Analyzing polymeric nanofibrous scaffold performances in diabetic animal models for translational chronic wound healing research
-
Nowsheen Goonoo
Nowsheen Goonoo holds a PhD in Polymers and Biomaterials and is currently a Postdoctoral Researcher at the Biomaterials, Drug Delivery and Nanotechnology Unit, Center for Biomedical and Biomaterials Research. She was awarded the
Georg Forster Postdoctoral Fellow , Alexander Von Humboldt Foundation (AvH) in 2015, and spent 1 year at the University of Siegen, Germany. She was granted aReturn Fellowship for Research Fellows (AvH) in August 2017. Her research interests include cell-scaffold interactions and polymeric biomaterials for biomedical applications.Archana Bhaw-Luximon holds a PhD in Polymer Chemistry and is currently an Associate Professor and Unit Leader of the Biomaterials, Drug Delivery and Nanotechnology Unit, Center for Biomedical and Biomaterials Research. The research thrusts of her Unit are design of polymeric biomaterials for fabrication of scaffolds for skin and bone tissue regeneration, engineering nanocarriers for anti-cancer and anti-diabetic drugs/biomolecules, and exploitation of materials from land and marine resources such as sugarcane bagasse and seaweeds for high-end applications.


Abstract
Tissue-engineered scaffolds for skin wound healing have undergone tremendous progress since the advent of autologous skin grafts or autografts. The recognition that a three-dimensional (3D) scaffold more closely mimics the biochemical-mechanical milieu of wounds and advancing knowledge of cell biology has led to the next-generation of engineered biopolymeric nanoscaffolds. These can pave the way towards personalized wound care as they can address multiple requirements of skin physiology. A unifying approach is required for translational success of scaffolds, involving clinicians, biologists, and chemists. Following in vitro testing using cell lines, pre-clinical studies on animal models is the next crucial step towards scaffold clinical translation. Often, success noted in these studies does not replicate in human patients due to the heterogeneous nature of wound conditions and causes that cannot be found in a single animal model. In this review, we give an overview of the status of pre-clinical mice models for types 1 and 2 diabetic wound healing and relate this to studies conducted with polymeric nanoscaffolds. Translational shortcomings of these models are analyzed where they fail to provide a good replica of human conditions. Categorization and use of more sophisticated models, which can closely mimic the diabetic features, are addressed for translational use of nanoscaffolds.
Abbreviations: ALX, alloxan; BM, basement membrane; DFU, diabetic foot ulcer; ECM, extracellular matrix; eNOS, endothelial nitric oxide synthase; EPCs, endothelial progenitor cells; FGF, fibroblast growth factor; NP, nanoparticle; NZO, New Zealand obese; ORC, oxidized regenerated cellulose; PA, peptide amphiphile; PDGF, platelet-derived growth factor; STZ, streptozotocin; VEGF, vascular endothelial growth factor.
1 Introduction
Translational wound healing research is faced with the difficulty of having to address the heterogeneous nature of wounds, more so, in the case of chronic wounds such as diabetic foot ulcers (DFUs) and pressure ulcers, which often have an unexpected sequence of events [1]. Clinicians are faced with challenges such as age of patients, hormone levels, microbiota, and the various causes of wounds. The available wound care options do not necessarily categorize patients according to clinical requirements, and this is where reverse translational strategies become important to explore wound healing matrices. Developing and adapting animal models appropriate to the diversity of clinical setting become crucial. Many criticisms about animal models in pre-clinical research have been raised due to failure to translate to patients. The advent of new treatment in wound healing such as polymeric nanofiber scaffolds further demands carefully chosen models for translational success. In this context, electrospinning is considered as an efficient method for fabricating biomimicking polymeric fibers with tunable fiber diameters [2]. It also offers the possibility of developing multifunctional formulations via the combination of fibers with nanocarriers such as micelles, nanoparticles (NPs), etc., which, in turn, allows sustained release of drugs/growth factors to enhance tissue repair.
Extreme loss of skin function and structure due to burns, injury, or illness exposes the body to a risk of systemic infection and can cause significant physiological imbalance, which may eventually become life threatening [3]. Indeed, the skin forms an important physical and chemical interface protecting the host against dehydration, toxins, and invading microorganisms [4]. The standard clinical procedure for the treatment of large full-thickness skin defects is autologous skin grafts or autografts, which are obtained from the patients, themselves [5], [6]. However, autografts are limited and often lead to scarring in patients [7], [8]. Skin harvested from other individuals, i.e. allografts present several ethical problems and are influenced by availability and potential disease transmission [7]. During the past few years, progress in the field of tissue engineering has led to the design and commercialization of several tissue-engineered products for skin regeneration. The latter may be classified according to the anatomical structure to be regenerated, namely, epidermal, dermal, or dermal/epidermal (or composite) substitutes. Epidermal skin substitutes available for clinical applications may consist of cells of either autologous or allogenic origin, for example, Epicel® (Genzyme Biosurgery, USA) or MySkin® (CellTran Ltd, UK), respectively [9]. However, despite efficient epidermal coverage, autologous and allogenic epidermal substitutes result in poor attachment rates, which, in turn, lead to blister formation [10]. The lack of dermal tissue in full thickness wounds and the poor quality of scars following treatment with autografts led to the development of dermal substitutes such as Dermagraft® (Shire Regenerative Medicine, Inc, USA), Integra™ (Integra LifeSciences, USA), Karoderm™ (Karocell Tissue Engineering AB company, Sweden), etc. [11]. In an attempt to mimic the skin layers (dermis and epidermis) within the same construct, dermal/epidermal substitutes were fabricated, for instance, PermaDerm® (Regenicin Inc., USA) and Apligraf® (Novartis, USA). Despite enormous progress achieved on TE-based skin substitutes, several challenges still need to be overcome to achieve the optimal skin substitute, in particular, improved adhesion of cultured keratinocytes to the wound bed, enhanced rate of neovascularization of tissue-engineered skin, and better resistance of scaffolds materials towards wound contraction and fibrosis [5], [12], [13]. These drawbacks have opened avenues for designing advanced skin substitutes, which involve combining scaffolds, cells, and growth factors. Nanostructured scaffolds provide numerous advantages, such as three-dimensional (3D) micro environment, tailored mechanical and biological properties. Nano-engineered biomaterials in the form of NPs, nanofibers, or a combination of both have been designed for skin regeneration [4]. These scaffolds often perform well during the classical in vitro tests using cell lines such as fibroblasts, keratinocytes, among others. Subsequent in vivo experiments are often carried out on a chosen animal model to address one specific problem such as impaired wound healing in diabetic patients. A single in vivo model is, therefore, very often not suitable to conclude on the performance of the scaffold as most animal models do not replicate the human physiological conditions closely.
In this review, we outline current diabetic animal models used for in vivo testing of polymeric scaffolds and suggest ways to come up with avenues to correlate polymeric scaffold performance with in vivo models, which can facilitate both reverse translation and translational research. We point out that a synergy is required between polymer experts, biologists, and clinicians.
2 Skin: physiology and regeneration
The skin consists mainly of two morphologically distinct layers, namely, the epidermis and the dermis. The epidermis, which is the top thin avascular layer (0.1–0.2 mm in depth and nourished through diffusion from the dermis), comprises mainly of a self-renewing population of keratinocytes and is critical for survival as it acts as a barrier protecting the body against exogenous substances, chemicals, pathogens, and prevents dehydration through the regulation of fluid loss [14]. Underlying the epidermis is the dermis, which is a thicker vascularized layer (approx. 2 mm) of connective tissues consisting mainly of extracellular matrix (ECM), structural components such as collagen and elastin, and a vascular network [15], [16]. The nanoscale fibrillar ECM network is highly dynamic and plays an important role in hydration and in maintaining moisture levels. It also has a prominent role in directing cellular behavior through direct and indirect signaling [9]. Cells including fibroblasts, endothelial cells, smooth muscle cells, and mast cells are interdispersed within the ECM [15]. The epidermis is securely attached to the collagen fibers of the dermis by the complex basement membrane (BM) [17]. A collection of adhesion receptors extends through the BM and acts as anchoring filaments binding keratinocytes from the epidermis to the lower BM. Epidermal/dermal communication across various levels are essential to establish, maintain, or restore tissue homeostasis.
Following skin damage from burns, contusions, hematomas, or diseases, a consecutive cascade of events, i.e. wound healing takes place to restore the skin structure and function [18]. The wound healing process is characterized by three different phases, namely, inflammation, proliferation, and remodeling, which occur sequentially following skin damage [18]. Disruption of the epidermal layer immediately triggers keratinocytes and platelets to release cytokines like interleukins leading to the activation of the blood clotting cascade.
The release of vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF) by platelets stimulate the proliferation of endothelial cells initiating angiogenesis, which is crucial for the synthesis, deposition, and organization of a new ECM [19]. The presence of growth factors TGF-β1 and platelet-derived growth factor (PDGF) attracts fibroblasts into the wound site as well as initiates phenotypic changes in these cells converting fibroblasts into myofibroblasts. These cells then secrete large amounts of ECM and align themselves along the borders of the wound to generate a constrictive force, facilitating wound closure [20]. Fibroblasts in the wound site then secrete a wide range of molecules including growth factors and ECM proteins like VEGF, dermatopontin (DPT), and collagen that regulate the proliferation and migration of fibroblasts, keratinocytes, and endothelial cells [21].
The next phases, namely, migration and proliferation, are interdependent [18], [22]. Migration is characterized by infiltration of new epithelial cells into the damaged area to replace dead cells. The proliferation phase involves simultaneous movement of fibroblasts and endothelial cells to the damaged area forming a granular tissue composed of a new matrix and blood vessels [23].
The last phase, maturation, comprises the remodeling process, during which fibroblasts cover the damaged skin surface with a new skin layer and ideally leaving no evidence of scar [1]. In the final remodeling phase of wound healing, a network of collagen and elastin fibers, together with proteoglycans and glycoproteins, replaces the granulation tissue and initiates the maintenance of balance between matrix production and degradation [4].
2.1 Skin substitutes
2.1.1 Skin defects and wounds
Depending on the depth of the skin defect, different measures need to be taken. Injuries involving the epidermis, as well as those extending into the superficial layer of the dermis (partial thickness injuries), will heal naturally without surgical intervention, provided that there are sufficient keratinocyte stem cells present in the remaining epidermis or in the residual dermis [24]. However, if this is not the case, then, skin regeneration may be achieved using epithelial stem cells derived from hair follicles and/or sweat glands [24]. Defects extending into the deeper dermis require a more complex treatment, due to the absence of keratinocyte stem cells. The “gold standard” approach for the treatment of such wounds is to apply split-thickness skin grafts (taken from elsewhere in the body) that contain all epidermal and marginal parts of the dermis, thereby, transferring self-renewing keratinocyte stem cells to the affected area. However, if the wound injury is extensive, it is often necessary to “build up” the skin in two stages whereby one material is used to provide a dermal equivalent, which must become vascularized before an epidermal material (most often a very thin split-thickness graft, but occasionally cultured cells or tissue-engineered skin) may be placed on top [14].
Scarring or wound contraction is a crucial hallmark of skin reconstitution [24]. The body must produce new collagen fibers to close the wound. However, the body cannot re-build the tissue exactly as it was, and thus, the new (scar) tissue has a distinct collagen pattern and a different skin texture (and quality) than the surrounding normal skin.
Overall, successful skin reconstruction with skin substitutes depends on the presence of self-renewing keratinocyte stem cells for re-epithelialization and a functional dermal substitute consisting of the appropriate cellular and acellular components that allow no or only limited scarring of the developing skin [24].
2.1.2 Commercially available skin substitutes
Skin substitutes refer to wound coverage materials that help in would closure and replace the functions of the skin, either temporarily or permanently, depending on the product characteristics [25]. The ideal substitute should allow immediate replacement of both the lost dermis and epidermis, and should be permanent. In addition, it should be cost effective, have a long shelf-life and easy storage conditions, durable with long-term wound stability, allow easy handling and application, and be able to withstand the shear forces and infection [26].
Temporary skin substitutes provide transient physiologic wound closure, with protection from mechanical trauma, and act as a physical barrier to bacteria and create a moist wound environment. Such substitutes are used to facilitate epithelialization and control pain, as dressing on clean superficial wounds until epithelialization, and to provide temporary physiological closure of deep wounds after excision while awaiting autografting [26]. Permanent substitutes, on the other hand, achieve permanent wound closure. Skin substitutes may be cellular autogeneic (cells derived from the patient), cellular allogeneic, or acellular (deprived of cells). Several skin substitutes are available on the market for epidermal, dermal, or epidermal/dermal replacement, and each has advantages and limitations [27], [28]. Tissue-engineered skin refers to a material made up of cells, ECM, or a combination of both. Most of the commercially available skin substitutes provide only temporary coverage of skin surfaces and need to be replaced later. An extensive review of skin substitutes has been described by Debels et al. [29].
3 In vivo models for diabetic ulcers
Despite the availability of a number of skin replacement options, there is no ideal skin substitute available, to date, that meets all the required criteria of the wound healing process in humans. Indeed, the design of a bioengineered skin replicating the essential physiological characteristics for replacing diseased or damaged parts remains a real challenge. Current efforts are being focused on the fabrication of biodegradable and biocompatible nanomaterials that structurally and functionally mimic the natural skin environment. As discussed in several reviews, various nanotherapeutic approaches are being investigated for wound healing, in particular, NPs and scaffolds, which target the main phases of wound repair (inflammation, remodeling, and wound contraction) [30], [31], [32], [33], [34]. Before being tested clinically, these materials have to be tested in vitro and, more importantly, in vivo such that the outcome on humans may be better predicted. As a result, appropriate animal models need to be selected depending on the wound healing situation being investigated. Animal models need to mimic, as closely as possible, the physiological conditions and characteristics of healthy or diseased wounds as in humans. Hence, wise and careful choice of suitable animal models is a top priority for proper data interpretation. In this section, the benefits and limitations of current animal-based models will be discussed with particular highlight on the urgent need for improved predictive pre-clinical models for wound healing research.
Diabetes is a chronic disease that occurs either when the pancreas does not produce enough insulin or when the body cannot effectively use the insulin it produces. There are two main types of diabetes, namely, type 1 diabetes and type 2 diabetes. Type 1 diabetes is due to an autoimmune destruction of the insulin-producing pancreatic β cells, and type 2 diabetes, which is the most common type, is caused by insulin resistance coupled by a failure of the β cell to compensate [35]. Diabetes is a major worldwide health problem, and according to WHO, there were 422 million people suffering from diabetes in 2014 [36]. A major complication of diabetes is impaired healing, which often leads to chronic wounds and limb amputation [37]. DFUs are the leading cause of non-traumatic lower limb amputations [38]. WHO projects that diabetes will be the seventh leading cause of death by 2030 [39]. Factors contributing to wound healing deficiency in diabetic patients include altered host response, diminished antibacterial defenses, prolonged inflammation, altered protease activity, a tendency for vascular abnormalities, the generation of an inadequate number of cells to accomplish rapid and robust healing, decreased growth factor production, inability to form a sufficient amount of ECM, and alterations in apoptosis [40], [41].
The main differences in the wound healing process of healthy individuals and those suffering from diabetes are depicted in Figure 1. The acute wound healing process in healthy persons is guided and maintained through integration of multiple signals in the form of cytokines and chemokines released by a wide range of cells including keratinocytes, fibroblasts, endothelial cells, macrophages, and platelets. During wound-induced hypoxia, VEGF released by macrophages, fibroblasts, and epithelial cells induces the phosphorylation and activation of eNOS in the bone marrow, leading to an increase in NO levels, which, in turn, triggers the mobilization of bone marrow endothelial progenitor cells (EPCs) to the circulation. The chemokine SDF-1α promotes the homing of these EPCs to the wound site, where they participate in neovasculogenesis. On the other hand, in diabetic patients (Figure 1, right), endothelial nitric oxide synthase (eNOS) phosphorylation in the bone marrow is impaired, which directly limits the mobilization of EPCs from the bone marrow into the circulation [42]. SDF-1α expression in epithelial cells and myofibroblasts is decreased in the diabetic wound, which prevents EPC homing to wounds, thereby, resulting in impaired wound healing.

Mechanisms of wound healing in healthy vs. diabetic patients.
In diabetes, the different phases of wound healing do not occur in synchrony, due to the complications of the disease. The pathophysiology of diabetes-related wound healing problems is complex and poorly understood. The use of animal models has provided valuable insights into the pathogenesis of both diabetes and chronic wounds. Indeed, the wound healing potential may be quantified by the analyses of re-epithelialization, granulation tissue formation, and blood vessel density, as well as VEGF and inflammatory response measurements. However, there are only a few published reports, to date, whereby the wound healing differences among differing diabetic animal models have been investigated. Most of the available models are based on rodents due to their small size, short generation time, easy availability, and economic considerations [35]. These models, thus, do not necessarily replicate the human conditions.
3.1 Animal models for type I diabetes
Animal models for type 1 diabetes range from animals with spontaneously developing autoimmune diabetes to chemical ablation of the pancreatic β cells [35]. The most commonly used models of type 1 diabetes and their limitations are outlined in Scheme 1. The non-existence of standard protocols, different mechanism of diabetes development in human vs. animal models, and the possibility of reversible diabetes are some of the problems identified in these models.
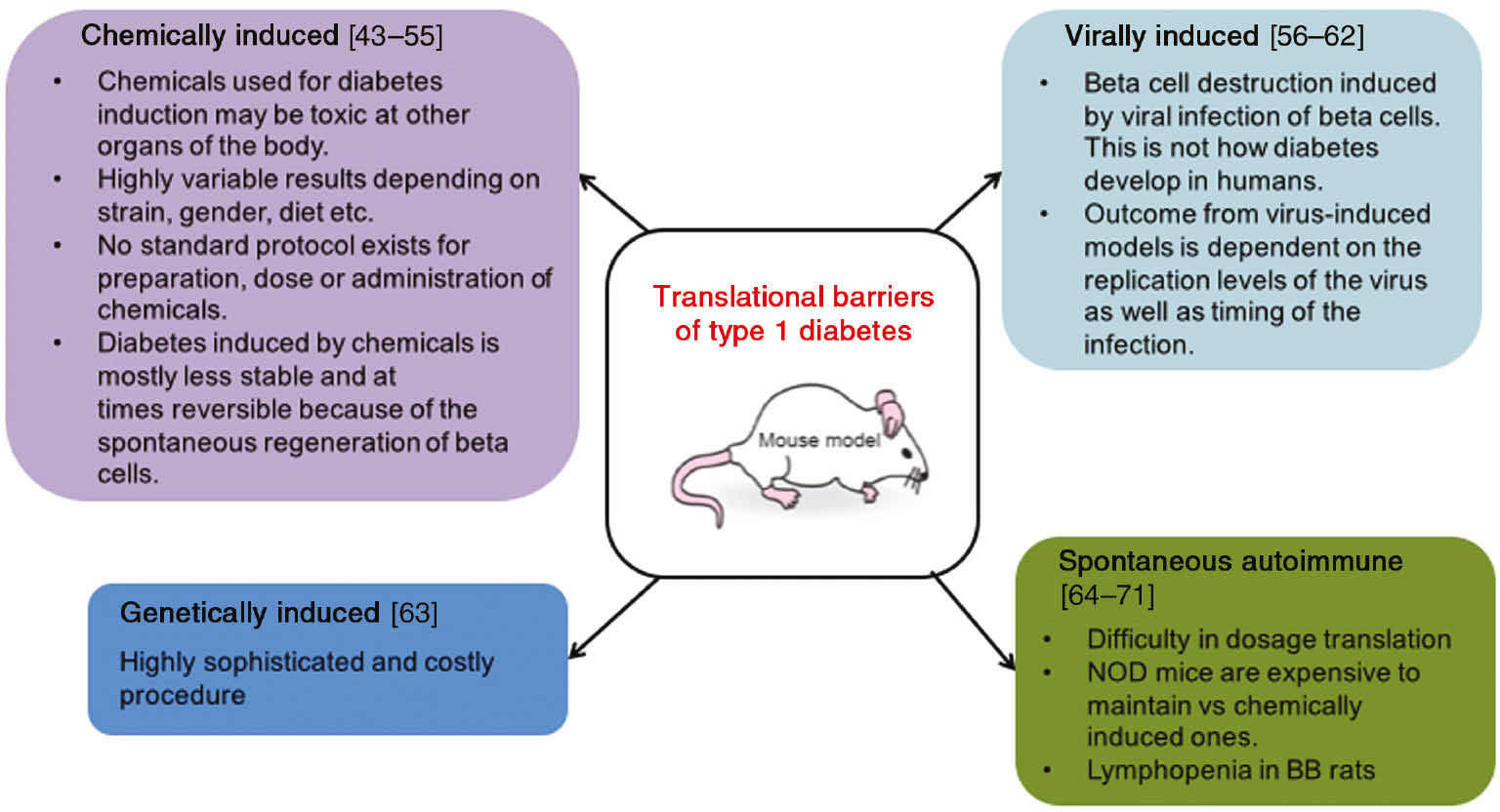
Translational barriers of type 1 mice models.
In chemically induced models of diabetes, a high percentage of the endogenous β cells are destroyed using cytotoxic glucose analogs such as streptozotocin (STZ) or alloxan (ALX), and thus, there is little endogenous insulin production, leading to hyperglycemia and weight loss. This method is versatile and can be used to induce type 1 or type 2 diabetes [43], [72]. STZ [2-deoxy-2-(3-(methyl-3-nitrosoureido)-d-glucopyranose] is synthesized by Streptomycetes achromogenes. It enters the pancreatic β cell through the Glut-2 glucose transporter after i.p. or i.v. injections and causes alkylation of the DNA [44], which, in turn, activates PARP leading to NAD+ depletion, a reduction in cellular ATP, and subsequent inhibition of insulin production [45]. On the other hand, ALX (2,4,5,6-tetraoxypyrimidine; 5,6-dioxyuracil) generates the reactive oxygen species (ROS) in a cyclic redox reaction with its reduction product, which contributes to DNA damage and eventually cell death [46]. STZ may be administered as a single high dose or as multiple low doses, and the dose can vary greatly depending on the animal gender and strain. Multiple low-dose STZs have been most currently used for wound healing studies involving polymeric scaffolds (Table 1). An analysis of the results obtained indicates that translation from mice to humans faces many barriers apart from the fact that the skin of mice is physiologically, morphologically, and functionally different from the human skin. Diabetic wounds are usually classified using a wound classification system. However, there are various classification systems which are in use, the Wagner’s and University of Texas being the two well-established ones [73]. Wagner used wound depth, and the University of Texas added infection and ischemia as the two other principal factors to be taken into consideration. These factors are heterogeneous and vary according to patient. Mice studies do not necessarily make use of this classification to mimic the diabetic wound condition. Adding this parameter would enhance translational capacity of polymeric scaffolds.
Summary of main features of polymeric scaffolds and their resulting performances in STZ-induced diabetic rodent models.
| Polymeric scaffold and main features | In vivo performance of scaffolds compared to control | Translation challenges to humans with type 1 diabetes |
|---|---|---|
| Nanofibers from self-assembled peptide amphiphiles (PA) [74] – Average diameters 10−20 nm, and lengths up to several micrometers – CD revealed β-sheet structure | – Higher wound closure rates (75% vs. 65% on day 14) – Acceleration in re-epithelialization The thickness of PA-treated epidermis was significantly greater – Granulation tissue per total wound area was 58% in the PA-treated group vs. 45% for control group – Thicker and more-organized collagen fibrils and bundles noted in the granulation tissue – Higher number of myofibroblasts at the wound margins – Enhanced neovascularization | – The skin of mice is physiologically, morphologically, and functionally different from human skin [76] – Not clear whether results obtained can be extrapolated to more advanced, larger lesions (>6-mm diameters). – High % of persons in randomized trials showing complete wound closure needed |
| Curcumin-loaded chitosan nanoparticles (NPs) within a collagen/alginate matrix [75] – Size of NPs: approximately 196.4 nm – CUR was in the amorphous state within the NPs – Porous architecture with a geometry ranging from 50 to 250 μm – CUR release of 94.66±5.23% from the nanohybrid scaffold at 360 h | – Mean percentage of wound contraction of the nanohybrid scaffold-treated group was 98.1 ± 3.4% vs. 44.6 ± 6.3% at day 15 – Well-formed thick granulation tissue with fibroblasts with negligible inflammatory cells and a new epithelial layer in the nanohybrid scaffold treated group while inflammatory cells were also present and the formation of granulation tissue was not evident in the control group | – The percent of wound area reduction at 1 and 4 weeks was proposed as predictor of healing at 16 weeks [77]. This study was carried out for 2 weeks only – Studies should also include non-healing DFUs (DFUs: wound area reduction less than 53% after 4 weeks) |
| Allogeneic non-diabetic adipose-derived stem cells (ADSCs) encapsulated within Pluronic F127 hydrogel [79] | – Accelerated wound closure accompanied by facilitated regeneration of granulation tissue – Enhanced angiogenesis (CD31 marker) and cell proliferation (Ki67 marker) at the wound site with upregulation of VEGF and the key wound healing growth factor TGFβ-1 | – Wound healing in humans does not occur via contraction but instead the formation of granulation tissue and re-epithelialization [78] – Approximately 80% of human DFUs occur in patients with type 2 diabetes |
| Microfiber mats of chitin/chitosan-glucan complex prepared by wet spinning [80] – FTIR-ATR spectrum of the complex showed the presence of β-configurations | – Enhanced and accelerated wound closure percentage of diabetic rats compared with diabetic non-treated ones
– Fully distinguishable and well symmetrically distributed collagen fibers observed under the new epidermal tissue | |
| Hyaluronic acid with silver nanoparticles [81] | – Formation and proliferation of hair follicles designating good dermal contraction and normal repair | |
| Nanofibrous glucophage-loaded collagen/PLGA scaffold membranes [82] – Fiber diameter: 203±41 nm – Porosity: 86.5±1.2% – Contact angle: 43.67±8.96° | – Electrospun glucophage-loaded collagen/PLGA nanofibers improved wound healing over that achieved using conventional gauze dressing (wound area 30% vs. 5%, respectively, after 14 days) – Denser soft tissue noted in the dermis layer – Higher collagen content | |
| Curcumin-loaded gum tragacanth/polycaprolactone electrospun fibers [83] | – Faster re-epithelialization – Increased collagen content in the dermis – Markedly fast wound closure with well-formed granulation tissue | |
| Neurotensin-loaded collagen dressings [84] | – Faster healing (17% wound area reduction) in the early phases of wound healing – Significantly reduced inflammatory cytokine expression, namely, TNF-α and IL-1β and decreased the inflammatory infiltrate at day 3 post-wounding – Reduced metalloproteinase 9 (MMP-9) in diabetic models, which significantly increased fibroblast migration and collagen (collagen type I, α 2 (COL1A2) and collagen type III, α 1 (COL3A1)) expression and deposition | |
| Chitosan-crosslinked collagen sponge [85] | – Shortest healing time; unhealed wound area of 15.8 vs. 180.1 mm2 for the control at day 14 – Quickest tissue collagen generation, the earliest and highest TGF-β1 expression, and dermal cell proliferation | |
| Astragulus polysaccharide-loaded PLGA fibrous mats [86] – Fiber diameter: 570–660 nm – Pore size: 79–103 μm – Porosity: 80–85% | – New blood vessel formation – Formation of dense collagen fibers and differentiated skin appendages | |
| Apigenin (APN)-loaded gellan gum-chitosan hydrogels [87] – Entrapment efficiency: 87.15±1.20% – 96.11% APN released in 24 h | – Significant increase in the wound contraction (95% vs. 71% on day 16) – Increase in antioxidant levels of the treated wounds compared to control group (23 vs. 10 μmol/50 mg tissue, respectively) | |
| Monocyte chemoattractant protein-1 (MCP-1)-loaded gelatin/PGA fibers [88] – Fiber diameter: 600–900 nm – Porosity: 69±10% | – Wound closure rates were 48.34%±10.23% in the group treated with the eluting scaffold vs. 80.27%±15.56% in the control group, on day 5 – A complete and thick epithelial layer was noted, whereas the control group showed an incomplete one – Largest population of F4/80+ murine macrophages in wound bed by day 3 | |
| Polyvinyl alcohol/chitosan nanofibrous mats [89] | – More blood vessels and higher cell numbers – Significant reduction observed in the length of epidermis gap and dermis area – Accelerated wound healing | |
| Nanobiocomposites consisting of cellulose nanocrystals and silver nanoparticles [90] | – Accelerated wound closure with no sign of infection – Growth of hair follicles – Early collagen formation and epithelialization | |
| Neomycin sulfate-loaded polyvinyl alcohol, polyvinyl pyrrolidone, and sodium alginate hydrogel [91] | – Improved wound curing – Disappearance of granulation tissue | |
| Curcumin-loaded composite chitosan and poly(propylene carbonate) fibers [92] | – Enhanced wound healing efficacy – Higher granulation scores and higher collagen contents |
3.2 Animal models for type 2 diabetes
Type 2 diabetes is modeled in both obese and non-obese animal models with varying degrees of insulin resistance and β-cell failure [35]. Most of the type 2 diabetes animal models are obese, reflecting the human condition where obesity is closely linked to type 2 diabetes development. Some of the most commonly used models of type 2 diabetes and their shortcomings for translation are outlined in Scheme 2. Problems such as generation of the model, correct human replication, and difference between male and female mice have been identified.

Translational barriers of type 2 diabetes mice models.
3.1 Spontaneous or genetically derived diabetic animals
3.1.1 Monogenic obese models
Monogenic obese diabetic animal models are commonly used in research despite the fact that obesity in humans is rarely caused by a monogenic mutation. The most widely used monogenic models of obesity are defective in leptin signaling. Leptin induces satiety, and thus, a lack of functional leptin results in hyperphagia and subsequent obesity. Examples of such models include the Lepob/ob mouse, which is deficient in leptin, and the Leprdb/db mouse and the Zucker diabetic fatty (ZDF) rat, which are deficient in the leptin receptor. However, the usage of these diabetic models are significantly limited [93], [94] as leptin plays an important role during wound healing, and therefore, the healing process of these gene-modified mice may be interfered by the mutation of leptin and/or the leptin receptor [95].
Lepob/ob mouse
The Lepob/ob mouse is a model of severe obesity. Their weight starts to increase at 2 weeks of age, and the animals develop hyperinsulinemia. Hyperglycemia becomes apparent by 4 weeks of age, with blood glucose concentrations continuing to rise, and peaking at 3–5 months, after which, they drop as the mouse becomes older [96]. These mice display other metabolic aberrations such as hyperlipidemia, a disturbance in temperature regulation, lower physical activity [96], and infertility [97]. Although there are some abnormalities in insulin release [98], the pancreatic islets maintain insulin secretion. The lack of complete β-cell failure in this model indicates that diabetes is not severe, and therefore, this model is not completely representative of human type 2 diabetes.
Leprdb/db mice
This model is due to an autosomal recessive mutation in the leptin receptor [99]. These animals are obese, hyperphagic, hyperinsulinemic, and hyperglycemic. Obesity is apparent from 3 to 4 weeks of age with hyperinsulinemia becoming evident at around 2 weeks of age and hyperglycemia developing at 4–8 weeks. These mice models develop ketosis after a few months of age and have a relative short lifespan [49].
Zucker fatty rats and Zucker diabetic fatty (ZDF) rats
These animals have a mutated leptin receptor [100] that causes hyperphagia. The rats become obese at around 4 weeks of age. These rats also show hyperinsulinemia, hyperlipidemia, hypertensivity, and have impaired glucose tolerance [49].
3.2.1.2 Polygenic obese models
Polygenic models of obesity may provide a more accurate model of the human condition. However, unlike the monogenic models, there are no wild-type controls. In addition, the male sex bias is more extreme in these models [101].
KK mice
KK mice are mildly obese, and they develop severe hyperinsulinemia, demonstrate insulin resistance in both muscle and adipose tissue, and show signs of diabetic nephropathy. The pancreatic islets in these animal models are hypertrophic, and the β cells are degranulated [102].
OLETF rats
OLETF rats display mild obesity and late-onset hyperglycemia (after 18 weeks), with males only suffering from diabetes. Three stages of histological changes are noted in the pancreatic islets. First, cellular infiltration and degeneration is seen when the animals are between 6 and 20 weeks old. This is followed by a stage of hyperplasia between 20 and 40 weeks. In the final stage, the islets become fibrotic and are replaced by connective tissue [103]. These rats also exhibit renal complications [104].
New Zealand obese (NZO) mice
These animals are hyperphagic and obese, which may be a consequence of leptin resistance [105]. About 50% of the males develop diabetes [106]. The islets are hyperplastic and hypertrophic at 3–6 months of age. However, β-cell loss occurs later, and there are signs of latent autoimmune diabetes in adults [107].
TallyHo/Jng mice
In these mice, adiposity is increased, and plasma triglycerides, cholesterol, and free fatty acid levels are elevated. Only male mice show hyperglycemia, which develops as early as between 10 and 14 weeks of age. The pancreatic islets are hypertrophied and degranulated, and hyperinsulinemia is evident. The TallyHo mouse has not yet been completely characterized for diabetic complications [101], although a recent study has used this model to study diabetic wound healing [108].
NoncNZO10/LtJ mice
These obese mice develop liver and skeletal muscle insulin resistance at 8 weeks and chronic hyperglycemia from about 12 weeks [101]. Diabetic nephropathy has been observed in some males aged about 1 year [101]. This model has been suggested to be suitable for studies in diabetic wound healing [109].
3.2.1.3 Non-obese models
Non-obese models include models that have β-cell inadequacy, which is what ultimately leads to type 2 diabetes in humans [110].
Goto-Kakizaki rats
This is a lean model of type 2 diabetes, which is characterized by glucose intolerance and defective glucose- induced insulin secretion. The β-cell mass and functions are abnormal [111], [112]. However, islet morphology and metabolism seem to differ between differing colonies of these rats.
3.2.2 Diet/Nutrition-induced diabetic animals
In these models, normal chow (on a caloric basis usually around 26% protein, 63% carbohydrate, and 11% fat) is exchanged for a diet where the number of calories from fat is substantially increased (around 58% of energy derived from fat). Indeed, high-fat feeding leads to obesity, hyperinsulinemia, and altered glucose homeostasis due to insufficient compensation by the islets [113]. The weight gain is associated with insulin resistance and the lack of β-cell compensation. Nutrition-induced diabetes is often used in transgenic or knock-out models, which may not show an overt diabetic phenotype under normal conditions.
3.2.3 Surgical diabetic models
This method consists of complete or partial pancreatectomy in animals. Partial pancreatectomy in animals performed as 70% or 90% dissection of the pancreas has been reported in various animal species mostly in dogs, pigs, rabbit, and also rats [114], [115]. It does not cause severe form of diabetes and is characterized by moderate hyperglycemia.
Compared to type I diabetes, very few in vivo studies with polymeric scaffolds have used animal models to investigate type 2 diabetes (Table 2). The db/db mouse seems to be able to mimic the human condition more closely. These studies most commonly looked at wound closure as the measurable outcome. Depending on the nature of scaffold and added molecule/growth factor, other parameters were then considered such as recruitment of macrophage or fibroblasts. In general, in vivo studies conducted using type 1 and type 2 diabetes models investigated the wound closure rates, re-epithelialization, organization of collagen fibers in the granulation tissue, and neovascularization. Investigations regarding the formation of skin appendages such as hair follicles were also made in type 1 diabetes models. However, the classification of wounds was also not addressed in type 2 models. In a study carried out on the implementation of a DFU classification system for research purposes to predict lower extremity amputation in type 2 diabetic patients, it was found that wound diameters and the surface area were not significantly associated with any type of amputation [127]. However, a significant association was found between the major lower extreme amputation and limb ischemia. Minor toe amputation was associated with deep wounds and those with grades 3 and 4 sepsis. Neuropathy was associated with both minor and major amputations. This study used the wound classification adopted by the International Consensus for the Diabetic. These criteria were the degree of limb ischemia, sensory neuropathy, depth and surface area of the wound, severity of sepsis, and ESRF (end-stage renal disease). Thus, it becomes important to evaluate polymeric scaffolds taking into consideration a system of wound classification.
Summary of main features of polymeric scaffolds and their resulting performances in other types of diabetic models (virally, genetically, or autoimmune-induced models).
| Animal model | Polymeric scaffold and its main features | In vivo performance compared to control | Translation in humans with type 2 diabetes | |
|---|---|---|---|---|
| Male Lewis rats and Zucker diabetic fatty diabetic rats [116] | Chitosan scaffolds loaded with plasmid DNA encoding perlecan domain I and VEGF189 Porous scaffolds with surface pore sizes ranging between 0.1 and 0.15 mm in diameter | – Re-epithelialization of the wounds with the formation of multiple layers of keratinocytes, while the control exhibited wound contraction as well as limited re-epithelialization and cell infiltration – Higher amount of fibrillar collagen deposited in the healing region of wounds – Significantly greater number of blood vessels in the wound site | – The db/db mice is a better model for the study of epithelial migration as considerably more of the repair occurs by epithelial migration compared to approximately 90% wound contraction seen in other mice models [117] – Compared to the models used, the db/db mouse has a reduced number of epidermal nerves, which is similar to humans with diabetes – db/db mouse was shown to have a comparable overexpression of neutral endopeptidase (NEP) to that seen in diabetic patients [118], [119] | |
| Male diabetic BKS.Cg-m+/+Lepr, db/db mice [120] | Fibrin-based scaffold incorporating VEGF- and bFGF-loaded PLGA nanoparticles – Sizes of unloaded NPs and the VEGF- and bFGF-loaded NPs were 116 ± 16 and 130 ± 18 nm (mean ± SD), respectively Encapsulation efficiencies of 75% and 95% were noted for VEGF and bFGF, respectively | – The closure rate of mice treated with the NP-loaded scaffold was similar to that observed in mice treated with scaffolds containing free VEGF and bFGF – Complete re-epithelialization, noted with enhanced granulation tissue formation/maturity and collagen deposition – At day 15, the mean size of the open wound area was significantly smaller in groups of mice treated with scaffold/GF-loaded NPs and scaffold/GFs (about 85% wound closure) compared to mice treated with control scaffold (20%) – A lower number of inflammatory cells and a higher number of fibroblast-like cells were visualized in the granulation tissue of wounds – Results obtained with the scaffold/GF-loaded NPs indicate enhanced delivery and increased concentrations of growth factors in the wound bed | – This model develops mild hyperglycemia and mild glucose intolerance [121] | |
| – Leprdb/db mice | Dextrin-rhEGF conjugate [122] | – At day 16, no significant differences in neo-dermal tissue formation were apparent between the dextrin conjugate groups and untreated controls – Significant reduction in wound area – Increased wound re-epithelialization – Significant increase in angiogenesis | – This strain heals wounds primarily by the formation of granulation tissue rather than by contraction [123] | |
| Composites consisting of silver nanoparticles and chitosan fibers [124] | – Delayed wound healing probably due to oxidative stress caused by nanoparticles – The wound repair could be restored by washing the covered wound with saline | |||
| Adult female genetically diabetic C57BL/KsJ (db/db) mice [125] | Oxidized regenerated cellulose (ORC)/collagen | – Significantly accelerated wound closure – Increase in cellularity with more mature wounds – Enhanced the formation and maturation of granulation tissue in the wounds of diabetic animals – Increased recruitment of macrophages and fibroblasts to the wound site | – Wound splinting has been shown to be an accurate and reproducible technique to minimize wound contraction, and allow wound healing to occur through the processes of granulation and re-epithelialization in mice. However, in this study, this method was not used and instead, full thickness excisional wounds were created through the panniculus carnosus and the hypodermis [126] |
4 Recommendations
Polymeric scaffolds offer the possibility to move toward personalized medicine. As highlighted in this review, very few in vivo studies have used animal models to investigate wound healing in type 2 diabetes models compared to type 1 diabetes. Furthermore, most studies conducted to improve wound healing following type 1 diabetes used animal models whereby diabetes was induced via the use of chemicals such as STZ or ALX. However, data obtained from such studies need careful interpretation due to toxicity caused to other organs by these chemicals. Hence, sufficient controls should be employed to ensure correct interpretation of results. Furthermore, the results obtained from such studies are highly variable depending on the animal strain, gender, etc. On the other hand, the studies investigating wound closure in type 2 diabetic models employed genetically derived animals. Moreover, these animals have a short lifespan, and most die from ketosis. In addition, while the vast majority of wounds are made on mouse trunk (dorsal) skin, healing progression varies depending on the species and body site. Models used, so far, are not completely representative of human type 2 diabetes given that diabetes development including pre-diabetic state, pathophysiology, and comorbidities are drastically different in these models compared with human disease, resulting in the lack of model translatability. A key feature of human diabetes is an intact leptin pathway; however, the most commonly used rat and mouse models feature monogenic mutations in either the leptin receptor (ZDF rat, db/db mouse) or the leptin molecule (ob/ob mouse). Diet-induced models with an intact leptin pathway rely on high-fat diets to induce diabetes, and result in models lacking key disease features such as hyperglycemia. Importantly, many current rodent models, including the ZDF rat, also lack the pre-diabetic state observed in human disease [126]. Indeed, the diabetic wound models described in this review are far from ideal and only represent the best available at present. Clearly, the best is yet to come. Until then, coupling together two or more existing impaired wound healing models in a combinatorial approach may offer an improved prediction of the human state. Ninety percent of type 2 diabetics are overweight or obese. Thus, coupling the different strains Lepob and Leprdb mice for type 2 diabetes, which can mimic different complications associated with diabetes and the use of new models such as TallyHo/JngJ mice polygenic model for obesity and diabetes, are new avenues to test the performance of polymeric scaffolds. The two novel polygenic mouse models of type 2 diabetes, TallyHo/JngJ and NONcNZO10/LtJ, contrast with C57BLKS/J-Leprdb (db/db) monogenic diabesity model as they are believed to be more representative of the obesity underlying human type 2 diabetes.
The ZDSD rat, recently developed by crossing the lean ZDF rat to an obesity-prone Sprague-Dawley (SD) rat with polygenic obesity, is believed to be the most translatable diabetes rodent model (Figure 2). This model has an intact leptin pathway and progressively develops many human-like features of diabetes and metabolic syndrome while on a normal chow diet. Insulin resistance and β-cell failure are observed in the ZDSD rat, in a manner similar to the human disease. Furthermore, insulin resistance persists across the pre-diabetic and diabetic states in both ZDSD rats and humans, in contrast to currently available rodent models, e.g. the ZDF rat, where β-cell failure is observed at a much earlier time frame [128].

Increased translatability of ZDSD rat.
Reverse translation identifying the causes of treatment failure is crucial in diabetic wound healing and would allow the engineering of more adapted scaffolds and use of proper multi pre-clinical animal models as well as shed light on the molecular mechanism of wound healing. These factors will open up the possibility to categorize available skin substitutes as well as new nanoscaffold treatments according to the patients’ conditions and, thus, enhance success rate.
5 Conclusions
The improvement of diabetic wound healing still represents a major clinical challenge to researchers. The challenge faced is to identify a suitable animal model that best represents the human situation. However, the majority of diabetic wound healing models are in rodents and are hindered by rapid contraction and, thus, do not reflect epithelial cell migration, as seen in the human wound [129]. Confusingly, the observed wound repair phenotype differs between the diabetic models described and the wound type employed [130], [131], complicating the prediction of the human clinical state. Hence, ideally, more than one animal model should be used to represent the diversity seen in human diabetic patients. A synergy is required between clinicians, biologists, and biomedical engineer/polymer chemists for the success of polymeric scaffolds.
About the authors

Nowsheen Goonoo holds a PhD in Polymers and Biomaterials and is currently a Postdoctoral Researcher at the Biomaterials, Drug Delivery and Nanotechnology Unit, Center for Biomedical and Biomaterials Research. She was awarded the Georg Forster Postdoctoral Fellow, Alexander Von Humboldt Foundation (AvH) in 2015, and spent 1 year at the University of Siegen, Germany. She was granted a Return Fellowship for Research Fellows (AvH) in August 2017. Her research interests include cell-scaffold interactions and polymeric biomaterials for biomedical applications.

Archana Bhaw-Luximon holds a PhD in Polymer Chemistry and is currently an Associate Professor and Unit Leader of the Biomaterials, Drug Delivery and Nanotechnology Unit, Center for Biomedical and Biomaterials Research. The research thrusts of her Unit are design of polymeric biomaterials for fabrication of scaffolds for skin and bone tissue regeneration, engineering nanocarriers for anti-cancer and anti-diabetic drugs/biomolecules, and exploitation of materials from land and marine resources such as sugarcane bagasse and seaweeds for high-end applications.
Acknowledgments
The authors thank the Mauritius Research Council for funding biomaterials and drug delivery research at the Biomaterials, Drug Delivery and Nanotechnology Unit, Centre for Biomedical and Biomaterials Research. All authors have read the journal’s authorship agreement.
References
[1] Nunan R, Harding KG, Martin P. Clinical challenges of chronic wounds: searching for an optimal animal model to recapitulate their complexity. Dis. Model Mech. 2014, 7, 1205–1213.10.1242/dmm.016782Search in Google Scholar PubMed PubMed Central
[2] Mei L, Wang YL, Tong AP, Guo G. Facile electrospinning of an efficient drug delivery. Expert Opin. Drug Deliv. 2016, 25, 1–13.10.1517/17425247.2016.1142525Search in Google Scholar PubMed
[3] Haddad AG, Giatsidis G, Orgill DP, Halvorson EG. Skin substitutes and bioscaffolds: temporary and permanent coverage. Clin. Plast. Surg. 2017, 44, 627–634.10.1016/j.cps.2017.02.019Search in Google Scholar PubMed
[4] Korrapati PS, Karthikeyan K, Satish A, Krishnaswamy VR, Venugopal JR, Ramakrishna S. Recent advancements in nanotechnological strategies in selection, design and delivery of biomolecules for skin regeneration. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 67, 747–765.10.1016/j.msec.2016.05.074Search in Google Scholar PubMed
[5] Pereira RF, Barrias CC, Granja PL, Bartolo PJ. Advanced biofabrication strategies for skin regeneration and repair. Nanomedicine (London UK) 2013, 8, 603–621.10.2217/nnm.13.50Search in Google Scholar PubMed
[6] Reddy VJ, Radhakrishnan S, Ravichandran R, Mukherjee S, Balamurugan R, Sundarrajan S, Ramakrishna S. Nanofibrous structured biomimetic strategies for skin tissue regeneration. Wound Repair Regen. 2013, 21, 1–16.10.1111/j.1524-475X.2012.00861.xSearch in Google Scholar PubMed
[7] Pham C, Greenwood J, Cleland H, Woodruff P, Maddern G. Bioengineered skin substitutes for the management of burns: a systematic review. Burns 2007, 33, 946–957.10.1016/j.burns.2007.03.020Search in Google Scholar PubMed
[8] Zhong SP, Zhang YZ, Lim CT. Tissue scaffolds for skin wound healing and dermal reconstruction. WIREs Interdiscip. Rev. Nanomed. Nanobiotechnol. 2010, 2, 510–525.10.1002/wnan.100Search in Google Scholar PubMed
[9] Dias JR, Granja PL, Bartolo PJ. Advances in electrospun skin substitutes. Prog. Mater. Sci. 2016, 84, 314–334.10.1016/j.pmatsci.2016.09.006Search in Google Scholar
[10] Groeber F, Holeiter M, Hampel M, Hinderer S, Schenke-Layland K. Skin tissue engineering – in vivo and in vitro applications. Adv. Drug Deliv. Rev. 2011, 63, 352–366.10.1016/j.addr.2011.01.005Search in Google Scholar PubMed
[11] Van der Veen VC, Van der Wal, MBA, Van Leeuwen MCE, Ulrich MMW, Middelkoop E. Biological background of dermal substitutes. Burns 2010, 36, 305–321.10.1016/j.burns.2009.07.012Search in Google Scholar PubMed
[12] Poinern GE, Fawcett D, Ng YJ, Ali N, Brundavanam RK, Jiang ZT. Nanoengineering a biocompatible inorganic scaffold for skin wound healing. J. Biomed. Nanotechnol. 2010, 6, 497–510.10.1166/jbn.2010.1148Search in Google Scholar PubMed
[13] Kearney JN. Clinical evaluation of skin substitutes. Burns 2001, 27, 545–551.10.1016/S0305-4179(01)00020-1Search in Google Scholar PubMed
[14] McNeil S. Biomaterials for tissue engineering of skin. Mater. Today 2008, 11, 26–36.10.1016/S1369-7021(08)70087-7Search in Google Scholar
[15] Casey G. The physiology of the skin. Nurs. Stand. 2002, 16, 47–51.10.7748/ns2002.05.16.34.47.c3193Search in Google Scholar PubMed
[16] Tortora GJ, Nielsen M. Principles of Anatomy and Physiology, 13th ed., Wiley: Hoboken, NJ, USA, 2013.Search in Google Scholar
[17] Breitkreutz D, Mirancea N, Nischt R. Basement membranes in skin: unique matrix structures with diverse functions? Histochem. Cell Biol. 2009, 132, 1–10.10.1007/s00418-009-0586-0Search in Google Scholar PubMed
[18] Enoch S, Leaper DJ. Basic science of wound healing. Surgery (Oxford) 2005, 23, 37–42.10.1383/surg.23.2.37.60352Search in Google Scholar
[19] Polverini PJ. Angiogenesis and wound healing: basic discoveries, clinical implications, and therapeutic opportunities. Endod. Topics 2011, 24, 130–145.10.1111/etp.12005Search in Google Scholar
[20] Hinz, B. Formation and function of the myofibroblast during tissue repair. J. Investig. Dermatol. 2007, 127, 526–537.10.1038/sj.jid.5700613Search in Google Scholar PubMed
[21] Okamoto O, Fujiwara S. Dermatopontin, a novel player in the biology of the extracellular matrix. Connect. Tissue Res. 2006, 47, 177–189.10.1080/03008200600846564Search in Google Scholar PubMed
[22] Young A, McNaught CE. The physiology of wound healing. Surgery (Oxford) 2011, 29, 475–479.10.1016/j.mpsur.2011.06.011Search in Google Scholar
[23] MacKay D, Miller AL. Nutritional support for wound healing. Altern. Med. Rev. 2003, 8, 359–377.Search in Google Scholar PubMed
[24] Böttcher-Haberzeth S, Biedermann T, Reichmann E. Burns 2010, 36, 450–460.10.1016/j.burns.2009.08.016Search in Google Scholar PubMed
[25] Shores JT, Gabriel A, Gupta S. Skin substitutes and alternatives: a review. Adv. Skin Wound Care 2007, 20, 493–508.10.1097/01.ASW.0000288217.83128.f3Search in Google Scholar PubMed
[26] Sheridan RL, Moreno C. Skin substitutes in burns. Burns 2001, 27, 92.10.1016/S0305-4179(00)00076-0Search in Google Scholar PubMed
[27] Alrubaiy L, Al-Rubaiy KK. Skin substitutes: a brief review of types and clinical applications. Oman Med. J. 2009, 24, 4–6.10.5001/omj.2009.2Search in Google Scholar PubMed PubMed Central
[28] Halim AS, Khoo TL, Yussof Shah JM. Biologic and synthetic skin substitutes: an overview. Indian J. Plast. Surg. 2010, 43(Suppl S1), 23–28.10.4103/0970-0358.70712Search in Google Scholar PubMed PubMed Central
[29] Debels H, Hamdi M, Abberton K, Morrison W. Dermal matrices and bioengineered skin substitutes: a critical review of current options. Plast. Reconstr. Surg. Glob. Open 2015, 3, e284.10.1097/GOX.0000000000000219Search in Google Scholar PubMed PubMed Central
[30] Hamdan S, Pastar I, Drakulich S, Dikici E, Tomic-Canic M, Deo S, Daunert S. Nanotechnology-driven therapeutic interventions in wound healing: potential uses and applications. ACS Cent. Sci. 2017, 3, 163–175.10.1021/acscentsci.6b00371Search in Google Scholar PubMed PubMed Central
[31] Tocco I, Zavan B, Bassetto F, Vindigni V. Nanotechnology-based therapies for skin wound regeneration. J. Nanomater. 2012, 2012, Article ID 714134, 11 pages.10.1155/2012/714134Search in Google Scholar
[32] Colonna MR, Flavia L, Gabriele D, d’Alcontres Fransesco S. In Regenerative Approaches in Wound Healing: New Alternatives for Older Tools, Wound Healing – New insights into Ancient Challenges, Alexandrescu, V, Ed., InTech, 2016. DOI: 10.5772/64133. Available from: https://www.intechopen.com/books/wound-healing-new-insights-into-ancient-challenges/regenerative-approaches-in-wound-healing-new-alternatives-for-older-tools.10.5772/64133Search in Google Scholar
[33] Cortivo R, Vindigni V, Iacobellis L, Abatangelo G, Pinton P. Nanoscale particle therapies for wounds and ulcers. Nanomedicine (Lond) 2010, 5, 641–656.10.2217/nnm.10.32Search in Google Scholar PubMed
[34] Parani M, Lokhande G, Singh A, Gaharwar AK. Engineered nanomaterials for infection control and healing acute and chronic wounds. ACS Appl. Mater. Interfaces 2016, 8, 10049–10069.10.1021/acsami.6b00291Search in Google Scholar PubMed
[35] King AJ. The use of animal models in diabetes research. Br. J. Pharmacol. 2012, 166, 877–894.10.1111/j.1476-5381.2012.01911.xSearch in Google Scholar
[36] Global Report on Diabetes. World Health Organization, Geneva, 2016; Avaialbe online at http://apps.who.int/iris/bitstream/10665/204871/1/9789241565257_eng.pdf.Search in Google Scholar
[37] Lioupis C. Effects of diabetes mellitus on wound healing: an update. J. Wound Care 2005, 14, 84–86.10.12968/jowc.2005.14.2.26738Search in Google Scholar PubMed
[38] Yazdanpanah L, Nasiri M, Adarvishi S. Literature review on the management of diabetic foot ulcer. World J. Diabetes 2015, 6, 37–53.10.4239/wjd.v6.i1.37Search in Google Scholar PubMed
[39] Mathers CD, Loncar D. Projections of global mortality and burden of disease from 2002 to 2030. PLoS Med. 2006, 3, e442.10.1371/journal.pmed.0030442Search in Google Scholar PubMed
[40] Claxton MJ, Armstrong DG, Boulton AJM. Healing the diabetic wound and keeping it healed: modalities for the early 21st century. Curr. Diab. Rep. 2002, 2, 510–518.10.1007/s11892-002-0121-3Search in Google Scholar PubMed
[41] Falanga V. Wound healing and its impairment in the diabetic foot. Lancet 2005, 366, 1736–1743.10.1016/S0140-6736(05)67700-8Search in Google Scholar PubMed
[42] Gallagher KA, Liu ZJ, Xiao M, Chen H, Goldstein LJ, Buerk DG, Nedeau A, Thom SR, Velazquez OC. Diabetic impairments in NO-mediated endothelial progenitor cell mobilization and homing are reversed by hyperoxia and SDF-1α. J. Clin. Invest. 2007, 117, 1249–1259.10.1172/JCI29710Search in Google Scholar PubMed PubMed Central
[43] Lee JH, Yang SH, Oh JM, Lee MG. Pharmacokinetics of drugs in rats with diabetes mellitus induced by alloxan or streptozocin: comparison with those in patients with type I diabetes mellitus. J. Pharm. Pharmacol. 2010, 62, 1–23.10.1211/jpp.62.01.0001Search in Google Scholar PubMed
[44] AKC Jain. A new classification of diabetic foot complications: a simple and effective teaching tool. The Journal of Diabetic Foot Complications 2012, 4, 1–5.Search in Google Scholar
[45] Mathews CE. Utility of murine models for the study of spontaneous autoimmune type 1 diabetes. Pediatr. Diabetes 2005, 6, 165–177.10.1111/j.1399-543X.2005.00123.xSearch in Google Scholar PubMed
[46] Brito-Castillas Y, Melian C, Wägner AM. Study of the pathogenesis and treatment of diabetes mellitus through animal models. Endocrinol. Nutr. 2016, 63, 345–353.10.1016/j.endonu.2016.03.011Search in Google Scholar PubMed
[47] Szkudelski T. The mechanism of alloxan and streptozotocin action in B cells of the rat pancreas. Physiol. Res. 2001, 50, 537–546.Search in Google Scholar
[48] Sandler S, Swenne I. Streptozotocin, but not alloxan, induces DNA repair synthesis in mouse pancreatic islets in vitro. Diabetologia 1983, 25, 444–447.10.1007/BF00282526Search in Google Scholar PubMed
[49] Nerup J, Mandrup-Poulsen T, Helqvist S, Andersen HU, Pociot F, Reimers JI, Cuartero BG, Karlsen AE, Bjerre U, Lorenzen T. On the pathogenesis of IDDM. Diabetologia 1994, 37(Suppl. 2), S82–S89.10.1007/BF00400830Search in Google Scholar PubMed
[50] Dekel Y, Glucksam Y, Elron-Gross I, Margalit R. Insights into modeling streptozotocin-induced diabetes in ICR mice. Lab Anim. (NY) 2009, 38, 55–60.10.1038/laban0209-55Search in Google Scholar PubMed
[51] Hayashi K, Kojima R, Ito M. Strain differences in the diabetogenic activity of streptozotocin in mice. Biol. Pharm. Bull. 2006, 29, 1110–1119.10.1248/bpb.29.1110Search in Google Scholar PubMed
[52] Srinivasan K, Ramarao P. Animal models in type 2 diabetes research: an overview. Indian J. Med. Res. 2007, 125, 451–472.Search in Google Scholar PubMed
[53] Grossman EJ, Lee DD, Tao J, Wilson RA, Park SY, Bell GI, Chong AS. Glycemic control promotes pancreatic beta-cell regeneration in streptozotocin-induced diabetic mice. PLoS One 2010, 5, e8749.10.1371/journal.pone.0008749Search in Google Scholar PubMed PubMed Central
[54] Muller YD, Golshayan D, Ehirchiou D, Wyss JC, Giovannoni L, Meier R, Serre-Beinier V, Yung GP, Morel P, Bühler LH, Seebach JD. Immunosuppressive effects of streptozotocin-induced diabetes result in absolute lymphopenia and a relative increase of T regulatory cells. Diabetes 2011, 60, 2331–2340.10.2337/db11-0159Search in Google Scholar PubMed PubMed Central
[55] Wang Z, Gleichmann H. GLUT2 in pancreatic islets: crucial target molecule in diabetes induced with multiple low doses of streptozotocin in mice. Diabetes 1998, 47, 50–56.10.2337/diab.47.1.50Search in Google Scholar PubMed
[56] Lukić ML, Stosić-Grujicić S, Shahin A. Effector mechanisms in low-dose streptozotocin-induced diabetes. Dev. Immunol. 1998, 6, 119–128.10.1155/1998/92198Search in Google Scholar PubMed PubMed Central
[57] Reddy S, Wu D, Elliott RB. Low dose streptozotocin causes diabetes in severe combined immunodeficient (Scid) mice without immune cell infiltration of the pancreatic islets. Autoimmunity 1994, 20, 83–92.10.3109/08916939509001931Search in Google Scholar PubMed
[58] Deeds MC, Anderson JM, Armstrong AS, Gastineau DA, Hiddinga HJ, Jahangir A, Eberhadt NL, Kudva YC. Single dose streptozotocin induced diabetes: considerations for study design in islet transplantation models. Lab Anim. 2011, 45, 131–140.10.1258/la.2010.010090Search in Google Scholar PubMed PubMed Central
[59] Kang Y, Chatterjee NK, Nodwell MJ, Yoon JW. Complete nucleotide sequence of a strain of coxsackie B4 virus of human origin that induces diabetes in mice and its comparison with nondiabetogenic coxsackie B4 JBV strain. J. Med. Virol. 1994, 44, 353–361.10.1002/jmv.1890440408Search in Google Scholar PubMed
[60] Jaidane H, Sane F, Gharbi J, Aouni M, Romond MB, Hober D. Coxsackievirus B4 and type 1 diabetes pathogenesis: contribution of animal models. Diabetes Metab. Res. Rev. 2009, 25, 591–603.10.1002/dmrr.995Search in Google Scholar PubMed
[61] Craighead JE, McLane MF. Diabetes mellitus: induction in mice by encephalomyocarditis virus. Science 1968, 162, 913–914.10.1126/science.162.3856.913Search in Google Scholar PubMed
[62] Shimada A, Maruyama T. Encephalomyocarditis-virus induced diabetes model resembles ‘fulminant’ type 1 diabetes in humans. Diabetologia 2004, 47, 1854–1855.10.1007/s00125-004-1538-9Search in Google Scholar PubMed
[63] Guberski DL, Thomas VA, Shek WR, Like AA, Handler ES, Rossini AA, Wallace JE, Welsh RM. Induction of type I diabetes by Kilham’s rat virus in diabetes-resistant BB/Wor rats. Science 1991, 254, 1010–1013.10.1126/science.1658938Search in Google Scholar PubMed
[64] Von Herrath MG, Filippi C, Coppieters K. How viral infections enhance or prevent type 1 diabetes-from mouse to man. J. Med. Virol. 2011, 83, 1672.10.1002/jmv.22063Search in Google Scholar PubMed
[65] Jun HS, Yoon JW. A new look at viruses in type 1 diabetes. Diabetes Metab. Res. Rev. 2003, 19, 8–31.10.1002/dmrr.337Search in Google Scholar PubMed
[66] Kim J, Nishina HP, Naggert JK. Genetic models for non-insulin dependent diabetes mellitus in rodents. J. Basic Clin. Physiol. Pharmacol. 1998, 9, 325–345.10.1515/JBCPP.1998.9.2-4.325Search in Google Scholar PubMed
[67] Yang Y, Santamaria P. Lessons on autoimmune diabetes from animal models. Clin. Sci. (Lond) 2006, 110, 627–639.10.1042/CS20050330Search in Google Scholar PubMed
[68] Yoon JW, Jun HS. Cellular and molecular pathogenic mechanisms of insulin-dependent diabetes mellitus. Ann. N. Y. Acad. Sci. 2001, 928, 200–211.10.1111/j.1749-6632.2001.tb05650.xSearch in Google Scholar PubMed
[69] Pozzilli P, Signore A, Williams AJ, Beales PE. NOD mouse colonies around the world – recent facts and figures. Immunol. Today 1993, 14, 193–196.10.1016/0167-5699(93)90160-MSearch in Google Scholar PubMed
[70] Driver JP, Serreze DV, Chen YG. Mouse models for the study of autoimmune type 1 diabetes: a NOD to similarities and differences to human disease. Semin. Immunopathol. 2011, 33, 67–87.10.1007/s00281-010-0204-1Search in Google Scholar PubMed
[71] von Herrath M, Nepom GT. Animal models of human type 1 diabetes. Nat. Immunol. 2009, 10, 129–132.10.1038/ni0209-129Search in Google Scholar PubMed
[72] von Herrath MG, Nepom GT. Lost in translation: barriers to implementing clinical immunotherapeutics for autoimmunity. J. Exp. Med. 2005, 202, 1159–1162.10.1084/jem.20051224Search in Google Scholar PubMed PubMed Central
[73] Mordes JP, Bortell R, Blankenhorn EP, Rossini AA, Greiner DL. Rat models of type 1 diabetes: genetics, environment, and autoimmunity. ILAR J. 2004, 45, 278–291.10.1093/ilar.45.3.278Search in Google Scholar PubMed
[74] Senturk B, Mercan S, Delibasi T, Guler MO, Tekinay AB. Angiogenic peptide nanofibers improve wound healing in STZ-induced diabetic rats. ACS Biomater. Sci. Eng. 2016, 2, 1180–1189.10.1021/acsbiomaterials.6b00238Search in Google Scholar PubMed
[75] Karri VV, Kuppusamy G, Talluri SV, Mannemala SS, Kollipara R, Wadhwani AD, Mulukutla S, Raju KR, Malayandi R. Curcumin loaded chitosan nanoparticles impregnated into collagen-alginate scaffolds for diabetic wound healing. Int. J. Biol. Macromol. 2016, 93(Pt B), 1519–1529.10.1016/j.ijbiomac.2016.05.038Search in Google Scholar PubMed
[76] Olerud JE. Models for diabetic wound healing and healing into percutaneous devices. J. Biomater. Sci. Polym. Ed. 2008, 19, 1007–1020.10.1163/156856208784909426Search in Google Scholar PubMed
[77] Lavery LA, Barnes SA, Keith MS, Seaman JW, Armstrong DG. Prediction of healing for postoperative diabetic foot wounds based on early wound area progression. Diabetes Care 2008, 31, 26–29.10.2337/dc07-1300Search in Google Scholar PubMed
[78] Wong VW, Sorkin M, Glotzbach JP, Longaker MT, Gurtner GC. Surgical approaches to create murine models of human wound healing. J. Biomed. Biotechnol. 2011, 2011, Article ID 969618. 8 pages10.1155/2011/969618Search in Google Scholar PubMed PubMed Central
[79] Kaisang L, Siyu W, Lijun F, Daoyan P, Xian CJ, Jie S. Adipose-derived stem cells seeded in Pluronic F127 hydrogel promotes diabetic wound healing. J. Surg. Res. 2017, 217, 63–74.10.1016/j.jss.2017.04.032Search in Google Scholar PubMed
[80] Abdel-Mohsen AM, Jancar J, Massoud D, Fohlerova Z, Elhadidy H, Spotz Z, Heheish A. Novel chitin/chitosan-glucan wound dressing: isolation, characterization, antibacterial activity and wound healing properties. Int. J. Pharm. 2016, 510, 86–99.10.1016/j.ijpharm.2016.06.003Search in Google Scholar PubMed
[81] Fouda MM, Abdel-Mohsen AM, Ebaid H, Hassan I, Al-Tamimi J, Abdel-Rahman RM, Metwalli A, Alhazza I, Rady A, El-Faham A, Jancar J. Wound healing of different molecular weight of hyaluronan; in-vivo study. Int. J. Biol. Macromol. 2016, 89, 582–591.10.1016/j.ijbiomac.2016.05.021Search in Google Scholar PubMed
[82] Lee CH, Chang SH, Chen WJ, Hung KC, Lin YH, Liu SJ, Hsieh MJ, Pang JH, Juang JH. Augmentation of diabetic wound healing and enhancement of collagen content using nanofibrous glucophage-loaded collagen/PLGA scaffold membranes. J. Colloid Interface Sci. 2015, 439, 88–97.10.1016/j.jcis.2014.10.028Search in Google Scholar PubMed
[83] Ranjbar-Mohammadi M, Rabbani S, Bahrami SH, Joghataei MT, Moayer F. Antibacterial performance and in vivo diabetic wound healing of curcumin loaded gum tragacanth/poly(ε-caprolactone) electrospun nanofibers. Mater. Sci. Eng. C Mater. Biol. Appl. 2016, 69, 1183–1191.10.1016/j.msec.2016.08.032Search in Google Scholar PubMed
[84] Moura LIF, Dias AMA, Suesca E, Casadiegos S, Leal EC, Fontanilla MR, Carvalho L, de Sousa HC, Carvalho E. Neurotensin-loaded collagen dressings reduce inflammation and improve wound healing in diabetic mice. Biochim. Biophys. Acta 2014, 1842, 32–43.10.1016/j.bbadis.2013.10.009Search in Google Scholar PubMed
[85] Wang W, Lin S, Xiao Y, Huang Y, Tan Y, Cai L, Li X. Acceleration of diabetic wound healing with chitosan-crosslinked collagen sponge containing recombinant human acidic fibroblast growth factor in healing-impaired STZ diabetic rats. Life Sci. 2008, 82, 190–204.10.1016/j.lfs.2007.11.009Search in Google Scholar PubMed
[86] Yang Y, Wang F, Yin D, Fang Z, Huang L. Astragulus polysaccharide-loaded fibrous mats promote the restoration of microcirculation in/around skin wounds to accelerate wound healing in a diabetic rat model. Colloids Surf. B Biointerfaces 2015, 136, 111–118.10.1016/j.colsurfb.2015.09.006Search in Google Scholar PubMed
[87] Shukla R, Kashaw SK, Jain AP, Lodhi S. Fabrication of Apigenin loaded gellan gum–chitosan hydrogels (GGCH-HGs) for effective diabetic wound healing. Int. J. Biol. Macromol. 2016, 91, 1110–1119.10.1016/j.ijbiomac.2016.06.075Search in Google Scholar PubMed
[88] Yin H, Ding G, Shi X, Guo W, Ni Z, Fu H, Fu Z. A bioengineered drug-eluting scaffold accelerated cutaneous wound healing in diabetic mice. Colloids Surf. B Biointerfaces 2016, 145, 226–231.10.1016/j.colsurfb.2016.04.056Search in Google Scholar PubMed
[89] Ahmadi MS, Rabbani KM, Moshtaghian SJ, Talebi A, Khezri M. Application of Chitosan/PVA Nano fiber as a potential wound dressing for streptozotocin-induced diabetic rats. Int. J. Biol. Macromol. 2016, 92, 1162–1168.10.1016/j.ijbiomac.2016.06.035Search in Google Scholar PubMed
[90] Singla R, Soni S, Patial V, Kulurkar PM, Kumari ASM, Padwad YS, Yadav SK. In vivo diabetic wound healing potential of nanobiocomposites containing bamboo cellulose nanocrystals impregnated with silver nanoparticles. Int. J. Biol. Macromol. 2017, DOI: 10.1016/j.ijbiomac.2017.06.10.10.1016/j.ijbiomac.2017.06.10Search in Google Scholar PubMed
[91] Choi JS, Kim DW, Kim DS, Kim JO, Yong CS, Cho KH, Youn YS, Jin SG, Choi HG. Novel neomycin sulfate-loaded hydrogel dressing with enhanced physical dressing properties and wound-curing effect. Drug Deliv. 2016, 23, 2806–2812.10.3109/10717544.2015.1089958Search in Google Scholar PubMed
[92] Mei L, Fan RR, Li XL, Wang YL, Han B, Gu YC, Zhou LX, Zheng Y, Tong AP, Guo G. Nanofibers for improving the wound repair process: the combination of a grafted chitosan and an antioxidant agent. Polym Chem-UK. 2017, 8, 1664–1671.10.1039/C7PY00038CSearch in Google Scholar
[93] Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994, 372, 425–32.10.1038/372425a0Search in Google Scholar PubMed
[94] Sharma K, McCue P, Dunn SR. Diabetic kidney disease in the db/db mouse. Am. J. Physiol. Renal Physiol. 2003, 284, F1138–F1144.10.1152/ajprenal.00315.2002Search in Google Scholar PubMed
[95] Murad A, Nath AK, Cha ST, Demir E, Flores-Riveros J, Sierra-Honigmann MR. Leptin is an autocrine/paracrine regulator of wound healing. FASEB J. 2003, 17, 1895–1897.10.1096/fj.03-0068fjeSearch in Google Scholar PubMed
[96] Lindstrom P. The physiology of obese-hyperglycemic mice [ob/ob mice]. Sci. World J. 2007, 7, 666–685.10.1100/tsw.2007.117Search in Google Scholar PubMed PubMed Central
[97] Chehab FF, Lim ME, Lu R. Correction of the sterility defect in homozygous obese female mice by treatment with the human recombinant leptin. Nat. Genet. 1996, 12, 318–320.10.1038/ng0396-318Search in Google Scholar PubMed
[98] Lavine RL, Voyles N, Perrino PV, Recant L. Functional abnormalities of islets of Langerhans of obese hyperglycemic mouse. Am. J. Physiol. 1977, 233, E86–E90.10.1152/ajpendo.1977.233.2.E86Search in Google Scholar
[99] Chen H, Charlat O, Tartaglia LA, Woolf EA, Weng X, Ellis SJ, Lakey ND, Culpepper J, Moore KJ, Breitbart RE, Duyk GM, Tepper RI, Morgenstern JP. Evidence that the diabetes gene encodes the leptin receptor: identification of a mutation in the leptin receptor gene in db/db mice. Cell 1996, 84, 491–495.10.1016/S0092-8674(00)81294-5Search in Google Scholar PubMed
[100] Phillips MS, Liu Q, Hammond HA, Dugan V, Hey PJ, Caskey CJ, Hess JF. Leptin receptor missense mutation in the fatty Zucker rat. Nat. Genet. 1996, 13, 18–19.10.1038/ng0596-18Search in Google Scholar PubMed
[101] Leiter EH. Selecting the ‘right’ mouse model for metabolic syndrome and type 2 diabetes research. Methods Mol. Biol. 2009, 560, 1–17.10.1007/978-1-59745-448-3_1Search in Google Scholar PubMed
[102] Ikeda H. KK mouse. Diabetes Res. Clin. Pract. 1994, 24(Suppl.), S313–S316.10.1016/0168-8227(94)90268-2Search in Google Scholar
[103] Kawano K, Hirashima T, Mori S, Natori T. OLETF (Otsuka Long-Evans Tokushima Fatty) rat: a new NIDDM rat strain. Diabetes Res. Clin. Pract. 1994, 24(Suppl.), S317–S320.10.1016/0168-8227(94)90269-0Search in Google Scholar
[104] Lee MY, Shim MS, Kim BH, Hong SW, Choi R, Lee EY, Nam SM, Kim GW, Shin JY, Shin YG, Chung CH. Effects of spironolactone and losartan on diabetic nephropathy in a type 2 diabetic rat model. Diabetes Metab. J. 2011, 35, 130–137.10.4093/dmj.2011.35.2.130Search in Google Scholar PubMed PubMed Central
[105] Leiter EH, Reifsnyder PC. Differential levels of diabetogenic stress in two new mouse models of obesity and type 2 diabetes. Diabetes 2004, 53(Suppl. 1), S4–S11.10.2337/diabetes.53.2007.S4Search in Google Scholar
[106] Haskell BD, Flurkey K, Duffy TM, Sargent EE, Leiter EH. The diabetes-prone NZO/HlLt strain. I. Immunophenotypic comparison to the related NZB/BlNJ and NZW/LacJ strains. Lab Invest. 2002, 82, 833–842.10.1097/01.LAB.0000018915.53257.00Search in Google Scholar PubMed
[107] Junger E, Herberg L, Jeruschke K, Leiter EH. The diabetes-prone NZO/Hl strain. II. Pancreatic immunopathology. Lab Invest. 2002, 82, 843–853.10.1097/01.LAB.0000018917.69993.BASearch in Google Scholar PubMed
[108] Buck DW, Jin DP, Geringer M, Jong HS, Galiano RD, Mustoe TA. The TallyHo polygenic mouse model of diabetes: implications in wound healing. Plast. Reconstr. Surg. 2011, 128, 427e–437e.10.1097/PRS.0b013e31822b7333Search in Google Scholar
[109] Fang RC, Kryger ZB, Buck DW, De la Garza M, Galiano RD, Mustoe TA. Limitations of the db/db mouse in translational wound healing research: is the NONcNZO10 polygenic mouse model superior? Wound Repair Regen. 2010, 18, 605–613.10.1111/j.1524-475X.2010.00634.xSearch in Google Scholar PubMed
[110] Weir GC, Marselli L, Marchetti P, Katsuta H, Jung MH, Bonner-Weir S. Towards better understanding of the contributions of overwork and glucotoxicity to the beta-cell inadequacy of type 2 diabetes. Diabetes Obes. Metab. 2009, 11(Suppl. 4), 82–90.10.1111/j.1463-1326.2009.01113.xSearch in Google Scholar PubMed
[111] Portha B, Giroix MH, Serradas P, Gangnerau MN, Movassat J, Rajas F, Bailbe D, Plachot C, Mithieux G, Marie JC. Beta-cell function and viability in the spontaneously diabetic GK rat: information from the GK/Par colony. Diabetes 2001, 50(Suppl. 1), S89–S93.10.2337/diabetes.50.2007.S89Search in Google Scholar
[112] Ostenson CG, Efendic S. Islet gene expression and function in type 2 diabetes; studies in the Goto-Kakizaki rat and humans. Diabetes Obes. Metab. 2007, 9(Suppl. 2), 180–186.10.1111/j.1463-1326.2007.00787.xSearch in Google Scholar PubMed
[113] Winzell MS, Ahren B. The high-fat diet-fed mouse: a model for studying mechanisms and treatment of impaired glucose tolerance and type 2 diabetes. Diabetes 2004, 53(Suppl. 3), S215–S219.10.2337/diabetes.53.suppl_3.S215Search in Google Scholar
[114] Ibanez-Camacho R, Meckes-Lozaya M, Mellado-Campos V. The hypoglucemic effect of Opuntia streptocanthe studied in different animal experimental models. J. Ethnopharmacol. 1983, 7, 175–181.10.1016/0378-8741(83)90019-3Search in Google Scholar PubMed
[115] Sasaki S, Nio Y, Hirahara N, Sato Y, Inoue Y, Iguchi C, Yamasawa K, Tamura K. Intraperitoneally implanted artificial pancrease with transkaryotic beta-cells on microcarrier beads in a diffusion chamber improves hyperglycemia after 90% pancreatectomy in rats. In Vivo 2000, 14, 535–541.Search in Google Scholar
[116] Lord MS, Ellis AL, Farrugia BL, Whitelock JM, Grenett H, Li C, O’Grady RL, DeCarlo AA. Perlecan and vascular endothelial growth factor-encoding DNA-loaded chitosan scaffolds promote angiogenesis and wound healing. J. Control. Release 2017, 250, 48–61.10.1016/j.jconrel.2017.02.009Search in Google Scholar PubMed
[117] Gibran NS, Jang YC, Isik FF, Greenhalgh DG, Muffley LA, Underwood RA, Usui ML, Larsen J, Smith DG, Bunnett N, Ansel JC, Olerud JE. Diminished neuropeptide levels contribute to the impaired cutaneous healing response associated with diabetes mellitus. J. Surg. Res. 2002, 108, 122–128.10.1006/jsre.2002.6525Search in Google Scholar PubMed
[118] Underwood RA, Gibran NS, Muffley LA, Usui ML, Olerud JE. Color subtractive-computer-assisted image analysis for quantification of cutaneous nerves in a diabetic mouse model. J. Histochem. Cytochem. 2001, 49, 1285–1291.10.1177/002215540104901011Search in Google Scholar
[119] Spenny ML, Muangman P, Sullivan SR, Bunnett NW, Ansel JC, Olerud JE, Gibran NS. Neutral endopeptidase inhibition in diabetic wound repair. Wound Repair Regen. 2002, 10, 295–301.10.1046/j.1524-475X.2002.10504.xSearch in Google Scholar PubMed
[120] Losi P, Errico C, Lisella A, Sanguinetti E, Chiellini F, Soldani G. Fibrin-based scaffold incorporating VEGF- and bFGF-loaded nanoparticles stimulates wound healing in diabetic mice. Acta Biomater. 2013, 9, 7814–7821.10.1016/j.actbio.2013.04.019Search in Google Scholar PubMed
[121] Asrafuzzaman Md, Cao Y, Afroz R, Kamato D, Gray S, Little PJ. Animal models for assessing the impact of natural products on the aetiology and metabolic pathophysiology of Type 2 diabetes. Biomed. Pharmacother. 2017, 89, 1242–1251.10.1016/j.biopha.2017.03.010Search in Google Scholar PubMed
[122] Hardwicke JT, Hart J, Bell A, Duncan R, Thomas DW, Mosely R. The effect of dextrin–rhEGF on the healing of full-thickness, excisional wounds in the (db/db) diabetic mouse. J. Control. Release 2011, 152, 411–417.10.1016/j.jconrel.2011.03.016Search in Google Scholar PubMed
[123] Xiao Y, Reis LA, Feric N, Knee EJ, Gu J, Cao S, Laschinger C, Londono C, Antolovich J, McGuigan AP, Radisic M. Diabetic wound regeneration using peptide-modified hydrogels to target re-epithelialization. Proc. Natl. Acad. Sci. USA 2016, 113, E5792–E5801.10.1073/pnas.1612277113Search in Google Scholar
[124] Kinoda J, Ishihara M, Hattori H, Nakamura S, Fukuda K, Yokoe H. Cytotoxicity of silver nanoparticle and chitin-nanofiber sheet composites caused by oxidative stress. Nanomaterials (Basel). 2016, 6, E189.10.3390/nano6100189Search in Google Scholar PubMed
[125] Hart J, Silcock D, Gunnigle S, Cullen B, Light ND, Watt PW. The role of oxidised regenerated cellulose/collagen in wound repair: effects in vitro on fibroblast biology and in vivo in a model of compromised healing. Int. J. Biochem. Cell Biol. 2002, 34, 1557–1570.10.1016/S1357-2725(02)00062-6Search in Google Scholar
[126] Widatalla AHW, Mahadi SEI, Shawer MA, Elsayem HA, Ahmed ME. Implementation of diabetic foot ulcer classification system for research purposes to predict lower extremity amputation. Int. J. Diabetes Dev. Ctries. 2009, 29, 1–5.10.4103/0973-3930.50707Search in Google Scholar PubMed PubMed Central
[127] Galiano RD, Michaels J 5th, Dobryansky M, Levine JP, Gurtner GC. Quantitative and reproducible murine model of excisional wound healing. Wound Repair Regen. 2004, 12, 485–492.10.1111/j.1067-1927.2004.12404.xSearch in Google Scholar PubMed
[128] Translational Platforms to Model Pre-Diabetes, Diabetes, and Diabetic Complications, Cardiovascular and Metabolic Disease, White paper, May 2017, www.crownbio.com.Search in Google Scholar
[129] Breen A, Mc Redmond G, Dockery P, O’Brien T, Pandit A. Assessment of wound healing in the alloxan-induced diabetic rabbit ear model. J. Invest. Surg. 2008, 21, 261–269.10.1080/08941930802216807Search in Google Scholar PubMed
[130] Michaels J, Churgin SS, Blechman KM, Greives MR, Arani SBA, Galiano RD, Gurtner GC. db/db mice exhibit severe wound-healing impairments compared with other murine diabetic strains in a silicone-splinted excisional wound model. Wound Repair Regen. 2007, 15, 665–670.10.1111/j.1524-475X.2007.00273.xSearch in Google Scholar PubMed
[131] Seitz O, Schürmann C, Hermes N, Müller E, Pfeilschifter J, Frank S, Goren I. Wound healing in mice with high-fat diet- or ob gene-induced diabetes-obesity syndromes: a comparative study. Exp Diabetes Res. 2010, 2010, 476969.10.1155/2010/476969Search in Google Scholar PubMed PubMed Central
©2017 Walter de Gruyter GmbH, Berlin/Boston
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Articles in the same Issue
- Frontmatter
- In this issue
- Communication
- Fabrication and simulation of 1540-nm transmission by 532-nm excitation in photonic crystal of Er:ZnO film
- Self-assembly and enhanced visible-light-driven photocatalytic activity of reduced graphene oxide-Bi2WO6 photocatalysts
- Reviews
- Enhanced oral bioavailability of nisoldipine-piperine-loaded poly-lactic-co-glycolic acid nanoparticles
- Coating strategies for atomic layer deposition
- Magnetically recoverable nano-catalysts in sulfoxidation reactions
- Selective modification of inner surface of halloysite nanotubes: a review
- Analyzing polymeric nanofibrous scaffold performances in diabetic animal models for translational chronic wound healing research
- Photoluminescence from colloidal silicon nanoparticles: significant effect of surface
- Current trends in changing the channel in MOSFETs by III–V semiconducting nanostructures
Articles in the same Issue
- Frontmatter
- In this issue
- Communication
- Fabrication and simulation of 1540-nm transmission by 532-nm excitation in photonic crystal of Er:ZnO film
- Self-assembly and enhanced visible-light-driven photocatalytic activity of reduced graphene oxide-Bi2WO6 photocatalysts
- Reviews
- Enhanced oral bioavailability of nisoldipine-piperine-loaded poly-lactic-co-glycolic acid nanoparticles
- Coating strategies for atomic layer deposition
- Magnetically recoverable nano-catalysts in sulfoxidation reactions
- Selective modification of inner surface of halloysite nanotubes: a review
- Analyzing polymeric nanofibrous scaffold performances in diabetic animal models for translational chronic wound healing research
- Photoluminescence from colloidal silicon nanoparticles: significant effect of surface
- Current trends in changing the channel in MOSFETs by III–V semiconducting nanostructures