The enteric nervous system: “A little brain in the gut”
-
Anita Annahazi
Anita Annahazi studied Medicine at the University of Szeged, Hungary, where she received her MD for her thesis completed in the Cerebral Circulation Research Group of Professor Ferenc Bari. In the following years, while specializing in internal medicine at the First Department of Medicine, she joined the Gastrointestinal Motility Team. After her PhD project on the pathogenesis of irritable bowel syndrome and inflammatory bowel disease, under the supervision of Professor Tibor Wittmann and Dr Richard Roka, she pursued the development of noninvasive diagnostic approaches for intestinal diseases. As a post-doctoral fellow she spent two and a half years in the laboratory of Dr Lionel Bueno in the Neurogastroenterology and Nutrition Unit of INRA in Toulouse, France. Since 2015 she has been working at the Chair of Professor Michael Schemann at the Technical University of Munich, Germany. Currently she focuses on the role of the enteric nervous system in different gastrointestinal pathologies.
 and
Michael Schemann
and
Michael Schemann
Michael Schemann studied Agricultural Biology at the University of Hohenheim, Germany where he also received his Dr. rer. nat. with a project on the relation between intestinal motility and gastrointestinal transit under the supervision of Hans-Jörg Ehrlein. His interest in the enteric nervous system started during his post-doctoral research with Jackie Wood at Ohio State University, Columbus, Ohio, USA. Shortly after moving back to the University of Hohenheim he moved to the Max-Planck Institute for Physiological and Clinical Research in Bad Nauheim with a Heisenberg Fellowship. Before he moved to his current position as Full Professor for Human Biology at the Technical University of Munich, he was Professor at the Institute of Physiology at the University of Veterinary Medicine in Hannover. His research interests lie in the field of translational Neurogastroenterology and focus on nerve mediated gut function under physiological and pathological conditions.

Abstract
The gut’s own autonomous nervous system, the enteric nervous system (ENS), has fascinated scientists for more than 100 years. It functions, in the true sense of the word, autonomously, by performing complex tasks and controlling vital functions independently of extrinsic inputs. At the same time, the ENS is bombarded with signals from other cells in the gut wall and lumen and has to integrate all of these inputs. We describe the main functions of the ENS under physiological conditions and give a few examples of its role in gut diseases. The ENS has received increasing attention recently as scientists outside the field of Neurogastroenterology realize its important role in the pathogenesis of Parkinson’s, autism and multiple sclerosis.
Zusammenfassung
Darmfunktionen werden durch das autonom agierende enterische Nervensystem (ENS) reguliert. Es kontrolliert vitale Funktionen des Darms unabhängig von extrinsischen Einflüssen. Das ENS muss eine Fülle von Signalen anderer Zellen in der Darmwand oder Faktoren im Darmlumen integrieren. In diesem Artikel beschreiben wir die wesentlichen Funktionen des ENS und erläutern Beispiele aus der klinischen Neurogastroenterologie. Darüber hinaus eröffnen sich neue Aspekte für das Verständnis systemischer neurologischer Erkrankungen wie Parkinson, Autismus oder Multipler Sklerose, bei denen die Rolle des Darms und des ENS immer offensichtlicher wird.
Introduction
The gastrointestinal tract (GI) fulfils complex tasks that are essential for survival. Apart from mechanical and chemical digestion of food, transit of luminal content, and absorption of nutrients, it functions as an important immune organ by recognizing and fighting luminal pathogens. These complicated processes are regulated by a unique autonomic network of neurons called the enteric nervous system (ENS). Although the ENS communicates with the central nervous system (CNS) as well as sympathetic and parasympathetic nerves, it operates independently. This is why an isolated intestinal segment separated from all external input behaves as if it would do inside the body. It is because of such abilities that the ENS is often referred to as a “second brain”. The Hydra provides evolutionary evidence that the ENS was present before a CNS (Furness and Stebbing, 2018). This animal has a net-like nervous system located in the wall of the gut tube, which does not form brain-like aggregations. This nervous system contains sensory, motor and interneurons, is related to the circular and longitudinal layers of the body wall of the Hydra, and controls movements needed for digestion: peristalsis, mixing and expulsion. Consequently, it must be considered as an ENS-equivalent, while no CNS is present in this species (Shimizu et al., 2004). It is therefore fair to assume that the brain is an encephalized ENS, with the latter being considered as the “first brain” to appear during evolution. The objectives of this review are to present an overview of the structure and function of the mammalian “little brain”, and to describe some of the important pathologies caused by its dysfunction.
Anatomy of the ENS
There are around 200–600 million nerve cells in the mammalian ENS which are organized, along with glial cells, into interconnected ganglia. The ENS extends from the esophagus to the anal sphincter and has branches to the liver, gall bladder, biliary tract and pancreas. The ganglia form neuronal networks – the so-called plexuses – that are interconnected (Figure 1). The myenteric plexus lies between the longitudinal and circular layers and controls muscle activity. The submucous plexuses are located beneath the mucosa and control epithelial functions. Both plexuses modulate blood flow and the activity of the enteric immune system. The ENS develops mainly from vagal neural crest cells, which migrate into and along the bowel over 5 gestational weeks in humans (Stamp, 2017). A small number of neurons, however, originate from sacral neural crest cells and Schwann cell precursors.
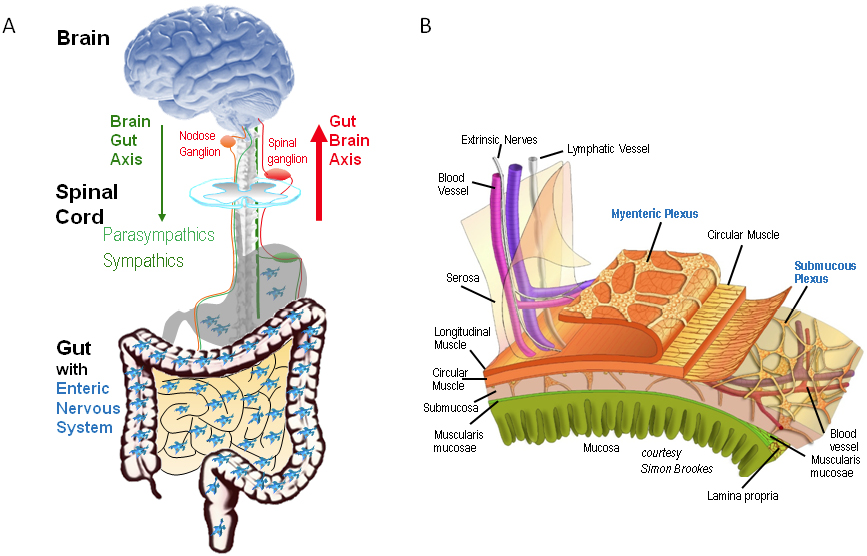
Functional anatomy of gut-brain communication and the enteric nervous system (ENS).
Panel A is a simplified scheme to demonstrate control of gut functions at different levels. The most relevant is the ENS, which controls gut functions independent of extrinsic inputs. Nevertheless, the gut is connected to the brain via nerves which function as the gut-brain axis – sensory neurons with cell bodies in dorsal root ganglia or in parasympathetic relay ganglia, e. g. nodose ganglion, and efferent nerves of the parasympathetic and sympathetic nervous systems. Note that there is no region in the brain exclusively dedicated to gut functions. Panel B illustrates the myenteric and submucous plexus layers of the ENS in the gut wall.
The figure was provided by Prof. Simon Brookes, Flinders University, Adelaide, Australia.
Central connections
Although the ENS is able to function autonomously, connections between the ENS and CNS exist and are referred to as the gut-brain axis (Furness and Stebbing, 2018; Schemann and Grundy, 1992; Mayer, 2011; see Figure 1). The gut’s extrinsic nerve supply carries efferent as well as afferent nerves and uses the ENS as an interface. This explains why parasympathetic nerves support motility and secretion during the digestive period, whereas sympathetic nerves inhibit those functions through presynaptic inhibition at synapses within the ENS (Furness et al., 2014). The vast majority of nerves travelling with vagal and spinal trunks are afferents, which, apart from transmitting sensory inputs from the GI tract to the CNS, have collateral branches to blood vessels and enteric ganglia, providing local axon reflexes. Further connection occurs through intestinofugal neurons in the ENS, which project to the sympathetic postganglionic neurons within prevertebral ganglia and even to the trachea, gall bladder and pancreas (Furness and Stebbing, 2018). Furthermore, the brain also provides input to the ENS via hormonal pathways, such as the hypothalamic-pituitary-adrenal (HPA) and sympatho-adrenal axes, and descending monoaminergic pathways (Mayer, 2011). A significant part of signaling from the gut to the brain is performed by enteroendocrine cells (EEC). They are located in the gut wall and represent the largest endocrine organ in the body, serving to detect mechanical (e. g. shear forces) as well as chemical stimuli, including nutrients, microbial products or toxins. Communication to the CNS is via blood or sensory nerves travelling with the sympathetic or parasympathetic trunks (Latorre et al., 2016). There are more than 20 types of EECs, producing different endocrine and paracrine mediators, which participate in the regulation of GI motility and secretion, pancreatic enzyme and bile secretion, and the regulation of food intake.
Function of the ENS
Despite the connections between the CNS and the ENS, the vital functions of the GI tract almost completely depend on the ENS, while the CNS monitors gut activity and may, at most, modulate it (Furness et al., 2014). The total depletion of the ENS, which occurs in cases of the congenital lack of enteric ganglia in a colonic segment in Hirschsprung’s disease (detailed later) is lethal, despite supply by extrinsic nerves remaining. The most prominent function of the myenteric plexus is the regulation of the motility of the circular and longitudinal muscles. The submucous plexus mainly controls ion and water secretion, absorption of ions, vitamins and nutrients, as well as release of endocrine and paracrine mediators.
The ENS contains sensory neurons, interneurons and motor neurons, but these are often multifunctional and fulfil various tasks (Furness et al., 2014; Kugler et al., 2015; Smith et al., 2007). Sensory neurons are able to detect mechanical and chemical stimuli. Mechanosensitive ENS neurons respond to tensile and compressive stress rather than to shear stress (Mazzuoli-Weber and Schemann, 2015). The chemosensitive ENS neurons express receptors for amino acids, fatty acids, glucose, pH, osmolarity, temperature, odorants and tastes (Blackshaw et al., 2007; Neunlist et al., 2001; Bertrand et al., 1997). Some glucose-sensitive enteric neurons show behavior reminiscent of glucosensitive and glucoresponsive neurons in the hypothalamus (Liu et al., 1999).
Studies investigating the presence of neurotransmitters, neuropeptides and neuromodulators revealed that identical types occur in the CNS and ENS. It is assumed that the complexity of transmitter and receptor expression is similar, if not identical, between the ENS and CNS (Furness, 2006). For example, unlike in the CNS, glycine acts as an excitatory transmitter in the adult ENS (Neunlist et al., 2001). Some of the signaling molecules released by the ENS are primary transmitters and others serve modulatory roles. In Table 1 we list only those that are considered primary transmitters as they are central to the initiation and maintenance of reflex activated muscle or epithelial activity. Sensory neurons innervate motor neurons directly or via inhibitory or excitatory interneurons. Excitatory or inhibitory motor neurons project to the various muscle layers or epithelium to affect motility or secretion, respectively. Motor neurons also innervate enteric endocrine cells and lymphoid aggregations in the GI tract (Furness et al., 2014).
We recently reviewed the evidence that the ENS is able to perform higher functions. Although a rigid scientific proof or disproof is still required, it seems that the ENS is able to learn, memorize and forget (Schemann et al., 2019). All the electrophysiological and molecular proxies for habituation, facilitation and conditioned learning exist in the ENS. If this proves to be correct, it may revolutionize the way we interpret plasticity in the ENS and pathogenesis of gut diseases (Schemann et al., 2019).
The peristaltic reflex
Motility of the GI organs is indispensable for survival (Huizinga and Lammers, 2009). The basis for all motility patterns is the peristaltic reflex, which was described as early as the end of the 19th century (Bayliss and Starling, 1899; Lüderitz, 1890; see Figure 2). Lüderitz provided a comprehensive description of peristalsis as the basis for proximal to distal movement of content for the first time in anesthetized animals. He noticed that mechanical distension induced a response which consisted of a contraction proximal to the distension and muscle inhibition distal to the distension, and suggested that the ENS triggers this reflex. Bayliss and Starling confirmed the validity of this hypothesis by observing the peristaltic reflex even when all connections to the CNS were interrupted (Figure 2).
Today we know that the first step in this process is the activation of sensory neurons in the ENS by mechanical and/or chemical stimuli. Activation builds up in a sensory interneuronal network and eventually activates excitatory motor neurons projecting up the gut and inhibitory motor neurons projecting down the gut. This results in the release of acetylcholine proximal to the stimulus and nitric oxide (NO), adenosine triphosphate (ATP) and/or vasoactive intestinal peptide (VIP) distal to the stimulus. The axonal projections of these motor neurons are rather short, spanning about 1cm, which means that the peristaltic reflex has to be sequentially activated in cases where fast transit is appropriate (Figure 3). In contrast, sequential activation is halted if transit needs to be slowed down during the digestive period. Thus, the peristaltic reflex is not an all-or-nothing response, but instead is highly modulated. Interneurons with descending projections are required to initiate sequential activation. Inhibition of interneuronal synapses, the presence of synapses between interneurons and motor neurons, or a decrease in the sensitivity of sensory neurons are all means to halt sequential activation (Figure 3).
Summary of the functions of some of the main transmitters in the ENS.
|
Transmitter |
Released from |
Target (Receptor) |
Function |
|
Acetylcholine (ACh) |
Muscle motor neuron Secretomotor neuron Sensory neuron / Interneuron Parasympathetic nerves |
Muscle (muscarinic) Epithelial secretion (muscarinic) Enteric neurons (nicotinic) Enteric neurons (nicotinic) Enteric neurons (nicotinic) |
Promotility Prosecretory Enhance nerve activity Enhance nerve activity Enhance transmitter release from enteric nerves |
|
Substance P |
Muscle motor neuron Sensory neuron / Interneuron |
Muscle (NK-2) Enteric neurons (NK-1,2,3) |
Promotility Enhance nerve activity |
|
ATP, β-nicotinamide adenine dinucleotide |
Muscle motor neuron Interneuron |
Muscle (P2Y1) Interneuron, Motor neuron (P2X) |
Antimotility Enhance nerve activity |
|
Nitric Oxide |
Motoneuron Secretomotor neuron Interneuron |
Muscle (increased cGMP) Epithelial secretion Enteric neurons (presynaptic inhibition of non-cholinergic non adrenergic transmitters) |
Antimotility Prosecretory Inhibit nerve activity (slow EPSP) |
|
Vasoactive Intestinal Peptide |
Muscle motor neuron Secretomotor neuron |
Muscle Epithelial secretion (VPAC2) |
Antimotility Prosecretory |
|
Neuropeptide Y |
Interneuron Secretomotor neuron |
Presynaptic inhibition of ACh release Epithelial secretion |
Inhibit nerve activity (fast EPSP) Antisecretory |
|
Enkephaline Endorphin |
Interneuron Muscle motor neuron Secretomotor neuron |
Presynaptic inhibition of ACh release (µ,δ,κ) Muscle Epithelial secretion |
Inhibit nerve activity (fast EPSP) Antimotility Antisecretory |
|
Serotonin (5-HT) |
Sensory neuron / Interneuron |
Presynaptic potentiation of ACh release (5-HT4) Enteric motor and interneurons (5-HT3) Excitatory motor neuron (5-HT1p) Inhibitory motor neuron (5-HT1p) |
Enhance nerve activity (fast EPSP) Enhance nerve activity Enhance peristalsis Enhance peristalsis |
|
Noradrenaline |
Sympathetic nerves |
Enteric interneurons, presynaptic inhibition of ACh release (α2) |
Inhibition of enteric neurons |
The actual motility pattern is modified by additional factors, such as the activity of pacemaker cells (Interstitial cells of Cajal), hormones, immune mediators, and sympathetic and parasympathetic inputs. This in turn creates the complex gut movements of propulsion, segmentation and storage which permit the digestion and absorption of nutrients (Mazzuoli-Weber and Schemann, 2015; Huizinga and Lammers, 2009).
Pathologies of the ENS
Diseases linked to disorders of the gut are diverse in their symptoms and severity. They can range from bothersome, but not life threatening, disturbances of motility, such as in irritable bowel syndrome (IBS), to the potentially fatal motility impairments seen in Hirschsprung’s disease. There are numerous diseases in which the ENS is expected to play a role (Table 2). Strikingly, it turns out that the ENS is involved in the pathogenesis of diseases not commonly linked to the gut, such as multiple sclerosis, autism, Parkinson’s disease and cancer. A common feature of functional gut disorders is that therapy is, all in all, unsatisfying, and causal therapies are particularly lacking. As the discussion of all ENS pathologies is far beyond the scope of this review, some diseases have been selected to provide a glimpse into the diverse pathomechanisms.
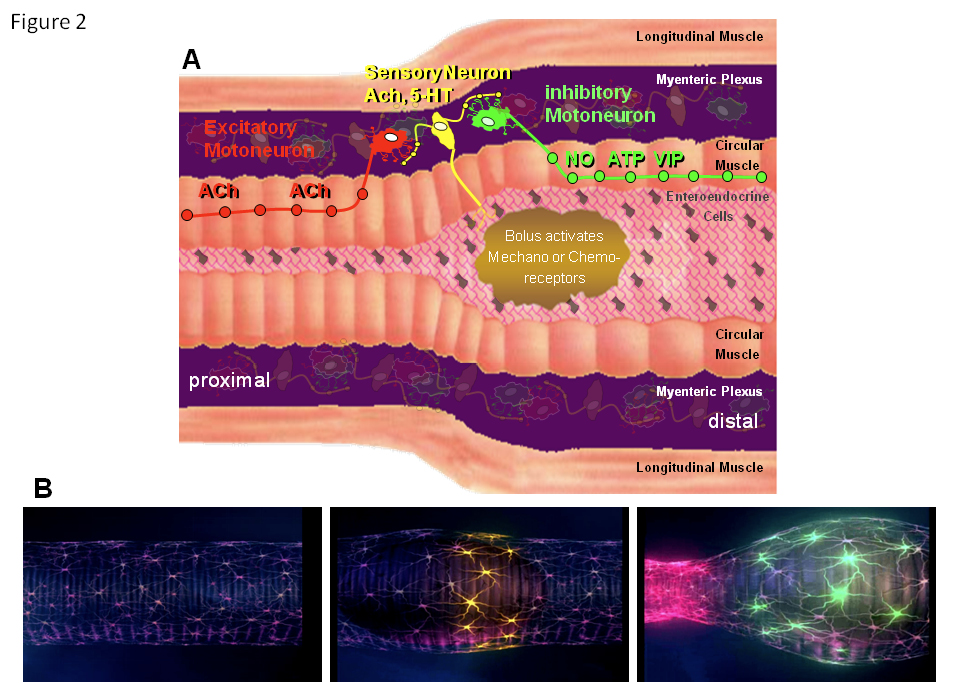
Circuits in the ENS activate the peristaltic reflex.
Panel A shows a schematic drawing of the intestinal wall with the different layers. Circuits in the myenteric plexus responsible for initiating the peristaltic reflex are highlighted. Initially, sensory neurons in the ENS are activated by mechanical or chemical stimuli. They then activate excitatory and inhibitory muscle motor neurons with polarized projection patterns. The inhibitory motor neurons project down the gut and release nitric oxide, ATP and vasoactive intestinal peptide, whereas the excitatory motor neurons project up the gut to release acetylcholine. The projection length for both is about 1cm, maximum. This circuit guarantees that a bolus triggers a proximal contraction and a distal inhibition of the muscle, allowing the content to be pushed in an anal direction. For simplicity, interneurons are not shown (see Figure 3).
Panel B shows the spatiotemporally coordinated activity within the circuit. It starts with quiescence (left). As soon as the bolus enters the region, mechanosensitive neurons (middle; yellow) start to fire actions potentials. This will eventually lead to activation of excitatory (right; red) and inhibitory motor neurons (right; green).
One of the most devastating gut motility disorders is Hirschsprung’s disease, also called congenital megacolon or intestinal aganglionosis, and first described by Harald Hirschsprung in 1888 (Hirschsprung, 1888; Sergi, 2015). The underlying histology was discovered shortly after the initial description and consists of sparse or lacking ganglia in the colonic ENS but a normal ganglionic network in the ileum (Tittel, 1901). It is now known that the disease is caused by the disruption of normal neural crest cell migration or development (Butler Tjaden and Trainor, 2013). The resulting aganglionosis may be limited to narrow segments of the gut or extend to the entire colon. The lack of ganglia causes tonic contraction of the affected segment, resulting in a bowel obstruction, and typically appearing immediately after birth. The contraction is the result of continuous uncontrolled release of acetylcholine from extrinsic parasympathetic nerves and the lack of coordinated release of inhibitory transmitters of the ENS. The newborn thus fails to pass meconium in the first 24 hours. The routine treatment in Hirschsprung’s disease is the removal of the aganglionic segment, but as a late outcome of this life-saving procedure patients must deal with bowel disorders that result in reduced bowel-related quality of life during adulthood (Gustafson et al., 2019).
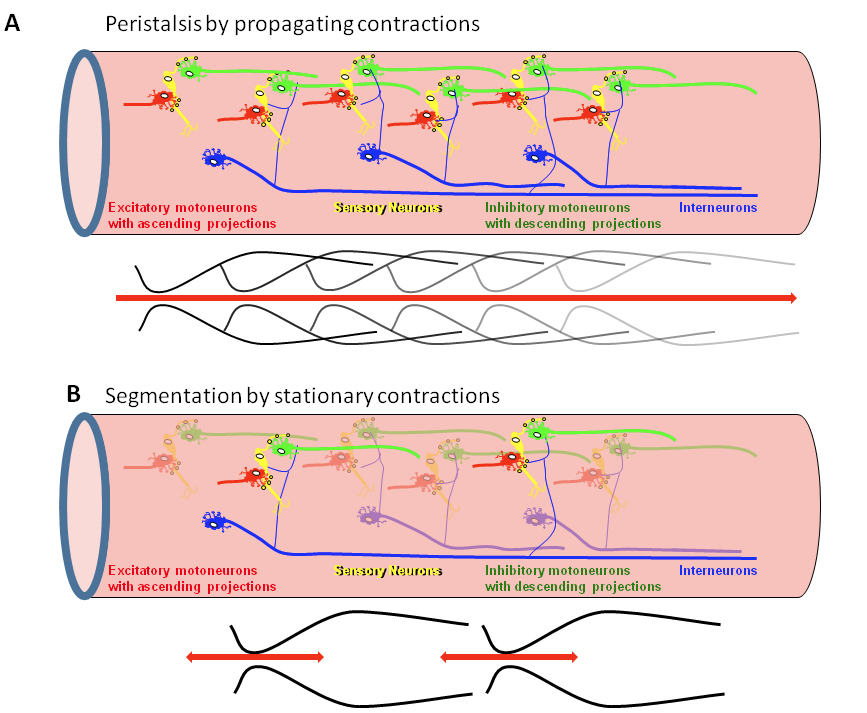
A model to explain how spatiotemporally coordinated peristaltic reflex circuits in the ENS lead to peristalsis.
Panel A shows a chain of peristaltic reflex circuits consisting of sensory neurons synapsing onto excitatory and inhibitory muscle motor neurons. The circuits are connected via excitatory interneurons which sensitize the peristaltic reflex circuits, thus allowing peristalsis and movement of intestinal content over longer distances.
Panel B shows some interneurons failing to sensitize consecutive peristaltic reflex circuits. This will prevent the spread of activation along the circuits. The result is disrupted propagation of peristaltic reflexes causing stationary contractions.
Recently, some promising results of experiments with ENS stem cell transplantation using optogenetic techniques have been published (Wang, 2018). Optogenetics uses a genetically implanted light sensitive channel to detect or control the activity of a specific cell type (Deisseroth et al., 2006). In a mouse model, transplanted enteric neural cells expressing the light-sensitive ion channel, channelrhodopsin, formed a ganglionated network and upon activation by light stimulus induced inhibitory or excitatory electrical events in the circular muscle (Stamp et al., 2017). Furthermore, enteric neuronal precursor cells isolated with magnetic immunoselection and transplanted into the colons of Piebald mice (a model of Hirschsprung’s disease with a reduced number of colonic neurons and an aganglionic distal colonic segment) or nNOS−/− mice successfully improved colonic contractility and relaxation, respectively (Ro et al., 2006; Anitha et al., 2008). In a study aiming to evaluate the feasibility of autologous transplantation in ENS disorders, neural crest progenitors were isolated from neonatal rats and transplanted into the chemically denervated distal colon of other rats (Pan et al., 2011). The transplanted cells successfully colonized the gut and reversed motility impairments induced by denervation.
Diseases associated with pathologies of the ENS or the gut-brain axis.
|
“Classical” gut diseases |
“Non-classical” gut diseases |
|
Achalasia Amyloidosis Chronic intestinal pseudoobstruction Diabetes Inflammatory Bowel Disease Functional gastrointestinal diseases; e. g. irritable bowel syndrome, functional dyspepsia Hirschsprung’s Disease Ileus Intoxication Food allergy or intolerance Mast cell mediator syndrome Slow transit constipation Tumor development (colon cancer) |
Alzheimer’s Disease Autism spectrum disorder Multiple sclerosis Paraneoplastic disorder Parkinson’s Disease Transmissible spongiform encephalopathy Viral or bacterial Infection |
Irritable bowel syndrome is a chronic functional GI disorder with an outstandingly high global prevalence of 11.2 % (Enck et al., 2016). Although the disease is not life threatening, with its distressing symptoms of abdominal pain, bloating and altered bowel habits, as well as limited, mainly symptomatic, therapy, it has devastating consequences for quality of life. Patients are classified according to their stool patterns as IBS-D (IBS with predominant diarrhea), IBS-C (IBS with predominant constipation) or IBS-M (IBS with mixed stool pattern) (Drossman, 2016). IBS is a multifactorial disease with numerous putative pathologies, including ENS disturbances and altered signaling along the gut-brain axis (Enck et al., 2016).
Physical or psychological stress, with its ability to disturb the hypothalamic-pituitary-adrenal axis, is a major player in IBS development and relapse (Moloney et al., 2015; Jahng and Kim, 2016). Stress induces histological and functional changes in the ENS (Li et al., 2016b). An increased number of mast cells in the lamina propria, a known phenomenon in IBS patients, has also been demonstrated in IBS models (Traini et al., 2016; O’Sullivan et al., 2000). It was shown that mast cells of IBS-D patients release more mediators, such as histamine or mast cell tryptase, that are capable of neuronal activation, and the number of mast cells in close proximity to nerves significantly correlated with severity and frequency of abdominal pain and/or discomfort in IBS patients (Barbara et al., 2004). In line with these findings, biopsy samples from the colon or rectum of IBS patients released proteases with 2 to 3 fold higher activity than biopsies from controls, and these biopsy supernatants with elevated protease activity caused hyperalgesia and allodynia in response to colorectal distension after intracolonic injection in mice (Cenac et al., 2007). It has been demonstrated that biopsy supernatants of IBS patients are able to activate enteric neurons due to their protease, histamine and serotonin content (Buhner et al., 2009; Buhner et al., 2012). Furthermore, biopsy supernatants of patients who showed hypersensitivity to rectal distension produced significantly stronger activation in submucous and dorsal root ganglion neurons (Buhner et al., 2014). Interestingly, submucous neurons in biopsies obtained from IBS patients responded significantly less strongly to a cocktail of compounds mimicking IBS biopsy supernatants (a mixture of serotonin, histamine, tryptase, and tumor necrosis factor alpha (TNF-α)) than neurons in biopsies from healthy controls, suggesting desensitization caused by the constant release of these mediators (Ostertag et al., 2015). Similarly to the increased protease activity in mucosal biopsy supernatants, IBS-D patients show elevated fecal serine-protease activity compared to healthy controls or IBS-C patients, which may be a factor triggering epithelial barrier dysfunction and visceral hypersensitivity (Róka et al., 2007; Gecse et al., 2008). An increased cysteine-protease activity has been found in fecal samples of a subgroup of IBS-C patients, which may play a role in the disruption of the intestinal barrier and visceral hypersensitivity (Annaházi et al., 2013). These findings offer new therapeutic options in IBS that target mast cell activation and degranulation, and mast cell products and receptors, some of which have already shown benefits in clinical trials (Zhang et al., 2016).
Structural changes in the ENS in IBS have led to suggestions by some authors that this may be an autoimmune phenomenon (Wood et al., 2012; Fan et al., 2018). Anti-enteric neuronal antibodies were found in the sera of a higher percentage of IBS patients than controls, and such sera induced apoptosis in guinea pig myenteric neurons (Wood et al., 2012; Fan et al., 2018). The authors suggest that a subgroup of patients with high anti-enteric neuronal antibody titer may benefit from antibody-depleting therapies (Fan et al., 2018).
In recent years, growing evidence supports the theory that the ENS, and in particular enteric neuropathies, are important players in several diseases that were previously considered to primarily affect the CNS, with ENS disorders representing risk factors in many cases. It is well known that bovine spongiform encephalopathy and kuru have their origin in the gut. Namely, the ingestion of infectious amyloids, so-called prions, transfers the disease by homologous seeding – in other words, by oligomers of the misfolded protein promoting the aggregation of that same protein.
Interestingly, it has been suggested that a similar pathogenesis, cross-seeding, may account for Parkinson’s disease (PD). In this case, a specific misfolded protein induces the polymerization of a different protein (for a review, see Friedland and Chapman, 2017; Chapelet et al., 2019). In the brain, alpha-synuclein aggregates consisting of misfolded proteins are indicative of PD, but are also found in the myenteric plexus of these patients during autopsy. Furthermore, most PD patients suffer from GI symptoms besides neurological deficits. These observations led to Braak and colleagues’ hypothesis that the disease may start in the stomach (part of the ENS) with aggregation of alpha-synuclein due to an environmental pathogen (Braak et al., 2006). The pathological process may reach the brain through the vagus nerve. Although this hypothesis is disputed (Lionnet et al., 2018), it has been shown that vagotomy indeed decreases the risk of PD (Svensson et al., 2015). Several bacterial species, for example E. coli, Staphylococcus, Streptococcus and Salmonella, produce extracellular amyloids that are capable of cross-seeding (Friedland and Chapman, 2017). Aged rats exposed to amyloid producing E. coli developed alpha-synuclein deposits both in their gut and brain, accompanied by microgliosis and astrogliosis (Chen et al., 2016). This concept is supported by the fact that PD patients exhibit changes in their gut microbiota not seen in healthy individuals (Chapelet et al., 2019). Furthermore, in animal studies, the gut microbiota has been shown to play a role in the development of neurological symptoms (Choi et al., 2018; Sampson et al., 2016). The involvement of the GI tract is also supported by the fact that certain major susceptibility genes for inflammatory bowel diseases are over-represented (Bialecka et al., 2007; Hui et al., 2018) in PD patients, and gut inflammation (Devos et al., 2013) and impaired intestinal barrier function (Clairembault et al., 2015; Davies et al., 1996) may also be present in at least a subset of PD patients. Today, it is widely accepted that in some PD patients the disease may start in the gut and be clinically evident as severe constipation before the CNS-triggered symptoms occur.
Another disorder of the CNS that affects the ENS is multiple sclerosis (MS). GI problems are very common among MS patients, yet the pathogenesis is distinct from that of PD. In a cohort of 218 patients, almost two thirds suffered from GI symptoms (Levinthal et al., 2013). Some of these symptoms, such as dysphagia or defecation problems, can be explained by the underlying musculoskeletal dysfunction, others, like nausea and vomiting, may be side effects of medical therapy. Nevertheless, a surprisingly high rate of patients show symptoms corresponding to functional gastrointestinal disorders, such as functional dyspepsia, functional constipation or IBS. GI pathologies were previously thought to be primarily explained by spinal cord lesions causing dysfunction of the autonomous nervous system. However, recently it has been suggested that the autoimmune process, which attacks the CNS, directly affects the ENS (Wunsch et al., 2017). In a mouse model of MS called experimental autoimmune encephalomyelitis (EAE), macrophages and T- and B-lymphocytes were observed in the myenteric plexus of the gut, even in the preclinical state. The invasion of immune cells was accompanied by a degeneration of the myenteric plexus, which preceded the degeneration of the spinal cord. GI transit time was significantly decreased and muscular cholinergic signaling and NO release were reduced in this MS model. The majority of mice with EAE had ENS-reactive autoantibodies, while antibodies against a component of the ENS could be detected in 10 of 33 human MS patients. These results were corroborated by another study showing accelerated gastric emptying and reduced colonic motility in EAE mice, accompanied by immunoreactivity against the ENS in sera (Spear et al., 2018).
A further CNS disorder that has recently been shown to simultaneously affect the ENS is autism spectrum disorder (ASD). GI symptoms in patients with ASD are very common, and they correlate strongly with disease severity (Adams et al., 2011). The GI tract is an important source of serotonin (5-HT), which, besides other functions, plays a role in GI motility. Almost one third of ASD patients have hyperserotonaemia, leading to a link between 5-HT and the GI symptoms of autism patients being suspected (for a review see: Israelyan and Margolis, 2019). Some gain-of-function mutations of the gene encoding the serotonin reuptake transporter, SERT, are associated with ASD, and a murine model with one of these mutations, the “SERT Ala56 mouse”, shows brain and behavioral anomalies typical of ASD. Interestingly, these mice also present with abnormalities of the ENS, such as a low neuronal count, slow intestinal motility and reduced frequency and speed of colonic peristaltic contractions (Margolis et al., 2016). Similarly to in other CNS disorders, altered microbial flora has been observed in ASD patients and is suspected to contribute to symptom development – probiotic treatments have therefore been suggested (Hsiao et al., 2013). Furthermore, it has been hypothesized that increased intestinal permeability, gut inflammation and motility impairments may be explained by enteric glial cell dysfunction (Grubišić and Parpura, 2015).
The role of the gut-brain axis, which includes the ENS, in carcinogenesis is underestimated and vastly understudied. For example, colon tumor development in the Apc(Min/+) mouse model is inhibited after vagotomy, but not by sympathetic denervation (Liu et al., 2015). Last but not least, the ENS is the target of paraneoplastic neurological autoimmunity often associated with small cell lung cancer. Autoantibodies against the Hu-antigen, which are released by the tumor, attack enteric neurons as the vast majority of them express Hu proteins. Chronic exposure of enteric neurons to anti-Hu containing patient serum causes neuronal damage. Even more fascinating is the finding that acute application of the patient serum for only a few hundreds of milliseconds evokes immediate spike discharge involving interaction between anti-HuD and nicotinic receptors on enteric neurons (Li et al., 2016a).
Summary and outlook
Neurogastroenterology, which interested only a small group of experts in the past, has been gradually coming into the spotlight and gaining visibility among both medical/scientific audiences and the lay public. Apart from known structural and functional disorders of the ENS, microbiome-gut-brain axis disturbances are now linked to a constantly growing range of pathologies, including Parkinson’s disease, autism spectrum disorder and multiple sclerosis. A better understanding of the physiology of the enteric nervous system and the pathogenesis of ENS disorders gained over recent years may lead to the development of new therapies.
About the authors

Anita Annahazi studied Medicine at the University of Szeged, Hungary, where she received her MD for her thesis completed in the Cerebral Circulation Research Group of Professor Ferenc Bari. In the following years, while specializing in internal medicine at the First Department of Medicine, she joined the Gastrointestinal Motility Team. After her PhD project on the pathogenesis of irritable bowel syndrome and inflammatory bowel disease, under the supervision of Professor Tibor Wittmann and Dr Richard Roka, she pursued the development of noninvasive diagnostic approaches for intestinal diseases. As a post-doctoral fellow she spent two and a half years in the laboratory of Dr Lionel Bueno in the Neurogastroenterology and Nutrition Unit of INRA in Toulouse, France. Since 2015 she has been working at the Chair of Professor Michael Schemann at the Technical University of Munich, Germany. Currently she focuses on the role of the enteric nervous system in different gastrointestinal pathologies.

Michael Schemann studied Agricultural Biology at the University of Hohenheim, Germany where he also received his Dr. rer. nat. with a project on the relation between intestinal motility and gastrointestinal transit under the supervision of Hans-Jörg Ehrlein. His interest in the enteric nervous system started during his post-doctoral research with Jackie Wood at Ohio State University, Columbus, Ohio, USA. Shortly after moving back to the University of Hohenheim he moved to the Max-Planck Institute for Physiological and Clinical Research in Bad Nauheim with a Heisenberg Fellowship. Before he moved to his current position as Full Professor for Human Biology at the Technical University of Munich, he was Professor at the Institute of Physiology at the University of Veterinary Medicine in Hannover. His research interests lie in the field of translational Neurogastroenterology and focus on nerve mediated gut function under physiological and pathological conditions.
References
Adams, J.B., Johansen, L.J., Powell, L.D., Quig, D., and Rubin, R.A. (2011). Gastrointestinal flora and gastrointestinal status in children with autism – comparisons to typical children and correlation with autism severity. BMC Gastroenterol. 11, 22.10.1186/1471-230X-11-22Search in Google Scholar PubMed PubMed Central
Anitha, M., Joseph, I., Ding, X., Torre, E.R., Sawchuk, M.A., Mwangi, S., … Srinivasan, S. (2008). Characterization of Fetal and Postnatal Enteric Neuronal Cell Lines With Improvement in Intestinal Neural Function. Gastroenterology 134, 1424–1435.10.1053/j.gastro.2008.02.018Search in Google Scholar PubMed PubMed Central
Annaházi, A., Ferrier, L., Bézirard, V., Lévêque, M., Eutamène, H., Ait-Belgnaoui, A., … Bueno, L. (2013). Luminal Cysteine-Proteases Degrade Colonic Tight Junction Structure and Are Responsible for Abdominal Pain in Constipation-Predominant IBS. Am. J. Gastroenterol. 108, 1322–1331.10.1038/ajg.2013.152Search in Google Scholar PubMed
Barbara, G., Stanghellini, V., De Giorgio, R., Cremon, C., Cottrell, G.S., Santini, … Corinaldesi, R. (2004). Activated mast cells in proximity to colonic nerves correlate with abdominal pain in irritable bowel syndrome. Gastroenterology 126, 693–702.10.1053/j.gastro.2003.11.055Search in Google Scholar PubMed
Bayliss, W.M., and Starling, E.H. (1899). The movements and innervation of the small intestine. J. Physiol. 24, 99–143.10.1113/jphysiol.1899.sp000752Search in Google Scholar PubMed PubMed Central
Bertrand, P.P., Kunze, W.A., Bornstein, J.C., Furness, J.B., and Smith, M.L. (1997). Analysis of the responses of myenteric neurons in the small intestine to chemical stimulation of the mucosa. Am. J. Physiol.: Gastrointest. Liver Physiol. 273, G422–G435.10.1152/ajpgi.1997.273.2.G422Search in Google Scholar
Bialecka, M., Kurzawski, M., Klodowska-Duda, G., Opala, G., Juzwiak, S., Kurzawski, G., Tan, E.-K., and Drozdzik, M. (2007). CARD15 variants in patients with sporadic Parkinson’s disease. Neurosci. Res. 57, 473–476.10.1016/j.neures.2006.11.012Search in Google Scholar PubMed
Blackshaw, L.A., Brookes, S.J.H., Grundy, D., and Schemann, M. (2007). Sensory transmission in the gastrointestinal tract. Neurogastroenterol. Motil. 19, 1–19.10.1111/j.1365-2982.2006.00871.xSearch in Google Scholar PubMed
Braak, H., de Vos, R.A.I., Bohl, J., and Del Tredici, K. (2006). Gastric α-synuclein immunoreactive inclusions in Meissner’s and Auerbach’s plexuses in cases staged for Parkinson’s disease-related brain pathology. Neurosci. Lett. 396, 67–72.10.1016/j.neulet.2005.11.012Search in Google Scholar PubMed
Buhner, S., Li, Q., Vignali, S., Barbara, G., De Giorgio, R., Stanghellini, V. … Schemann, M. (2009). Activation of Human Enteric Neurons by Supernatants of Colonic Biopsy Specimens From Patients With Irritable Bowel Syndrome. Gastroenterology 137, 1425–1434.10.1053/j.gastro.2009.07.005Search in Google Scholar PubMed
Buhner, S., Li, Q., Berger, T., Vignali, S., Barbara, G., De Giorgio, R., Stanghellini, V., and Schemann, M. (2012). Submucous rather than myenteric neurons are activated by mucosal biopsy supernatants from irritable bowel syndrome patients: Enteric plexuses and IBS supernatants. Neurogastroenterol. Motil. 24, 1134-e572.10.1111/nmo.12011Search in Google Scholar PubMed
Buhner, S., Braak, B., Li, Q., Kugler, E.M., Klooker, T., Wouters, M., … Schemann, M. (2014). Neuronal activation by mucosal biopsy supernatants from irritable bowel syndrome patients is linked to visceral sensitivity: Neuronal activation in irritable bowel syndrome. Exp. Physiol. 99, 1299–1311.10.1113/expphysiol.2014.080036Search in Google Scholar PubMed
Butler Tjaden, N.E., and Trainor, P.A. (2013). The developmental etiology and pathogenesis of Hirschsprung disease. Transl. Res. 162, 1–15.10.1016/j.trsl.2013.03.001Search in Google Scholar PubMed PubMed Central
Cenac, N., Andrews, C.N., Holzhausen, M., Chapman, K., Cottrell, G., Andrade-Gordon, P., … Vergnole, N. (2007). Role for protease activity in visceral pain in irritable bowel syndrome. J. Clin. Invest. 117, 636–647.10.1172/JCI29255Search in Google Scholar PubMed PubMed Central
Chapelet, G., Leclair-Visonneau, L., Clairembault, T., Neunlist, M., and Derkinderen, P. (2019). Can the gut be the missing piece in uncovering PD pathogenesis? Parkinsonism Relat. Disord. 59, 26–31.10.1016/j.parkreldis.2018.11.014Search in Google Scholar PubMed
Chen, S.G., Stribinskis, V., Rane, M.J., Demuth, D.R., Gozal, E., Roberts, A.M., … Friedland, R.P. (2016). Exposure to the Functional Bacterial Amyloid Protein Curli Enhances Alpha-Synuclein Aggregation in Aged Fischer 344 Rats and Caenorhabditis elegans. Sci. Rep. 6, 34477.10.1038/srep34477Search in Google Scholar PubMed PubMed Central
Choi, J.G., Kim, N., Ju, I.G., Eo, H., Lim, S.-M., Jang, S.-E., Kim, D.-H., and Oh, M.S. (2018). Oral administration of Proteus mirabilis damages dopaminergic neurons and motor functions in mice. Sci. Rep. 8, 1275.10.1038/s41598-018-19646-xSearch in Google Scholar PubMed PubMed Central
Clairembault, T., Leclair-Visonneau, L., Coron, E., Bourreille, A., Le Dily, S., Vavasseur, F., … Derkinderen, P. (2015). Structural alterations of the intestinal epithelial barrier in Parkinson’s disease. Acta Neuropathol. Commun. 3, 12.10.1186/s40478-015-0196-0Search in Google Scholar PubMed PubMed Central
Davies, K.N., King, D., Billington, D., and Barrett, J.A. (1996). Intestinal permeability and orocaecal transit time in elderly patients with Parkinson’s disease. Postgrad. Med. J. 72, 164–167.10.1136/pgmj.72.845.164Search in Google Scholar PubMed PubMed Central
Deisseroth, K., Feng, G., Majewska, A.K., Miesenbock, G., Ting, A., and Schnitzer, M.J. (2006). Next-Generation Optical Technologies for Illuminating Genetically Targeted Brain Circuits. J. Neurosci. 26, 10380–10386.10.1523/JNEUROSCI.3863-06.2006Search in Google Scholar PubMed PubMed Central
Devos, D., Lebouvier, T., Lardeux, B., Biraud, M., Rouaud, T., Pouclet, … Derkinderen, P. (2013). Colonic inflammation in Parkinson’s disease. Neurobiol. Dis. 50, 42–48.10.1016/j.nbd.2012.09.007Search in Google Scholar PubMed
Drossman, D.A. (2016). Functional Gastrointestinal Disorders: History, Pathophysiology, Clinical Features, and Rome IV. Gastroenterology 150, 1262–1279.e2.10.1053/j.gastro.2016.02.032Search in Google Scholar PubMed
Enck, P., Aziz, Q., Barbara, G., Farmer, A. D., Fukudo, S., Mayer, E.A., … Spiller, R.C. (2016). Irritable bowel syndrome. Nat. Rev. Dis. Primers 2, 16014.10.1038/nrdp.2016.14Search in Google Scholar PubMed PubMed Central
Fan, W., Fei, G., Li, X., Wang, X., Hu, C., Xin, H., … Fang, X. (2018). Sera with anti-enteric neuronal antibodies from patients with irritable bowel syndrome promote apoptosis in myenteric neurons of guinea pigs and human SH-Sy5Y cells. Neurogastroenterol. Motil. 30, e13457.10.1111/nmo.13457Search in Google Scholar PubMed
Friedland, R.P., and Chapman, M.R. (2017). The role of microbial amyloid in neurodegeneration. PLoS Pathog. 13, e1006654.10.1371/journal.ppat.1006654Search in Google Scholar PubMed PubMed Central
Furness, J.B. (2006). The enteric nervous system (Malden, Mass: Blackwell Pub).10.1002/9780470988756Search in Google Scholar
Furness, J.B., and Stebbing, M.J. (2018). The first brain: Species comparisons and evolutionary implications for the enteric and central nervous systems. Neurogastroenterol. Motil. 30, e13234.10.1111/nmo.13234Search in Google Scholar PubMed
Furness, J.B., Callaghan, B.P., Rivera, L.R., and Cho, H.-J. (2014). The Enteric Nervous System and Gastrointestinal Innervation: Integrated Local and Central Control. In Microbial Endocrinology: The Microbiota-Gut-Brain Axis in Health and Disease. M. Lyte and J.F. Cryan, eds. (New York, NY: Springer New York), pp. 39–71.10.1007/978-1-4939-0897-4_3Search in Google Scholar PubMed
Gecse, K., Roka, R., Ferrier, L., Leveque, M., Eutamene, H., Cartier, C., … Bueno, L. (2008). Increased faecal serine protease activity in diarrhoeic IBS patients: a colonic lumenal factor impairing colonic permeability and sensitivity. Gut 57, 591–599.10.1136/gut.2007.140210Search in Google Scholar PubMed
Grubišić, V., and Parpura, V. (2015). The second brain in autism spectrum disorder: could connexin 43 expressed in enteric glial cells play a role? Front. Cell. Neurosci. 9, 242.10.3389/fncel.2015.00242Search in Google Scholar PubMed PubMed Central
Gustafson, E., Larsson, T., and Danielson, J. (2019). Controlled outcome of Hirschsprung’s disease beyond adolescence: a single center experience. Pediatr. Surg. Int. 35, 181–185.10.1007/s00383-018-4391-5Search in Google Scholar PubMed PubMed Central
Hirschsprung, H. (1888). Stuhlträgheit Neugeborener in Folge von Dilatation und Hypertrophie des Colons. In Jahrbuch Für Kinderheilkunde Und Physische Erziehung, 27: pp. 1–7.Search in Google Scholar
Hsiao, E.Y., McBride, S.W., Hsien, S., Sharon, G., Hyde, E.R., McCue, T., … Mazmanian S.K. (2013). Microbiota Modulate Behavioral and Physiological Abnormalities Associated with Neurodevelopmental Disorders. Cell 155, 1451–1463.10.1016/j.cell.2013.11.024Search in Google Scholar PubMed PubMed Central
Hui, K.Y., Fernandez-Hernandez, H., Hu, J., Schaffner, A., Pankratz, N., Hsu, N.-Y., … Peter, I. (2018). Functional variants in the LRRK2 gene confer shared effects on risk for Crohn’s disease and Parkinson’s disease. Sci. Transl. Med. 10, eaai7795.10.1126/scitranslmed.aai7795Search in Google Scholar PubMed PubMed Central
Huizinga, J.D., and Lammers, W.J.E.P. (2009). Gut peristalsis is governed by a multitude of cooperating mechanisms. Am. J. Physiol. Gastrointest. Liver Physiol. 296, G1–G8.10.1152/ajpgi.90380.2008Search in Google Scholar PubMed
Israelyan, N., and Margolis, K.G. (2019). Reprint of: Serotonin as a link between the gut-brain-microbiome axis in autism spectrum disorders. Pharmacol. Res. 140, 115–120.10.1016/j.phrs.2018.12.023Search in Google Scholar PubMed PubMed Central
Jahng, J., and Kim, Y.S. (2016). Irritable Bowel Syndrome: Is It Really a Functional Disorder? A New Perspective on Alteration of Enteric Nervous System. Neurogastroenterol. Motil. 22, 163–165.10.5056/jnm16043Search in Google Scholar PubMed PubMed Central
Kugler, E.M., Michel, K., Zeller, F., Demir, I. E., Ceyhan, G.O., Schemann, M., and Mazzuoli-Weber, G. (2015). Mechanical stress activates neurites and somata of myenteric neurons. Front. Cell. Neurosci. 9, 342.10.3389/fncel.2015.00342Search in Google Scholar PubMed PubMed Central
Latorre, R., Sternini, C., De Giorgio, R., and Greenwood-Van Meerveld, B. (2016). Enteroendocrine cells: a review of their role in brain-gut communication. Neurogastroenterol. Motil. 28, 620–630.10.1111/nmo.12754Search in Google Scholar PubMed PubMed Central
Levinthal, D.J., Rahman, A., Nusrat, S., O’Leary, M., Heyman, R., and Bielefeldt, K. (2013). Adding to the Burden: Gastrointestinal Symptoms and Syndromes in Multiple Sclerosis. Mult. Scler. Int. 2013, 1–9.10.1155/2013/319201Search in Google Scholar PubMed PubMed Central
Li, Q., Michel, K., Annahazi, A., Demir, I. E., Ceyhan, G.O., Zeller, F., … Schemann, M. (2016a). Anti-Hu antibodies activate enteric and sensory neurons. Sci. Rep. 6, 38216.10.1038/srep38216Search in Google Scholar PubMed PubMed Central
Li, S., Fei, G., Fang, X., Yang, X., Sun, X., Qian, J., Wood, J.D., and Ke, M. (2016b). Changes in Enteric Neurons of Small Intestine in a Rat Model of Irritable Bowel Syndrome with Diarrhea. Neurogastroenterol. Motil. 22, 310–320.10.5056/jnm15082Search in Google Scholar PubMed PubMed Central
Lionnet, A., Leclair-Visonneau, L., Neunlist, M., Murayama, S., Takao, M., Adler, C.H., Derkinderen, P., and Beach, T.G. (2018). Does Parkinson’s disease start in the gut? Acta Neuropathol. 135, 1–12.10.1007/s00401-017-1777-8Search in Google Scholar PubMed
Liu, M., Seino, S., and Kirchgessner, A.L. (1999). Identification and characterization of glucoresponsive neurons in the enteric nervous system. J. Neurosci. 19, 10305–10317.10.1523/JNEUROSCI.19-23-10305.1999Search in Google Scholar
Liu, V., Dietrich, A., Kasparek, M.S., Benhaqi, P., Schneider, M.R., Schemann, M., Seeliger, H., and Kreis, M. E. (2015). Extrinsic intestinal denervation modulates tumor development in the small intestine of ApcMin/+ mice. J. Exp. Clin. Cancer Res. 34, 39.10.1186/s13046-015-0159-0Search in Google Scholar PubMed PubMed Central
Lüderitz, C. (1890). Experimentelle Untersuchungen über die Entstehung der Darmperistaltik. Archiv f. pathol. Anat. 122, 1–28.10.1007/BF01922779Search in Google Scholar
Margolis, K.G., Li, Z., Stevanovic, K., Saurman, V., Israelyan, N., Anderson, G.M., … Gershon, M.D. (2016). Serotonin transporter variant drives preventable gastrointestinal abnormalities in development and function. J. Clin. Invest. 126, 2221–2235.10.1172/JCI84877Search in Google Scholar PubMed PubMed Central
Mayer, E.A. (2011). Gut feelings: the emerging biology of gut–brain communication. Nat. Rev. Neurosci. 12, 453–466.10.1038/nrn3071Search in Google Scholar PubMed PubMed Central
Mazzuoli-Weber, G., and Schemann, M. (2015). Mechanosensitivity in the enteric nervous system. Front. Cell. Neurosci. 9, 408.10.3389/fncel.2015.00408Search in Google Scholar PubMed PubMed Central
Moloney, R.D., O’Mahony, S.M., Dinan, T.G., and Cryan, J.F. (2015). Stress-Induced Visceral Pain: Toward Animal Models of Irritable-Bowel Syndrome and Associated Comorbidities. Front. Psychiatry 6, 15.10.3389/fpsyt.2015.00015Search in Google Scholar PubMed PubMed Central
Neunlist, M., Michel, K., Reiche, D., Dobreva, G., Huber, K., and Schemann, M. (2001). Glycine activates myenteric neurones in adult guinea-pigs. J. Physiol. 536, 727–739.10.1111/j.1469-7793.2001.00727.xSearch in Google Scholar PubMed PubMed Central
Ostertag, D., Buhner, S., Michel, K., Pehl, C., Kurjak, M., Götzberger, M., … Schemann, M. (2015). Reduced Responses of Submucous Neurons from Irritable Bowel Syndrome Patients to a Cocktail Containing Histamine, Serotonin, TNFα, and Tryptase (IBS-Cocktail). Front. in Neurosci. 9, 465.10.3389/fnins.2015.00465Search in Google Scholar PubMed PubMed Central
O’Sullivan, M., Clayton, N., Breslin, N.P., Harman, I., Bountra, C., McLaren, A., and O’Morain, C.A. (2000). Increased mast cells in the irritable bowel syndrome. Neurogastroenterol. Motil. 12, 449–457.10.1046/j.1365-2982.2000.00221.xSearch in Google Scholar PubMed
Pan, W.K., Zheng, B.J., Gao, Y., Qin, H., and Liu, Y. (2011). Transplantation of Neonatal Gut Neural Crest Progenitors Reconstructs Ganglionic Function in Benzalkonium Chloride-Treated Homogenic Rat Colon. J. Surg. Res. 167, e221–e230.10.1016/j.jss.2011.01.016Search in Google Scholar PubMed
Ro, S., Hwang, S.J., Muto, M., Jewett, W.K., and Spencer, N.J. (2006). Anatomic modifications in the enteric nervous system of piebald mice and physiological consequences to colonic motor activity. Am. J. Physiol. Gastrointest. Liver Physiol. 290, G710–718.10.1152/ajpgi.00420.2005Search in Google Scholar PubMed
Róka, R., Rosztóczy, A., Leveque, M., Izbéki, F., Nagy, F., Molnár, T., … Bueno, L. (2007). A Pilot Study of Fecal Serine-Protease Activity: A Pathophysiologic Factor in Diarrhea-Predominant Irritable Bowel Syndrome. Clin. Gastroenterol. Hepatol. 5, 550–555.10.1016/j.cgh.2006.12.004Search in Google Scholar PubMed
Sampson, T.R., Debelius, J.W., Thron, T., Janssen, S., Shastri, G.G., Ilhan, Z.E., … Mazmanian, S.K. (2016). Gut Microbiota Regulate Motor Deficits and Neuroinflammation in a Model of Parkinson’s Disease. Cell 167, 1469–1480.e12.10.1016/j.cell.2016.11.018Search in Google Scholar PubMed PubMed Central
Schemann, M., and Grundy, D. (1992). Electrophysiological identification of vagally innervated enteric neurons in guinea pig stomach. Am. J. Physiol. Gastrointest. Liver Physiol. 263, G709–G718.10.1152/ajpgi.1992.263.5.G709Search in Google Scholar PubMed
Schemann, M., Frieling, T., and Enck, P. (2019). To learn, to remember, to forget—How smart is the gut? Acta Physiol. e13296.10.1111/apha.13296Search in Google Scholar PubMed PubMed Central
Sergi, C. (2015). Hirschsprung’s disease: Historical notes and pathological diagnosis on the occasion of the 100th anniversary of Dr. Harald Hirschsprung’s death. World J. Clin. Pediatr. 4, 120.10.5409/wjcp.v4.i4.120Search in Google Scholar PubMed PubMed Central
Shimizu, H., Koizumi, O., and Fujisawa, T. (2004). Three digestive movements in Hydra regulated by the diffuse nerve net in the body column. J. Comp. Physiol. A 190.10.1007/s00359-004-0518-3Search in Google Scholar PubMed
Smith, T.K., Spencer, N.J., Hennig, G.W., and Dickson, E.J. (2007). Recent advances in enteric neurobiology: mechanosensitive interneurons. Neurogastroenterol. Motil. 19, 869–878.10.1111/j.1365-2982.2007.01019.xSearch in Google Scholar PubMed
Spear, E.T., Holt, E.A., Joyce, E.J., Haag, M.M., Mawe, S.M., Hennig, G.W., Lavoie, B., Applebee, A.M., Teuscher, C., and Mawe, G.M. (2018). Altered gastrointestinal motility involving autoantibodies in the experimental autoimmune encephalomyelitis model of multiple sclerosis. Neurogastroenterol. Motil. 30, e13349.10.1111/nmo.13349Search in Google Scholar PubMed PubMed Central
Stamp, L.A. (2017). Cell therapy for GI motility disorders: comparison of cell sources and proposed steps for treating Hirschsprung disease. Am. J. Physiol. Gastrointest. Liver Physiol. 312, G348–G354.10.1152/ajpgi.00018.2017Search in Google Scholar PubMed
Stamp, L.A., Gwynne, R.M., Foong, J.P.P., Lomax, A.E., Hao, M.M., Kaplan, D. I., … Young, H.M. (2017). Optogenetic Demonstration of Functional Innervation of Mouse Colon by Neurons Derived From Transplanted Neural Cells. Gastroenterology 152, 1407–1418.10.1053/j.gastro.2017.01.005Search in Google Scholar PubMed
Svensson, E., Horváth-Puhó, E., Thomsen, R.W., Djurhuus, J.C., Pedersen, L., Borghammer, P., and Sørensen, H.T. (2015). Vagotomy and subsequent risk of Parkinson’s disease: Vagotomy and Risk of PD. Ann. Neurol. 78, 522–529.10.1002/ana.24448Search in Google Scholar PubMed
Tittel, K. (1901). Ueber eine angeborene Missbildung des Dickdarmes. Wien. klin. Wchnschr. 903–907.Search in Google Scholar
Traini, C., Evangelista, S., Girod, V., Faussone-Pellegrini, M.S., and Vannucchi, M.G. (2016). Changes of excitatory and inhibitory neurotransmitters in the colon of rats underwent to the wrap partial restraint stress. Neurogastroenterol. Motil. 28, 1172–1185.10.1111/nmo.12816Search in Google Scholar PubMed
Wang, W. (2018). Optogenetic manipulation of ENS – The brain in the gut. Life Sci. 192, 18–25.10.1016/j.lfs.2017.11.010Search in Google Scholar PubMed
Wood, J.D., Liu, S., Drossman, D.A., Ringel, Y., and Whitehead, W.E. (2012). Anti-Enteric Neuronal Antibodies and the Irritable Bowel Syndrome. Neurogastroenterol. Motil. 18, 78–85.10.5056/jnm.2012.18.1.78Search in Google Scholar PubMed PubMed Central
Wunsch, M., Jabari, S., Voussen, B., Enders, M., Srinivasan, S., Cossais, F., … Kuerten, S. (2017). The enteric nervous system is a potential autoimmune target in multiple sclerosis. Acta Neuropathol. 134, 281–295.10.1007/s00401-017-1742-6Search in Google Scholar PubMed
Zhang, L., Song, J., and Hou, X. (2016). Mast Cells and Irritable Bowel Syndrome: From the Bench to the Bedside. Neurogastroenterol. Motil. 22, 181–192.10.5056/jnm15137Search in Google Scholar PubMed PubMed Central
Legend to Movie
The movie shows the spike pattern in enteric neurons related to muscle movement.
The top image shows a myenteric ganglion labeled with a voltage sensitive dye. Individual neurons are seen as black ring-like structures because the dye incorporates into their outer membrane. The trace at the bottom shows the spike discharge in one of the neurons, which is indicated by the grey frame. The red dot moving over the trace corresponds to the false color-coded activity level in the movie (red is the peak of the spike). Each spike is coded as a red color. The recording period is several seconds and the spike burst corresponds to the frequency of intestinal contractions. The activity within the ganglion very likely reflects activity in muscle motor neurons.
© 2020 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Review Article
- Food reward and gut-brain signalling
- Synapses: Multitasking Global Players in the Brain
- Apolipoprotein E: Cholesterol metabolism and Alzheimer’s pathology
- The enteric nervous system: “A little brain in the gut”
- The function of lysosomes and their role in Parkinson’s disease
- Rezension
- Konrad Lehmann: Das schöpferische Gehirn Auf der Suche nach der Kreativität – eine Fahndung in sieben Tagen
- Obituary
- Prof. Dr. med. Dr. med. h.c. Georg W. Kreutzberg
- Nachrichten der Gesellschaft
- EU-Projektförderung 2020 Neurowissenschaften
Articles in the same Issue
- Titelseiten
- Review Article
- Food reward and gut-brain signalling
- Synapses: Multitasking Global Players in the Brain
- Apolipoprotein E: Cholesterol metabolism and Alzheimer’s pathology
- The enteric nervous system: “A little brain in the gut”
- The function of lysosomes and their role in Parkinson’s disease
- Rezension
- Konrad Lehmann: Das schöpferische Gehirn Auf der Suche nach der Kreativität – eine Fahndung in sieben Tagen
- Obituary
- Prof. Dr. med. Dr. med. h.c. Georg W. Kreutzberg
- Nachrichten der Gesellschaft
- EU-Projektförderung 2020 Neurowissenschaften