029529.Search in Google Scholar PubMed PubMed Central
[14] R. A. Alberty. Biochim. Biophys. Acta 1207, 1–11 (1994). https://doi.org/10.1016/0167-4838(94)90045-0.Search in Google Scholar PubMed
[15] A. Sabatini, M. Borsari M, L. M. Raff, W. R. Cannon, S Iotti. Chem. Int. 41(2), (2019). https://doi.org/10.1515/ci-2019-0211. http://www.iupac.org/project/2017-021-2-100.Search in Google Scholar
[16] S. Iotti, L. Raff, A. Sabatini. Biophys. Chem. 221, 49–57 (2017). https://doi.org/10.1016/j.bpc.2016.10.004.Search in Google Scholar PubMed
[17] P. Hoffmann, T. Maskow, C. Held, G. Sadowski. A thermodynamic investigation of the glucose-6-phosphate isomerization, Biophys. Chem. 195 (2014) 22–31. https://doi.org/10.1016/j.bpc.2014.08.002.Search in Google Scholar PubMed
[18] C. Held, G. Sadowski. Thermodynamics of Bioreactions, Ann. Rev. Chem. Biomol. Eng. 7, 395–414 (2016). https://doi.org/10.1146/annurev-chembioeng-080615-034704.Search in Google Scholar PubMed
[19] A. Ringbom. J. Chem. Ed. 35, 282–288 (1958). https://doi.org/10.1021/ed035p282.Search in Google Scholar
[20] J.-L. Burgot. Ionic Equilibria in Analytical Chemistry, Springer, New York (2012).10.1007/978-1-4419-8382-4Search in Google Scholar
© 2020 IUPAC & De Gruyter. This work is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License. For more information, please visit: http://creativecommons.org/licenses/by-nc-nd/4.0/
Articles in the same Issue
- Frontmatter
- In this issue
- Preface
- Winners of the 2020 IUPAC-SOLVAY International Award for Young Chemists
- Invited papers (IUPAC-Solvay Award)
- Transition metal catalyzed site-selective cysteine diversification of proteins
- Antioxidant metal oxide nanozymes: role in cellular redox homeostasis and therapeutics
- Invited papers (Diamond Jubilee Issue)
- Narratives of undergraduate research, mentorship, and teaching at UCLA
- Hot-carriers in organic photovoltaics
- Invited paper (Special Topic)
- Natural clay nanotube supported Mo and W catalysts for exhaustive oxidative desulfurization of model fuels
- IUPAC Technical Reports
- Chemical and biochemical thermodynamics reunification (IUPAC Technical Report)
- Good reporting practice for thermophysical and thermochemical property measurements (IUPAC Technical Report)
Articles in the same Issue
- Frontmatter
- In this issue
- Preface
- Winners of the 2020 IUPAC-SOLVAY International Award for Young Chemists
- Invited papers (IUPAC-Solvay Award)
- Transition metal catalyzed site-selective cysteine diversification of proteins
- Antioxidant metal oxide nanozymes: role in cellular redox homeostasis and therapeutics
- Invited papers (Diamond Jubilee Issue)
- Narratives of undergraduate research, mentorship, and teaching at UCLA
- Hot-carriers in organic photovoltaics
- Invited paper (Special Topic)
- Natural clay nanotube supported Mo and W catalysts for exhaustive oxidative desulfurization of model fuels
- IUPAC Technical Reports
- Chemical and biochemical thermodynamics reunification (IUPAC Technical Report)
- Good reporting practice for thermophysical and thermochemical property measurements (IUPAC Technical Report)