A biochemical view on the septins, a less known component of the cytoskeleton
-
Benjamin Grupp
Benjamin Grupp graduated in biochemistry and is currently PhD student at the University of Ulm/Germany. He applies
in vitro andin silico approaches to study biochemical features of yeast septins.Thomas Gronemeyer studied biochemistry in Bochum/Germany and obtained a PhD in protein engineering from the Federal Institute of Technology in Lausanne/Switzerland. Today he is permanent staff scientist and lecturer for molecular- and cell biology at the University of Ulm. His research focus lies on the biochemistry of yeast and mammalian septins.



Abstract
The septins are a conserved family of guanine nucleotide binding proteins, often named the fourth component of the cytoskeleton. They self-assemble into non-polar filaments and further into higher ordered structures. Properly assembled septin structures are required for a wide range of indispensable intracellular processes such as cytokinesis, vesicular transport, polarity establishment and cellular adhesion. Septins belong structurally to the P-Loop NTPases. However, unlike the small GTPases like Ras, septins do not mediate signals to effectors through GTP binding and hydrolysis. The role of nucleotide binding and subsequent GTP hydrolysis by the septins is rather controversially debated. We compile here the structural features from the existing septin crystal- and cryo-EM structures regarding protofilament formation, inter-subunit interface architecture and nucleotide binding and hydrolysis. These findings are supplemented with a summary of available biochemical studies providing information regarding nucleotide binding and hydrolysis of fungal and mammalian septins.
Introduction
In 1970/71 Leland H. Hartwell discovered four alleles in Saccharomyces cerevisiae causing a temperature-sensitive cytokinesis defect. He named the corresponding genes CDC3, CDC10, CDC11 and CDC12 after their role in the cell division cycle (Hartwell 1971). Later the protein family these proteins belong to was named “septins” in the lab of J. R. Pringle because of their localization as rings to the septate bud neck in S. cerevisiae (Sanders and Field 1994). In addition to the four initially identified septin subunits, another mitotic septin Shs1 (seventh homolog of septin) and the sporulation specific septins Spr3 and Spr28 (sporulation regulated) were discovered in (De Virgilio et al. 1996; Fares et al. 1996; Mino et al. 1998). In the following decades, septins could be identified in all eukaryotes except of higher plants and they seem to be structurally highly conserved meanwhile the number of septin genes per organism is variable (e.g. 7 in S. cerevisiae, each 13 in Homo sapiens and Mus musculus, 2 in Caenorhabditis elegans, 1 in Chlamydomonas) (Pan et al. 2007; Shuman and Momany 2022). Their molecular mass ranges from 30–65 kDa and the individual subunits arrange into linear, apolar protofilaments termed rods and subsequently into higher ordered structures such as linear filaments, gauzes or rings. The arrangement of the mammalian septins into the canonical hetero-octameric protofilaments shows the order SEPT2-6-7-3-3-6-7-2 whereas S. cerevisiae canonical hetero-octamers follow the order Cdc11-12-3-10-10-3-12-11 (Bertin et al. 2008; Soroor et al. 2020) (Figure 1).
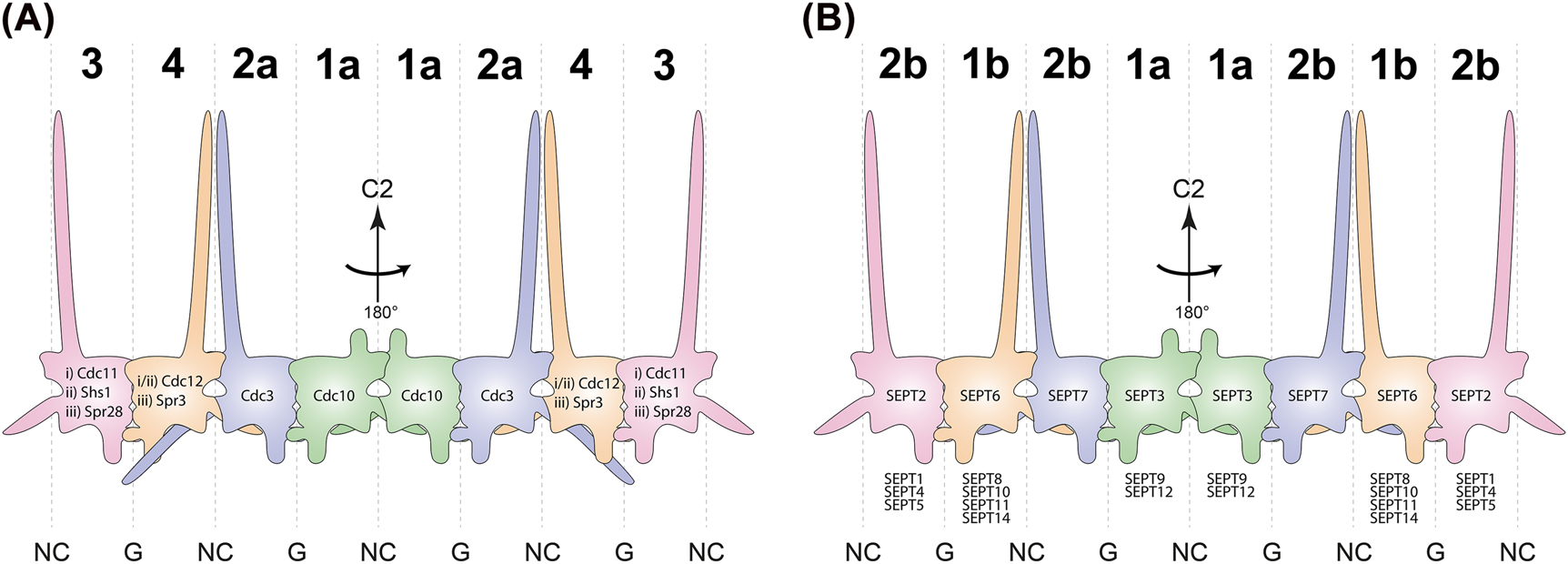
Composition of yeast- and human septin protofilaments. (A) Schematic representation of S. cerevisiae septin octamers. The protein identities are outlined within the respective subunit. The two central subunits are constant within all possible rod populations i–iii while the two terminal subunits determine the identity of the respective population (i: Cdc11 containing mitotic, ii: Shs1 containing mitotic, iii: meiotic). (B) Schematic representation of human septin octamers. The eponymous subunit of the respective Kinoshita group is indicated within the respective subunit, other members of the respective group are listed below. All protofilament species in A and B exhibit a cyclic symmetry (C2) around the central NC interface. Capital numbers on top determine the corresponding Momany group. Interfaces are indicated on bottom.
A common nomenclature for mammalian septins was only established in 2002 (Macara et al. 2002). Subsequently M. Kinoshita sorted the 13 human septins into four groups based on sequence homology which he named after their best-studied members: SEPT2 (including SEPT1, SEPT4 and SEPT5), SEPT3 (including SEPT9 and SEPT12), SEPT6 (including SEPT8, SEPT10, SEPT11 and SEPT14), and SEPT7 which is the only member of its group (Kinoshita 2003) (Figure 1). Kinoshita predicted the exchangeability of septin subunits within the septin rod against members of the same group based on molecular recognition which was later named “Kinoshita’s rule”.
The Kinoshita groups were recently updated by the Momany lab by defining new groups based on phylogenetic relationships and including also septins from other species (Shuman and Momany 2022). The septins were sorted into five orthologous groups with subgroups a and b for group 1 and 2 (Figure 1). Septins of the groups 1–4 assemble into nonpolar heteropolymers meanwhile members of group 5 are not incorporated into canonical heteropolymers as they seem to interact just transiently with the core septins. Groups 3–5 are unique to fungi (Pan et al. 2007; Hernández-Rodríguez et al. 2014; Auxier et al. 2019; Shuman and Momany 2022).
The individual septin subunits assemble into filamentous structures exhibiting a thickness of a few nm. Septins localize in vivo to the division site, the plasma membrane, the annuli of spermatozoa as well as the bases of cilia and dendrites (Menon and Gaestel 2017). In S. cerevisiae septins act mainly as scaffolds for protein recruitment and as diffusion barriers which regulate the protein distribution between mother and daughter cells (Oh and Bi 2011). They support actomyosin ring assembly as well as the establishment of polarized growth for cell division (Bi and Park 2012; Kozubowski et al. 2005; Ong et al. 2014). In humans, septins have been shown to interact with actin and microtubules and to play important roles in chromosome segregation, cytokinesis, exocytosis, oncogenesis, apoptosis and in the establishment of several diseases (Bartsch et al. 2011; Dolat et al. 2014; Estey et al. 2010; Ivanov et al. 2021; Marcus et al. 2019; Mavrakis et al. 2014; Sellin et al. 2011; Smith et al. 2015; Tokhtaeva et al. 2015). The regulation of localized septin assembly and functionality seems to be highly complex as it turned out to be dependent on the interacting septins, the expressed isoform, post-translational modifications – including phosphorylation, acetylation, ubiquitination and sumoylation – other interaction partners and maybe the guanine nucleotide binding state (Hernández-Rodríguez and Momany 2012; Johnson and Blobel 1999; Ribet et al. 2017; Shen et al. 2017; Wang et al. 2021; Weems and McMurray 2017; Werner and Yadav 2022). Moreover, recent studies have shown that some septins have the ability to detect and bind membranes with a specific curvature, most likely via amphipathic helices close to the C-terminus. The association to membranes has been shown to depend on the character of the membrane incorporated lipids (Beber et al. 2019; Woods et al. 2021).
Structural features of septins and interface architecture
The PDB database lists at the time of writing 23 entries from various human septin monomers, hetero-dimers or homo-multimers, two entries from Schistosoma septin 10, and each one from Chlamydomonas and S. cerevisiae. Besides that, five entries covering only the C-terminal coiled coils following the guanine nucleotide binding domains (G-domains) are available. Human structures comprise the subunits SEPT2, 3, 7, 9 and 12 as well as SEPT2/6, 2/3, 2/8 and 2/11 hetero-dimers (Brausemann et al. 2016; do Vale Castro et al. 2020; Mendonça et al. 2021; Rosa et al. 2020; Sirajuddin et al. 2007; Zent et al. 2011; Zeraik et al. 2014).
Only two structures from septin complexes comprising more than two subunits are available:
The first publication of a crystal structure of a septin was the human SEPT2/6/7 trimer (PDB-ID 2QAG) (Sirajuddin et al. 2007). The cryo-EM structure of a human hexamer composed of the same subunits was presented more than a decade later (PDB-ID 7M6J) (Mendonça et al. 2021).
These structures provide insight into the architecture of the inter-subunit contacts and were used for the following summary of septin structural features.
Inter-subunit contacts can be classified as two distinct interfaces: The first interface is termed G-interface as it is solely stabilized by interactions of G-domain components (see below), meanwhile the second interface is maintained by interactions between the septin specific variable N- and C-terminal extensions (NTE/CTE) flanking the G-domain. Therefore, this interface is called the NC-interface. Both interfaces appear in an alternating fashion along a septin protofilament (Mendonça et al. 2021; Sirajuddin et al. 2007) (Figure 1).
With the publication of the first septin crystal structure it was confirmed that septins are guanine nucleotide binding proteins belonging to the superclass of P-loop NTPases which was previously predicted by sequence analysis and shown by different in vitro assays (Baur et al. 2018; Farkasovsky et al. 2005; Versele and Thorner 2004; Vrabioiu et al. 2004).
They contain all necessary elements that classify active GTPases such as the switch 1 and 2 loops and the P-loop (Cavini et al. 2021; Fonseca Valadares et al. 2017). The G-domain resembles a modified Rossmann fold consisting of repetitive αβ-units which fold into a central β-sheet consisting of six β-strands (Longo et al. 2020; Weirich et al. 2008). All features highlighted in the following are illustrated in Figure 2. The central β-sheet is surrounded by seven α-helices plus the additional N-terminal α0-helix. Unique for the septins compared to other small GTPases are all segments located N-terminally to β1 and C-terminally to α4. In septins, α4 is followed by a polyacidic region (PAR) leading into the short α5′-helix, then into β6 which is followed by the septin unique element (SUE), a prominent feature of all septin G-domains. The SUE is folded into a characteristic β-meander consisting of three β-strands (SUE-βββ) and the C-terminal helices α5 and α6. Further septin-unique features in the G-domain are an elongated switch 2 loop participating in G-interface dimerization, an elongated α2-helix connecting the G- and NC-interfaces and the C-terminal loop participating in the NC-interface containing a polybasic motif (PB2). Upstream of the β1-strand septins contain the N-terminal extension (NTE) which is highly variable in length, ranging from a few residues in Cdc10 to several hundred in human SEPT9. The NTE seems to be largely unstructured except of the α0-helix which contains another polybasic motif (PB1) being involved in membrane binding (Bertin et al. 2010; Zhang et al. 1999). The C-terminal extension (CTE), located downstream the α6-helix in all septins (except of the subgroup 1a members), contains at least one heptad sequence which is predicted to form a coiled coil helix bundle within and across septin polymers. The bundles are oriented perpendicular to the septin filament axis following the directionality of the α6-helix (Finnigan et al. 2015a, 2015b; Leonardo et al. 2021).
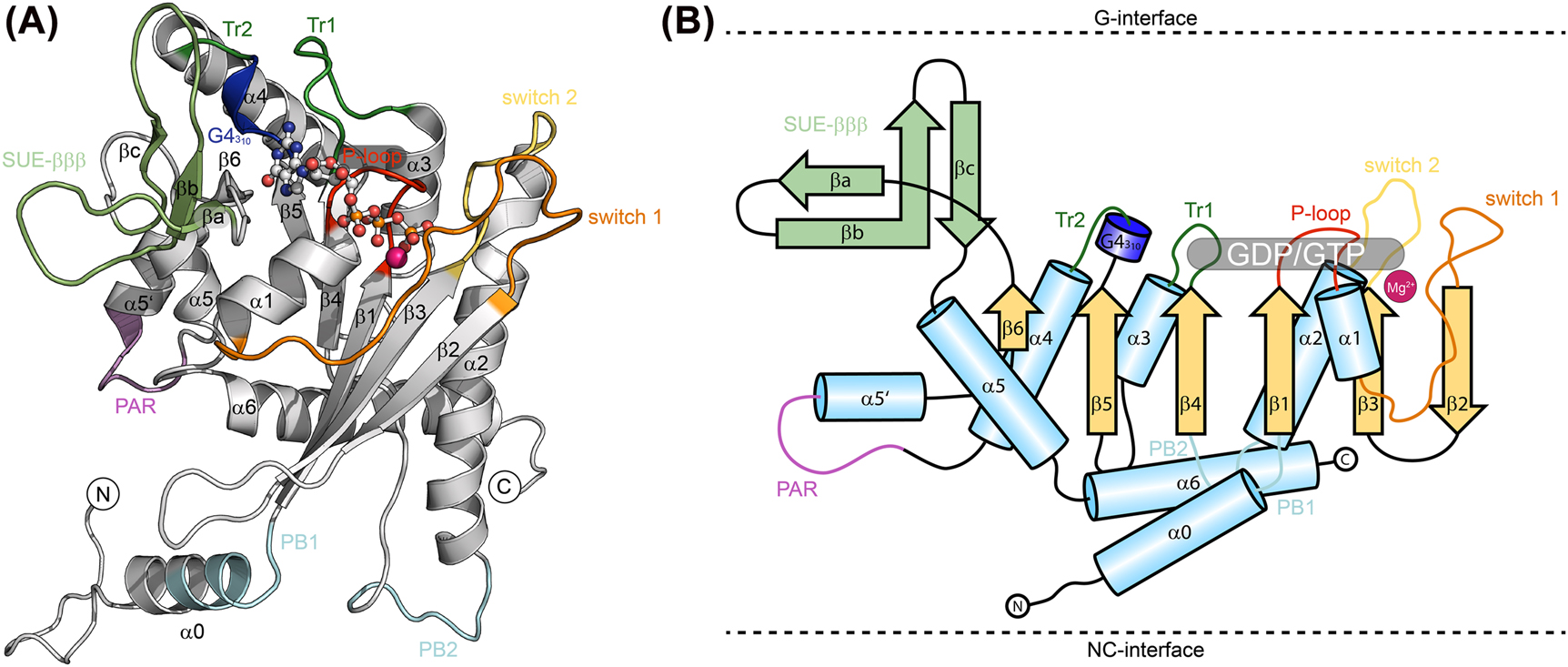
Representative crystal structure (SEPT6-GTP, extracted from PDB-ID 7M6J) (A), and schematic topology (B) of a septin subunit. Sheets and helices are numbered according to the classical G domain numbering of Ras like proteins. SUE, septin unique element; Tr, trans-loop; PB, poly-basic motif; PAR, poly-acidic region. The GTP and the Mg2+ in SEPT6 are shown as ball-and-stick presentation.
Multimerization via G- and NC-interfaces is a characteristic feature of septins. G-interfaces are stabilized by interactions between the P-loop, switch 1, switch 2, G4, trans loops 1 and 2 (Tr1 and Tr2, connecting β4 with α3 and β5 with α4, respectively), as well as the bound nucleotides (Figure 3). Furthermore, the G interface is strongly stabilized by hydrophobic interactions between the SUE-βββ of both subunits (Rosa et al. 2020; Sirajuddin et al. 2007; Zent et al. 2011). The selectivity for the preferred G-interface dimerization partner seems on one hand to be at least partly governed by the highly variable switch 1 loop and the switch 2 loop which adopt different conformations in physiological compared to non-physiological interfaces. On the other hand the selectivity might be further regulated by the bound nucleotide (Cavini et al. 2021). The only available septin crystal structure not having a bound guanine nucleotide is the Cdc11 apo protein monomer which is not forming a G- and/or NC-interface indicating that the nucleotide might be an integral part of the G-interface (Brausemann et al. 2016).
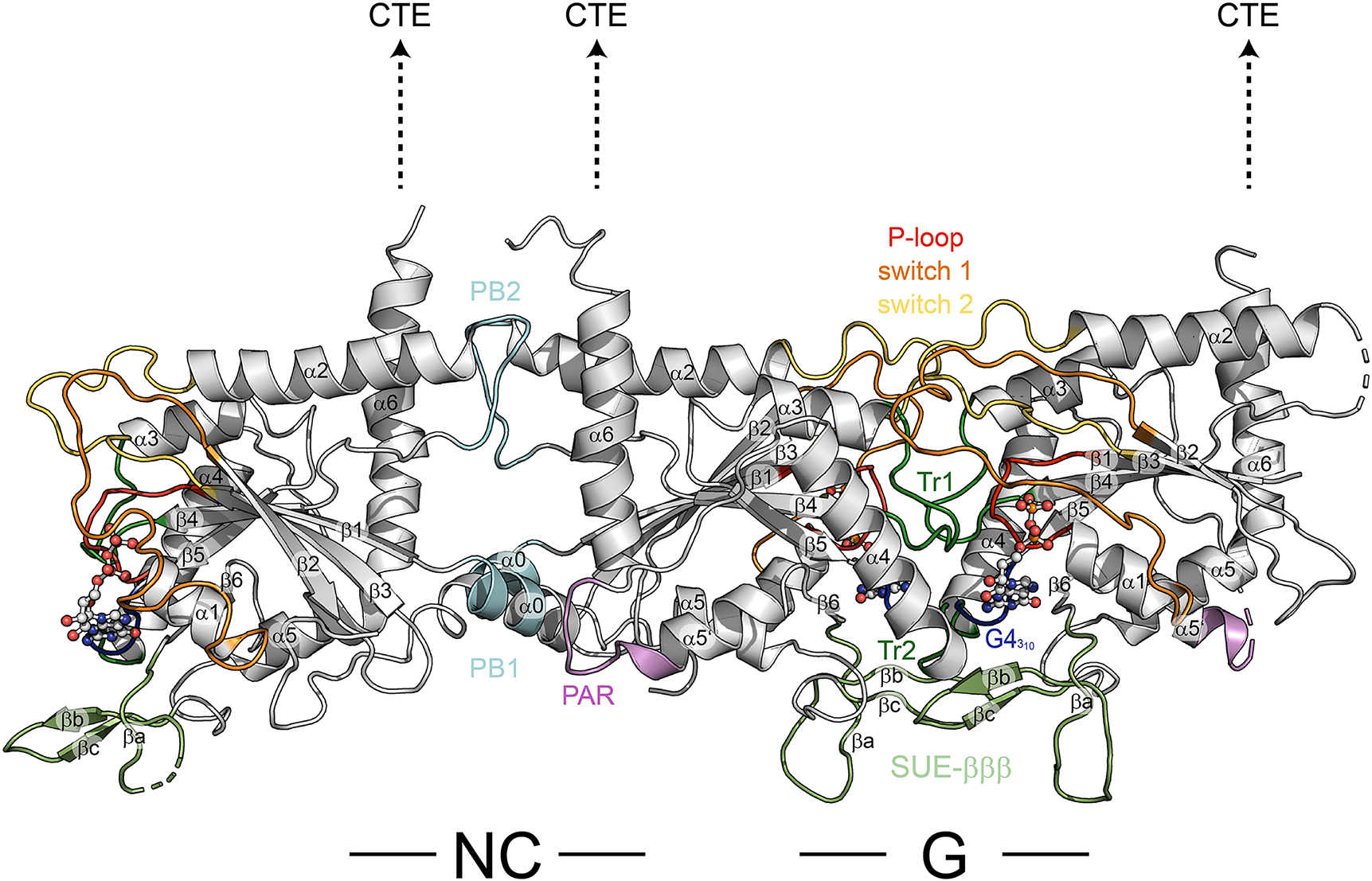
Trimer comprised of the human septins (from left to right) SEPT7-SEPT6-SEPT2 (extracted from PDB-ID 7M6J) showing an NC interface between SEPT7 and SEPT6 and a G interface between SEPT6 and SEPT2. The positions of the interfaces are indicated. The dashed arrows indicate the expected positions of the C-terminal extensions (CTE). Classical features of the G domain of Ras like proteins (switches 1 and 2, P-loop) and septin unique features are annotated; numbering of helices and sheets as in Figure 2. SUE, septin unique element; Tr, trans-loop; PB, poly-basic motif; PAR, poly-acidic region.
The NC-interface on the opposite of the G-domain is mainly stabilized by ionic interactions between charged amino acids of the α6-helix, the C-terminal region of the α2-helix with its helical dipole moment and PB2 (Figure 3). Moreover, it was suggested that the α0-helix and its preceding hook-loop are important for NC-interface stabilization (Cavini et al. 2021). The α0-helix interacts through its polybasic motif (PB1) with the PAR in the neighboring subunit (Figure 4), however, this interaction is not confirmed in all available septin crystal structures either because the PAR could not be resolved (PDB-ID 2QA5, 2QAG) or the α0-helix itself is sometimes not incorporated in the NC-interface (PDB-ID 4Z54) (do Vale Castro et al. 2020).
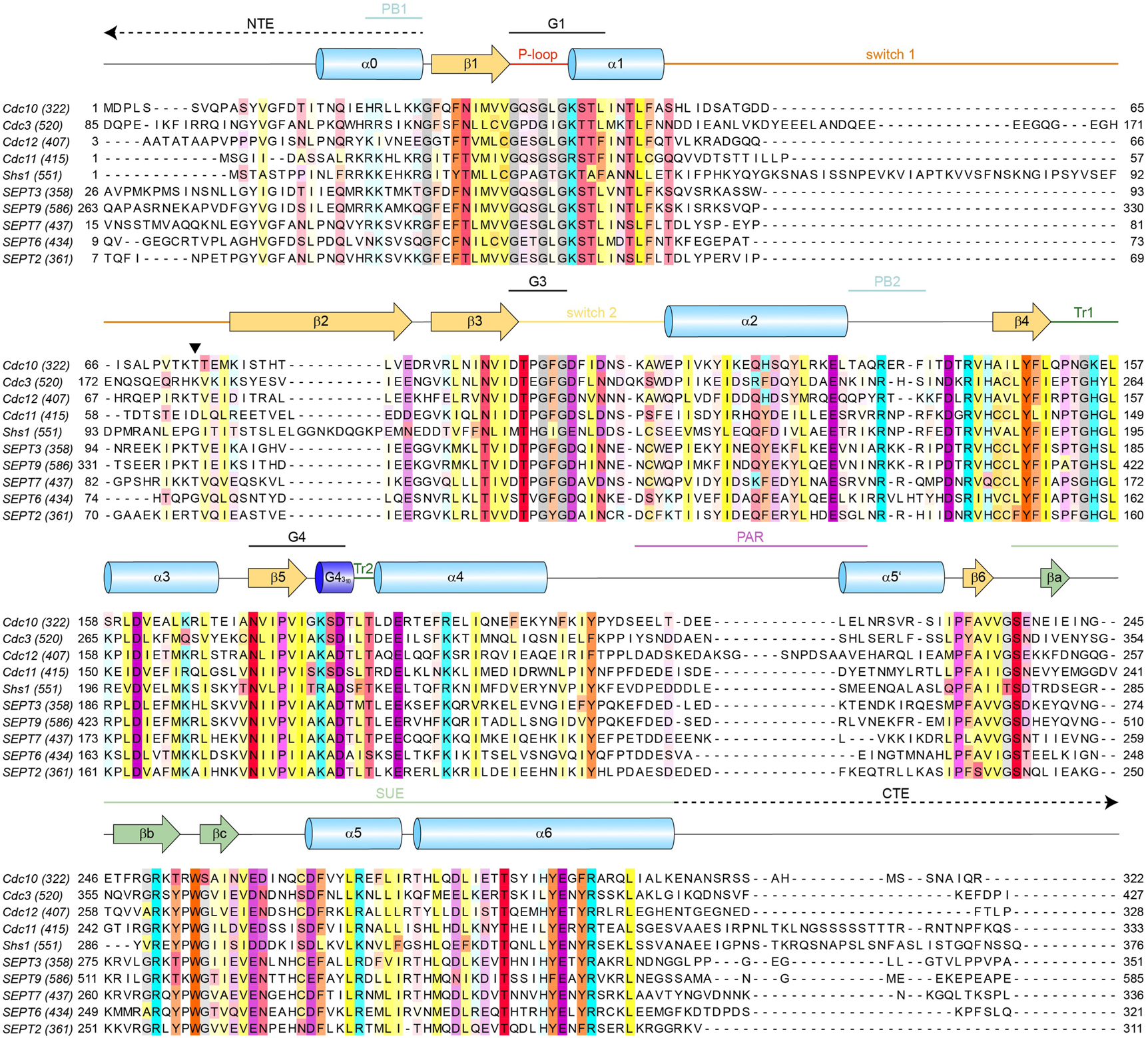
Sequence alignment of the mitotic septins from S. cerevisiae and selected human septins with prominent structural features annotated. SUE, septin unique element; Tr, trans-loop; PB, poly-basic motif; PAR, poly-acidic region. NTE, N-terminal extension; CTE, C-terminal extension. Switches 1 and 2, the P-loop and G1-4 motifs are the classical structural motifs found in all small GTPases. The black triangle marks the position of the catalytically active Thr(Sw1). Positions showing a conservation of ≥30% are colored according to their biochemical properties with the color intensity increasing with the degree of conservation. Purple – acidic; cyan – basic (including His); yellow – hydrophobic; orange – aromatic; red – polar; pink – proline; grey – glycine; gold – cysteine. The total amino acid number is provided in brackets behind the respective subunit identifier. For human septins always isoform 1 was used.
Septin protofilament formation
S. cerevisiae and mammalian septins assemble into either hetero-octameric or hetero-hexameric rod-like protofilaments (Bertin et al. 2008; Fischer et al. 2022; Garcia et al. 2011; Iv et al. 2021; Khan et al. 2018). Protofilament formation was shown to be an important prerequisite for association with some binding partners as new topologies are formed due to septin-septin multimerization (Sheffield et al. 2003). The protofilaments are nonpolar as they exhibit a cyclic symmetry (C2) around the homotypic interface formed between the central rod subunits (Figure 1).
Septin protofilament assembly appears to be a fast process in vivo which prevents the detection of any intermediates in wild type cells (Garcia et al. 2016; Johnson et al. 2015; Weems and McMurray 2017; Ziman et al. 1991). In 2017 Weems et al. deciphered the step-wise pathway of septin protofilament assembly in S. cerevisiae using a Bimolecular Fluorescence Complementation assay (Weems and McMurray 2017). In addition to revealing the order of septin interactions they predicted active communication between G- and NC-interfaces mediated by either the nucleotide binding state or the association of the native septin binding partner. In their model the Cdc11- and Shs1-rods form by homodimerization of Cdc10 which occurs before the interaction with the other septin subunits. During formation of Cdc11 containing rods, Cdc12 first associates with Cdc11 in a G-interface. This dimer then recruits Cdc3 to the NC-interface in Cdc12 inducing a conformational change in the long NTE of Cdc3 which seems to autoinhibit G-interface association with Cdc10. The Cdc11-rod protofilament assembly is finalized by both sided G-interface association of Cdc11-Cdc12-Cdc3 trimers with the central Cdc10-Cdc10 dimer. Furthermore, Weems et al. investigated the influence of bound guanine nucleotides on this assembly pathway by mutational analysis. However, the guanine nucleotide binding and hydrolysis mutations were introduced based on sequence homologies which makes the proposed role of guanine nucleotide binding still a matter of debate as the influence of these mutations was not further validated by in vitro experiments. Nevertheless, several studies indicate that nucleotide binding (and, for some septins, hydrolysis) is an early, one-time event in the septin life cycle that allows the adoption of conformations competent for septin protofilament assembly (Nagaraj et al. 2008; Sirajuddin et al. 2009; Weems et al. 2014; Zent and Wittinghofer 2014).
Septin (proto-)filaments can be produced in vitro by expressing the respective subunits from compatible bicistronic plasmids in Escherichia coli. Recombinantly expressed septin rods from yeast and human appear as hexamers or octamers under the high salt conditions (300 mM salt and up) usually applied in common purification workflows (Bertin et al. 2008; Renz et al. 2013). Filament formation can be induced by dialysis of the septin preparation in low salt buffer (below 100 mM). End-to-end filament formation by lowering the ionic strength of the surrounding medium can be explained by the structural findings described above which imply that NC interfaces are stabilized by ionic interactions between charged amino acids of the neighboring subunits. High ionic strength in the surrounding medium would screen the charges of the amino acids in the exposed terminal interface of the rods, thereby impeding end-to-end joining into a functional NC interface. Whereas human septin filaments form readily from both hexameric and octameric protofilaments (Fischer et al. 2022; Iv et al. 2021; Sheffield et al. 2003), contradictory results concerning yeast septin filaments are published. The majority of the available studies indicate that Cdc11-less septin preparations do not form filaments (Bertin et al., 2008; Garcia et al. 2011; Versele et al. 2004), however, Farkasovski et al. show filament formation of both Cdc3-Cdc12-Cdc10 and Cdc3-Cdc11-Cdc12 containing septin preparations at 100 mM NaCl (Farkasovsky et al. 2005) and in another study Cdc11-less filaments were produced on lipid monolayers (Bertin et al. 2010). In summary, the sequential order of septin subunit assembly into octameric rods could be elucidated for S. cerevisiae septins but the knowledge regarding the role of nucleotide binding and hydrolysis in septin (proto-)filament formation is still incomplete.
Structural basis for nucleotide binding and hydrolysis
Septins contain the conserved GTPase motifs G1 (in P-loop/Walker-A motif), G3 (in switch 2) and G4. Switch 1 and switch 2 are named after their large conformational changes committed during nucleotide hydrolysis (Vetter and Wittinghofer 2001) which has been also described at least for some septins (see below). Both loops have been observed to be structured and in contact with the nucleotide in the GTP-bound state in some septin subunits meanwhile the loss of the γ-phosphate in the GDP-bound state breaks the contacts making the loops highly disordered (Cavini et al. 2021; Sirajuddin et al. 2009; Vetter and Wittinghofer 2001). The G2 and G5 regions observed in other small GTPases are not found in septins (Cavini et al. 2021) (Figures 2 and 3).
In the following, aspects regarding the presumable GTP hydrolysis by septins are discussed in detail. A sequence alignment of the five mitotic yeast septins and the septins of the canonical human octamer (plus SEPT3) is shown in Figure 4, providing an overview of the involved structural elements and amino acid residues.
The most important amino acids for guanine nucleotide binding in small GTPases are present in all septins. These are the glycine residues in the P-loop which enable wrapping of the loop around the phosphate groups and they further interact directly with the β- and γ-phosphate via hydrogen bonding. Moreover, all septins except Cdc11 from yeast contain the conserved lysine residue in the P-loop. Cdc11 has an arginine in this position. The basic residue bridges the acidic β- and γ-phosphate groups and has a strong effect on the guanine nucleotide affinity (Sirajuddin et al. 2007). Furthermore, the serine side chain (sometimes replaced by a threonine) acts as Lewis base in a chelate complex formed with Mg2+ (if present) which is a prerequisite for catalytic activity or guanine nucleotide binding in several small GTPases (Rudack et al. 2012; Zhang et al. 2000) (Figure 5). The conserved glycine Gly(Sw2) (following the newly established nomenclature “residue(motif)” (Cavini et al. 2021)) in G3 interacts with the γ-phosphate of GTP via a hydrogen bond of its backbone nitrogen atom and the Asp(Sw2) binds a water molecule via a hydrogen bond which itself coordinates Mg2+ (Cavini et al. 2021) (Figure 5). The conserved Asp(G4) from the G4 motif connects to the guanine ring via a bifurcated hydrogen bond thereby creating specificity for binding of guanine nucleotides (Vetter and Wittinghofer 2001). Interestingly, not all septins contain the characteristic threonine residue Thr(Sw1) in the switch 1 (Figure 3, black triangle) which interacts with the γ-phosphate of bound GTP via its backbone nitrogen and coordinates Mg2+ with the side chain hydroxyl group. In septins Thr(Sw1) has been shown to increase the affinity for guanine nucleotides and to be important for hydrolytic activity as it is also involved in orienting the attacking water molecule with its backbone oxygen atom (Sirajuddin et al., 2009). Mutating Thr(Sw1) against alanine in the catalytically active yeast septins Cdc10 (Thr74) and Cdc12 (Thr75) results in temperature-sensitive mutants (Sirajuddin et al. 2009; Versele and Thorner 2004). Septins lacking the threonine belong all to the Momany groups 1b, 2a and 3 (Shuman and Momany 2022). The glutamine residue in switch 2 of many Ras-like small GTPases (Gln61 in Ras) which has been shown to be responsible for proper orientation of the attacking water molecule is replaced by another amino acid in septins (in most of the cases phenylalanine or tyrosine) (Calixto et al. 2019; Carvalho et al. 2015; Shuman and Momany 2022). Additionally, a conserved arginine residue Arg(βb) in the βb-strand and a conserved glycine residue Gly(“G5”) in a remnant G5 motif present in the loop connecting β6 and βa are in contact with the guanine ring. Arg(βb) as well as Lys(G4) form a classic π-cation stack sandwiching the guanine ring from both sides (Cavini et al. 2021). From the neighboring subunit usually a conserved glutamate residue Glu(Tr2) located at the N-terminus of α4 is in close proximity to the guanine nucleotide ring and forms a hydrogen bond to the ribose. Moreover, a histidine His(Tr1) (or less frequently a basic amino acid) of the neighboring subunit is in close proximity to the β- and γ-phosphate. His(Tr1) is located in the septin-specific loop (trans loop 1) connecting β4 and α3 (Cavini et al. 2021). Exclusively in algal septins the position of His(Tr1) is occupied by an arginine residue resembling an arginine finger which accelerates GTP hydrolysis (Pinto et al. 2017).
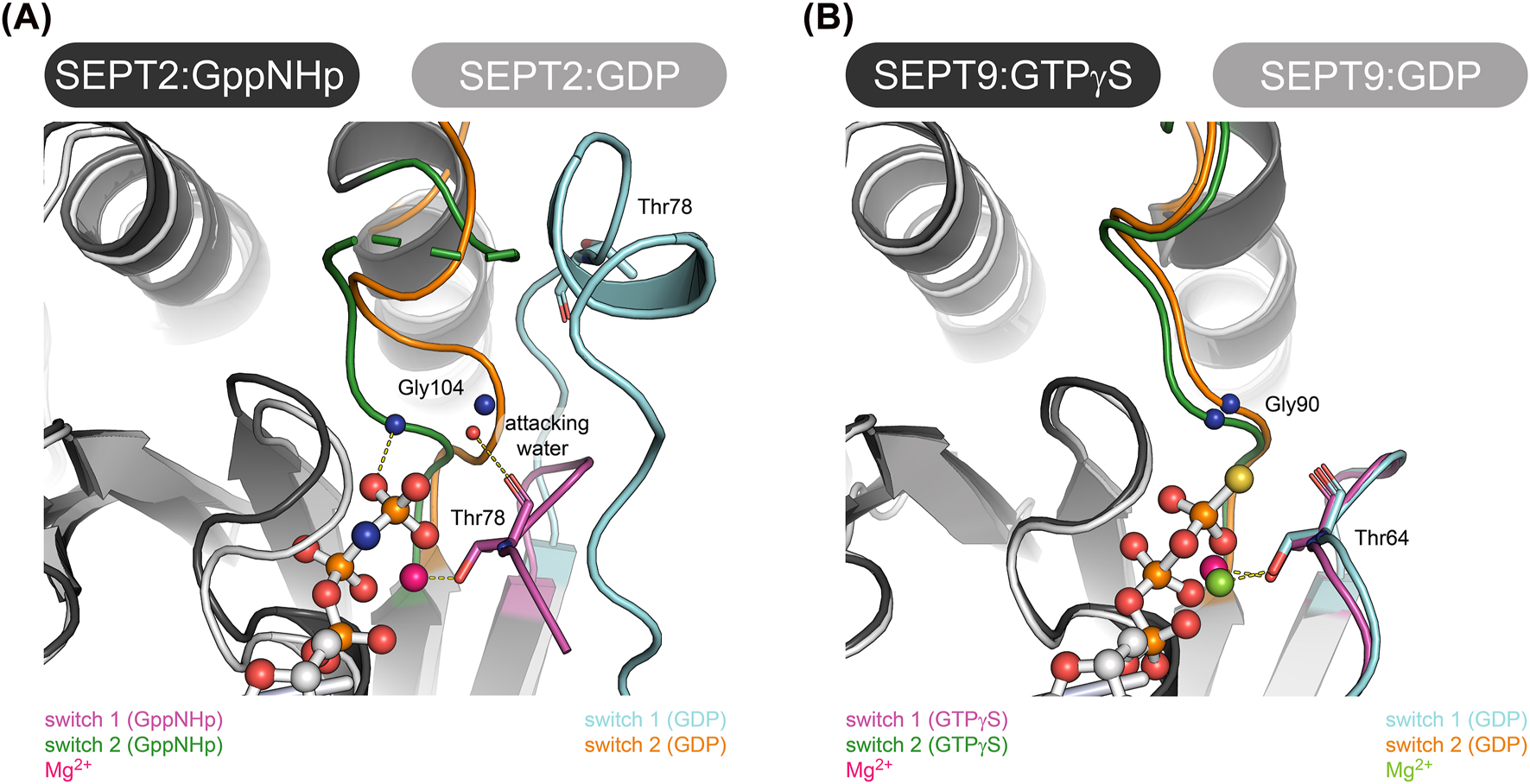
Structural features of the nucleotide binding pocket in SEPT2 and SEPT9. (A) Nucleotide binding pocket of GppNHp and GDP bound SEPT2 (PDB-IDs 3FTQ and 6UPA, respectively). Note the significant conformational variance between the two nucleotide states. The positions of the catalytically active Thr in switch 1 and Gly in switch 2 are indicated. (B) Nucleotide binding pocket of GTPγS and GDP bound SEPT9 (PDB-IDs 5CYP and 5CYO, respectively). Note the conformational consistency between the two nucleotide states. The positions of the catalytically active Thr in switch 1 and Gly in switch 2 are indicated. Alignments of the structures in A and B were made in Pymol.
Experimentally, GTP binding and hydrolysis of the septins was mainly assessed by introducing mutations in vivo at positions presumably being involved in nucleotide binding and hydrolysis and monitoring the effect on the phenotype of the respective cells. Mutations were selected based on sequence homologies with small GTPases like Ras or with the available septin crystal structures but not fostered by complementing biochemical assays. The results of these studies are reviewed elsewhere (Abbey et al. 2019; Field and Kellogg 1999; Mitchison and Field 2002). We will focus in the following on studies involving biochemical investigations. The results of all studies considered are summarized in Table 1.
Compilation of biochemical analyses regarding nucleotide uptake and GTP hydrolysis with denatured (*) or native (°) septin protein.
| Species | Septin | Source | Nucleotide uptake | GTP hydrolysis | Reference |
|---|---|---|---|---|---|
| Drosophila m. | PNUT/SEP1/SEP2 complex | Native | Yes | Yes, slow* | Field et al. (1996) |
| Xenopus l. | SEPT2 | Recombinant | Yes | Yes* | Mendoza et al. (2002) |
| Human | SEPT2 | Recombinant | Yes | Yes* | Kinoshita et al. (1997) |
| Human | SEPT2 | Recombinant | Yes, Mg2+ conc. Dependent for GTP | Yes, Mg2+dependent* | Huang et al. (2006) |
| Native | |||||
| Human | SEPT2/6/7 complex | Recombinant | Yes, slow | Yes, slow* | Sheffield et al. (2003) |
| Human | SEPT2, SEPT6, SEPT7, SEPT9 | Recombinant | Yes (all subunits) | Yes* | Zent and Wittinghofer (2014) |
| Human | SEPT2/6/7 hexamer | Recombinant | Yes, Mg2+ independent | Not detectable° | Fischer et al. (2022) |
| SEPT2/6/7/9 octamer | |||||
| S. cerevisiae | Cdc11/12/3/10 octamer | Native | Yes, slow (in Mg2+ presence), no turnover in vivo | Yes, fast* | Vrabioiu et al. (2004) |
| S. cerevisiae | Cdc3, Cdc10, Cdc11, Cdc12 | Recombinant | Yes, GTP only by Cdc10 and Cdc12 | Yes* | Versele et al. (2004) |
| S. cerevisiae | Cdc3/12 | Recombinant | Yes, but not Cdc3. | Yes, slow* | Farkasovsky et al. (2005) |
| Cdc11/12/3 | |||||
| Cdc12/3/10 | All four subunits in quaternary complex. | ||||
| Cdc11/12/3/10 complexes | |||||
| S. cerevisiae | Cdc11/12/3/10 octamer | Native | Yes | Not assayed | Nagaraj et al. (2008) |
| S. cerevisiae | Cdc12/3/10 hexamer | Recombinant | Yes (both complexes) | Not detectable° | Baur et al. (2018) |
| Cdc11/12/3/10 octamer | Mg2+ dependent |
GTP binding and hydrolysis by animal- and human septins
First attempts towards a biochemical investigation of septins were made in the late 1990s when a complex of the Drosophila septins PNUT, SEP1 and SEP2, isolated from Drosophila embryos, was characterized and found to bind and hydrolyze GTP (Field et al. 1996). As in many investigations to follow, nucleotide uptake was assessed by incubating the septin preparation with radioactive labelled GTP in a buffer containing EDTA and MgCl2 and the radioactivity was detected by thin layer chromatography (TLC) after denaturation of the protein.
Following this initial study, SEPT2 (recombinant human SEPT2 and SEPT2 from Xenopus laevis) was also shown to be able to bind and hydrolyze GTP (Kinoshita et al. 1997; Mendoza et al. 2002). Binding- and hydrolysis kinetics of SEPT2 were examined in a more detailed, separate study (Huang et al. 2006). SEPT2 was found to bind and exchange GTP in a Mg2+ concentration dependent manner whereas GDP binding and exchange was Mg2+ independent. Consequently, the authors suggested that SEPT2 has different conformations allowing to discriminate between triphosphate and diphosphate nucleotides. GTP hydrolysis, detected by TLC, was found to be independent of the Mg2+ concentration but was nearly undetectable when Mg2+ was not present. Interestingly, the determined kinetic constants differed between SEPT2 produced in bacteria and SEPT2 produced in a baculovirus expression system. Posttranslational modifications may thus alter the nucleotide binding properties of SEPT2. The detected GTP hydrolysis rate for SEPT2 (0.014 min−1) was confirmed later in a separate study (Zent and Wittinghofer 2014).
The availability of the GDP and GppNHp containing crystal structures of mammalian SEPT2 (PDB ID: 2QA5 (GDP) and later 3FTQ (GppNHp) and 6UPA (GDP)) allowed to establish a first model how septins may hydrolyze GTP (Sirajuddin et al. 2009). Thr78 (Thr(Sw1)) in SEPT2 was identified as the bona fide catalytically active residue. Together with a conserved Gly from the DXXG motif in switch 2 (Gly(Sw2)) it is supposed to mediate the classic “loaded spring” mechanism of Ras-like GTPases (Vetter and Wittinghofer 2001) (Figure 5A). The finding that the catalytically inactive Cdc3 and Cdc11 from yeast do not possess Thr(Sw1) support the idea that this is indeed the active residue. However, in the more recently published structures of SEPT3 and SEPT9 (do Vale Castro et al. 2020), Thr(Sw1) as well as Gly(Sw2) are not in hydrogen bonding distance to the γ-phosphate and in the respective GDP containing structures Thr(Sw1) remains in place close to the nucleotide (Figure 5B). This would contradict the idea of the “loaded spring” mechanism. Despite this finding, SEPT9 was shown to be the human septin subunit with the highest measured GTP hydrolysis rate (0.064 min−1 vs. 0.014 min−1 for SEPT2 and 0.0063 min−1 for SEPT7) (Zent and Wittinghofer 2014).
Entire human septin complexes were also assayed for nucleotide binding. The SEPT2/6/7 complex was found to bind and release GTP at slow pace with the half time of the release of previously bound [32P]-αGTP being approx. 150 min (Sheffield et al. 2003). Various concentrations of EDTA did not alter the nucleotide binding and slow GTP hydrolysis was detected by TLC. Within the recorded time span (120 min) the reaction followed a linear kinetics and thus half times could not be calculated.
More recently, human octameric septin complexes including SEPT9 became available and were assayed along SEPT2/6/7 complexes for nucleotide binding and hydrolysis employing [32P]-γGTP (Fischer et al. 2022). Both complexes bound nucleotide following a one phase exponential association kinetics with half times of 10 min (hexamer) and 16 min (octamer), respectively. Presence of Mg2+ led to slightly and barely significantly prolonged half times.
Release of previously bound [32P]-γGTP from the octamer had a half time of 47 min, approx. 3 times faster than previously detected for the hexamer (Sheffield et al. 2003) (see above).
Nucleotide exchange half times (i.e. uptake in complexes previously loaded with non-radioactive nucleotide) were in the range of 40 min, indicating that the septin hexamers and octamers readily exchange once bound nucleotide. Additionally, this indicates that the septin preparation is to a significant extent free of nucleotide prior to loading since the half time of the exchange reaction is 3–4 times slower than the nucleotide uptake of the non-loaded septin preparation. The latter finding is in line with other studies who found that septins expressed in E. coli are to about 50% in the apo form (Mendoza et al. 2002; Huang et al. 2006).
GTP hydrolysis was assayed in this study by detecting the release of radioactivity from the native complex over 120 min in standard GTPase hydrolysis buffer containing Mg2+. During this time span no release of [32P] or [32P]-γGTP could be detected indicating that hydrolysis products are either kept inside the nucleotide binding pocket or that nucleotide hydrolysis occurs so slowly that products are not detectable after 120 min.
Taken together, members from the Momany groups 1a (SEPT3 group) and 2b (SEPT2 group and SEPT7) are catalytically active whereas group 1b members (SEPT6 group) are not (Zent and Wittinghofer 2014). Apparently, groups 1a and 2b members differ in their GTP hydrolysis mechanism. Hydrolysis cannot be detected within the native septin complex.
GTP binding and hydrolysis by yeast septins
Considering the high sequence homology of human and yeast septins one would assume similar biochemical properties in both species. However, septin complexes from yeast differ in several aspects from their human counterparts.
Entire yeast septin complexes were first isolated from S. cerevisiae cells by immunoprecipitation (Vrabioiu et al. 2004). Nucleotide uptake of the isolated complexes was monitored by a filter assay and it was found that uptake takes place at a very low pace with the half time of the reaction being in the hours range. An in vivo turnover assay revealed that the septin subunits did not significantly turnover nucleotide. Strikingly, rapid GTP hydrolysis in the range of the uncatalyzed (without GAP) GTPase reaction of Ras was detected for the isolated complex via TLC. However, this is the only study showing such fast GTP hydrolysis. All other published studies (see below) point towards a slow GTPase reaction in septins and consequently the results provided by Vrabioiu et al. should be taken with care in this respect.
Another study used septin complexes purified by immunoprecipitation from yeast extracts to assess the role of nucleotide binding on complex formation (Nagaraj et al. 2008). GTP binding was confirmed by UV crosslinking of [32P]-γGTP to all four subunits. By introducing mutations in the P-loop and the G3 and G4 motifs of various subunits and subsequent in vivo and in vitro binding assays the authors suggest a role for nucleotide binding in septin-septin interactions but not for complex functionality.
One single study attempted to assay GTP hydrolysis of individual yeast septin subunits. Versele and Thorner (2004) reported the successful purification of recombinant Cdc11, Cdc12, Cdc3 and Cdc10 from E. coli lysates and found that Cdc10 and Cdc12 are catalytically active and Cdc3 and Cdc11 are not. However, they did not address the oligomerization state of their protein preparations. Following studies indicated that Cdc10 and Cdc12 are purified as multimeric aggregates from E. coli lysates (Baur et al. 2018; Farkasovsky et al. 2005). This finding would at least relativize the quantitative findings raised in this study. Mutation of Thr48(P-loop) to Asn in Cdc12 impeded GTP binding of this subunit and cells expressing this mutant in combination with the corresponding mutant in Cdc10 showed temperature sensitive morphology and cytokinesis abnormalities at the restricted temperature, resulting from a failure of septin collar formation in early S phase. Cdc11–Cdc12T(P-loop)N–Cdc3–Cdc10S(P-loop)N rods failed to form filament-like structures in vitro after dialysis in low salt buffer. The authors concluded that GTP binding to Cdc10 and Cdc12 is important for filament formation and septin collar assembly. However, it is not possible to distinguish if especially GTP binding or nucleotide binding in general is the crucial factor as it was not assayed if the septin preparations contain GDP.
A following study aimed at investigating nucleotide binding and hydrolysis of recombinant yeast septin complexes of different compositions (Farkasovsky et al. 2005). The authors tried also to purify individual septin subunits but stated that these were partially insoluble and prone to aggregation, did not contain bound nucleotides and did not bind exogenously added nucleotide. These findings are in line with other published findings (Baur et al. 2018). By binding of [32P]-αGTP and subsequent UV crosslinking, GTP binding was assigned to all subunits within the full complex containing all four subunits. In all other binary or ternary complexes Cdc3 could not bind GTP. Cdc10 and Cdc12 were identified as the only two subunits that exchange nucleotide.
This finding was further corroborated by another study published about a decade later (Baur et al. 2018). Truncated versions of Cdc3 and Cdc11 could be purified as monomeric apo proteins but Cdc10 and Cdc12 tended – in line with the findings of Farkasovski et al. – to aggregate. By analyzing the nucleotide uptake in the G interface dimers Cdc3–Cdc10 and Cdc11–Cdc12 the authors concluded that Cdc10 binds preferably GDP and Cdc12 is the only subunit capable of binding and hydrolyzing GTP. For Cdc10, this is in contrast to previous findings and Cdc10 possesses the Thr(Sw1). However, both investigated G interface dimers formed not dimers but rather tetramers or even larger assemblies composed of the two respective subunits. Thus, these septin subunits may not represent the physiological conformation and may behave differently than in the assembled rod. Hexameric (Cdc11-less) and octameric rods displayed similar nucleotide binding properties and it was consequently concluded that Cdc11 does not turnover nucleotide, in line with the findings of Farkasovski et al. Unlike in human septin complexes, Mg2+ enhanced the affinity for the nucleotide (halftimes of the uptake reaction 7 min for octamers and 10 min for hexamers) and led to a stabilization of the nucleotide in the binding pocket. Hydrolysis products could not be detected in the timeframe of the applied assay employing native septin protein, as also found for human septin complexes (see above).
The fifth mitotic septin, Shs1, was not yet assayed for nucleotide binding and hydrolysis in the context of defined recombinantly produced complexes but it does not possess the Thr(Sw1).
Taken together, S. cerevisiae seems to have only two catalytically active septin subunits, Cdc10 and Cdc12. They seem to differ mechanistically from their human counterparts.
Synopsis
Taken all findings from mammalian and yeast septins together, only few similarities between the different septin species can be extracted: All tested septin complexes have nucleotide bound with a GDP:GTP ratio of approx. 2:1 and all complexes are able to exchange once bound nucleotide in vitro. With one exception (Vrabioiu et al. 2004), the studies who tested GTP hydrolysis detected slow GTP hydrolysis rates. In octameric rods, both from human and yeast, the central subunit is GTPase active.
In all other respects the picture is less clear: Recombinantly expressed septins appear as a mixture of nucleotide containing and apo protein after purification and nucleotide exchange properties differ between the species. Particularly, nucleotide uptake appears to be independent of the presence of Mg2+ in human septin complexes (Fischer et al. 2022) whereas Mg2+ seem to increase the affinity for nucleotide in yeast septins (Baur et al. 2018).
The canonical human octamer (SEPT2-6-7-9-9-6-7-2) has one GTPase inactive subunit in the second position whereas the yeast octamer Cdc11-12-3-10-10-3-12-11 is composed of two inactive subunits in the first and third position. This might indicate that the assembly of the rods from individual subunits might follow different mechanisms in different species.
The role of the guanine nucleotide binding in septin function remains thus a subject of debate but it is suggested and commonly accepted that guanine nucleotide binding supports septin folding and stability. Since hydrolysis products are not released in vitro under native conditions (Baur et al. 2018; Fischer et al. 2022), we favor a model in which GTP hydrolysis occurs only once during the formation of septin rods, however with species specific mechanisms.
For some GTPase active subunits a loaded spring mechanism can be predicted from the available structural data (e.g. SEPT2) and for others – although catalytically active – not (group 1b septins including SEPT9). Mechanistically, GTP hydrolysis in septins remains a conundrum and more structural information, especially from yeast septins, as well as biochemical data are needed to develop a comprehensive model of nucleotide binding and hydrolysis on a molecular level.
About the authors

Benjamin Grupp graduated in biochemistry and is currently PhD student at the University of Ulm/Germany. He applies in vitro and in silico approaches to study biochemical features of yeast septins.

Thomas Gronemeyer studied biochemistry in Bochum/Germany and obtained a PhD in protein engineering from the Federal Institute of Technology in Lausanne/Switzerland. Today he is permanent staff scientist and lecturer for molecular- and cell biology at the University of Ulm. His research focus lies on the biochemistry of yeast and mammalian septins.
-
Author contribution: All the authors have accepted responsibility for the entire content of this submitted manuscript and approved submission.
-
Research funding: None declared.
-
Conflict of interest statement: The authors declare no conflicts of interest regarding this article.
References
Abbey, M., Gaestel, M., and Menon, M.B. (2019). Septins: active GTPases or just GTP‐binding proteins? Cytoskeleton 76: 55–62, https://doi.org/10.1002/cm.21451.Search in Google Scholar PubMed
Auxier, B., Dee, J., Berbee, M.L., and Momany, M. (2019). Diversity of opisthokont septin proteins reveals structural constraints and conserved motifs. BMC Evol. Biol. 19, https://doi.org/10.1186/s12862-018-1297-8.Search in Google Scholar PubMed PubMed Central
Bartsch, I., Sandrock, K., Lanza, F., Nurden, P., Hainmann, I., Pavlova, A., Greinacher, A., Tacke, U., Barth, M., Busse, A., et al.. (2011). Deletion of human GP1BB and SEPT5 is associated with Bernard-Soulier syndrome, platelet secretion defect, polymicrogyria, and developmental delay. Thromb. Haemostasis 106: 475–483, https://doi.org/10.1160/th11-05-0305.Search in Google Scholar
Baur, J.D., Rösler, R., Wiese, S., Johnsson, N., and Gronemeyer, T. (2018). Dissecting the nucleotide binding properties of the septins from S. cerevisiae. Cytoskeleton 76: 45–54, https://doi.org/10.1002/cm.21484.Search in Google Scholar PubMed
Beber, A., Taveneau, C., Nania, M., Tsai, F.C., Di Cicco, A., Bassereau, P., Lévy, D., Cabral, J.T., Isambert, H., Mangenot, S., et al.. (2019). Membrane reshaping by micrometric curvature sensitive septin filaments. Nat. Commun. 10: 420, https://doi.org/10.1038/s41467-019-08344-5.Search in Google Scholar PubMed PubMed Central
Bertin, A., Mcmurray, M.A., Grob, P., Park, S.-S., Iii, G.G., Patanwala, I., Ng, H.-L., Alber, T., Thorner, J., Nogales, E., et al.. (2008). Saccharomyces cerevisiae septins: supramolecular organization of heterooligomers and the mechanism of filament assembly. Proc. Natl. Acad. Sci. U.S.A. 105: 8274–8279, https://doi.org/10.1073/pnas.0803330105.Search in Google Scholar PubMed PubMed Central
Bertin, A., McMurray, M.A., Thai, L., Garcia, G., Votin, V., Grob, P., Allyn, T., Thorner, J., and Nogales, E. (2010). Phosphatidylinositol-4, 5-bisphosphate promotes budding yeast septin filament assembly and organization. J. Mol. Biol. 404: 711–731, https://doi.org/10.1016/j.jmb.2010.10.002.Search in Google Scholar PubMed PubMed Central
Bi, E. and Park, H.O. (2012). Cell polarization and cytokinesis in budding yeast. Genetics 191: 347–387, https://doi.org/10.1534/genetics.111.132886.Search in Google Scholar PubMed PubMed Central
Brausemann, A., Gerhardt, S., Schott, A.-K., Einsle, O., Große-Berkenbusch, A., Johnsson, N., and Gronemeyer, T. (2016). Crystal structure of Cdc11, a septin subunit from Saccharomyces cerevisiae. J. Struct. Biol. 193: 157–161, https://doi.org/10.1016/j.jsb.2016.01.004.Search in Google Scholar PubMed
Calixto, A.R., Moreira, C., Pabis, A., Kötting, C., Gerwert, K., Rudack, T., and Kamerlin, S.C.L. (2019). GTP hydrolysis without an active site base: a unifying mechanism for Ras and related GTPases. J. Am. Chem. Soc. 141: 10684–10701, https://doi.org/10.1021/jacs.9b03193.Search in Google Scholar PubMed
Carvalho, A.T.P., Szeler, K., Vavitsas, K., Åqvist, J., and Kamerlin, S.C.L. (2015). Modeling the mechanisms of biological GTP hydrolysis. Arch. Biochem. Biophys. 582: 80–90, https://doi.org/10.1016/j.abb.2015.02.027.Search in Google Scholar PubMed
Cavini, I.A., Leonardo, D.A., Rosa, H.V.D., Castro, D.K.S.V., D’Muniz Pereira, H., Valadares, N.F., Araujo, A.P.U., and Garratt, R.C. (2021). The structural biology of septins and their filaments: an update. Front. Cell Dev. Biol. 9, https://doi.org/10.3389/fcell.2021.765085.Search in Google Scholar PubMed PubMed Central
do Vale Castro, D.K.S., de Oliveira da Silva, S.M., Pereira, H.D., Macedo, J.N.A., Leonardo, D.A., Valadares, N.F., Kumagai, P.S., Brandão-Neto, J., Araújo, A.P.U., and Garratt, R.C. (2020). A complete compendium of crystal structures for the human SEPT3 subgroup reveals functional plasticity at a specific septin interface. IUCrJ 7: 462–479, https://doi.org/10.1107/s2052252520002973.Search in Google Scholar
Dolat, L., Hu, Q., and Spiliotis, E.T. (2014). Septin functions in organ system physiology and pathology. Biol. Chem. 395: 123–141, https://doi.org/10.1515/hsz-2013-0233.Search in Google Scholar PubMed PubMed Central
Estey, M.P., Di Ciano-Oliveira, C., Froese, C.D., Bejide, M.T., and Trimble, W.S. (2010). Distinct roles of septins in cytokinesis: SEPT9 mediates midbody abscission. J. Cell Biol. 191: 741–749, https://doi.org/10.1083/jcb.201006031.Search in Google Scholar PubMed PubMed Central
Fares, H., Goetsch, L., and Pringle, J.R. (1996). Identification of a developmentally regulated septin and involvement of the septins in spore formation in Saccharomyces cerevisiae. J. Cell Biol. 132: 399–411, https://doi.org/10.1083/jcb.132.3.399.Search in Google Scholar PubMed PubMed Central
Farkasovsky, M., Herter, P., Voss, B., and Wittinghofer, A. (2005). Nucleotide binding and filament assembly of recombinant yeast septin complexes. Biol. Chem. 386: 643–656, https://doi.org/10.1515/bc.2005.075.Search in Google Scholar PubMed
Field, C.M., Al-Awar, O., Rosenblatt, J., Wong, M.L., Alberts, B., and Mitchison, T.J. (1996). A purified Drosophila septin complex forms filaments and exhibits GTPase activity. J. Cell Biol. 133: 605–616, https://doi.org/10.1083/jcb.133.3.605.Search in Google Scholar PubMed PubMed Central
Field, C.M. and Kellogg, D. (1999). Septins: cytoskeletal polymers or signalling GTPases? Trends Cell Biol. 9: 387–394, https://doi.org/10.1016/s0962-8924(99)01632-3.Search in Google Scholar PubMed
Finnigan, G.C., Booth, E.A., Duvalyan, A., Liao, E.N., and Thorner, J. (2015a). The carboxy-terminal tails of septins Cdc11 and Shs1 recruit myosin-II binding factor Bni5 to the bud neck in Saccharomyces cerevisiae. Genetics 200: 843–862, https://doi.org/10.1534/genetics.115.176503.Search in Google Scholar PubMed PubMed Central
Finnigan, G.C., Takagi, J., Cho, C., and Thorner, J. (2015b). Comprehensive genetic analysis of paralogous terminal septin subunits Shs1 and Cdc11 in Saccharomyces cerevisiae. Genetics 200: 821–841, https://doi.org/10.1534/genetics.115.176495.Search in Google Scholar PubMed PubMed Central
Fischer, M., Frank, D., Rösler, R., Johnsson, N., and Gronemeyer, T. (2022). Biochemical characterization of a human septin octamer. Front. Cell Dev. Biol. 10: 771388, https://doi.org/10.3389/fcell.2022.771388.Search in Google Scholar PubMed PubMed Central
Fonseca Valadares, N., Pereira, M., Paula Ulian Araujo, A., and Charles Garratt, R. (2017). Septin structure and filament assembly. Biophys. Rev. 9: 481–500, https://doi.org/10.1007/s12551-017-0320-4.Search in Google Scholar PubMed PubMed Central
Garcia, G., Bertin, A., Li, Z., Song, Y., Mcmurray, M.A., Thorner, J., and Nogales, E. (2011). Subunit-dependent modulation of septin assembly: budding yeast septin Shs1 promotes ring and gauze formation. J. Cell Biol. 195: 993–1004, https://doi.org/10.1083/jcb.201107123.Search in Google Scholar PubMed PubMed Central
Garcia, G., Finnigan, G.C., Heasley, L.R., Sterling, S.M., Aggarwal, A., Pearson, C.G., Nogales, E., McMurray, M.A., and Thorner, J. (2016). Assembly, molecular organization, and membrane-binding properties of development-specific septins. J. Cell Biol. 212: 515–529, https://doi.org/10.1083/jcb.201511029.Search in Google Scholar PubMed PubMed Central
Hartwell, L. (1971). Genetic control of the cell division cycle in yeast *1IV. Genes controlling bud emergence and cytokinesis. Exp. Cell Res. 69: 265–276, https://doi.org/10.1016/0014-4827(71)90223-0.Search in Google Scholar PubMed
Hernández-Rodríguez, Y., Masuo, S., Johnson, D., Orlando, R., Smith, A., Couto-Rodriguez, M., and Momany, M. (2014). Distinct septin heteropolymers co-exist during multicellular development in the filamentous fungus Aspergillus nidulans. PLoS One 9, https://doi.org/10.1371/journal.pone.0092819.Search in Google Scholar PubMed PubMed Central
Hernández-Rodríguez, Y. and Momany, M. (2012). Posttranslational modifications and assembly of septin heteropolymers and higher-order structures. Curr. Opin. Microbiol. 15: 660–668, https://doi.org/10.1016/j.mib.2012.09.007.Search in Google Scholar PubMed
Huang, Y.W., Surka, M.C., Reynaud, D., Pace-Asciak, C., and Trimble, W.S. (2006). GTP binding and hydrolysis kinetics of human septin 2. FEBS J. 273: 3248–3260, https://doi.org/10.1111/j.1742-4658.2006.05333.x.Search in Google Scholar PubMed
Iv, F., Martins, C.S., Castro-Linares, G., Taveneau, C., Barbier, P., Verdier-Pinard, P., Camoin, L., Audebert, S., Tsai, F.-C., Ramond, L., et al.. (2021). Insights into animal septins using recombinant human septin octamers with distinct SEPT9 isoforms. J. Cell Sci. 134: jcs258484, https://doi.org/10.1242/jcs.258484.Search in Google Scholar PubMed
Ivanov, A.I., Le, H.T., Naydenov, N.G., and Rieder, F. (2021). Novel functions of the septin cytoskeleton: shaping up tissue inflammation and fibrosis. Am. J. Pathol. 191: 40–51, https://doi.org/10.1016/j.ajpath.2020.09.007.Search in Google Scholar PubMed PubMed Central
Johnson, C.R., Weems, A.D., Brewer, J.M., Thorner, J., and McMurray, M.A. (2015). Cytosolic chaperones mediate quality control of higher-order septin assembly in budding yeast. Mol. Biol. Cell 26: 1323–1344, https://doi.org/10.1091/mbc.e14-11-1531.Search in Google Scholar PubMed PubMed Central
Johnson, E.S. and Blobel, G. (1999). Cell cycle-regulated attachment of the ubiquitin-related protein SUMO to the yeast septins. J. Cell Biol. 147: 981–993, https://doi.org/10.1083/jcb.147.5.981.Search in Google Scholar PubMed PubMed Central
Khan, A., Newby, J., and Gladfelter, A.S. (2018). Control of septin filament flexibility and bundling by subunit composition and nucleotide interactions. Mol. Biol. Cell 29: 702–712, https://doi.org/10.1091/mbc.e17-10-0608.Search in Google Scholar PubMed PubMed Central
Kinoshita, M. (2003). Assembly of mammalian septins. J. Biochem. 134: 491–496, https://doi.org/10.1093/jb/mvg182.Search in Google Scholar PubMed
Kinoshita, M., Kumar, S., Mizoguchi, A., Ide, C., Kinoshita, A., Haraguchi, T., Hiraoka, Y., and Noda, M. (1997). Nedd5, a mammalian septin, is a novel cytoskeletal component interacting with actin-based structures. Genes Dev. 11: 1535–1547, https://doi.org/10.1101/gad.11.12.1535.Search in Google Scholar PubMed
Kozubowski, L., Larson, J.R., and Tatchell, K. (2005). Role of the septin ring in the asymmetric localization of proteins at the mother-bud neck in Saccharomyces cerevisiae. Mol. Biol. Cell 16: 3455–3466, https://doi.org/10.1091/mbc.e04-09-0764.Search in Google Scholar PubMed PubMed Central
Leonardo, D.A., Cavini, I.A., Sala, F.A., Mendonça, D.C., Rosa, H.V.D., Kumagai, P.S., Crusca, E., Valadares, N.F., Marques, I.A., Brandão-Neto, J., et al.. (2021). Orientational ambiguity in septin coiled coils and its structural basis. J. Mol. Biol. 433, https://doi.org/10.1016/j.jmb.2021.166889.Search in Google Scholar PubMed
Longo, L.M., Jabtoiiska, J., Vyas, P., Kanade, M., Kolodny, R., Ben-Tal, N., and Tawfik, D.S. (2020). On the emergence of P-Loop NTPase and Rossmann enzymes from a Beta-Alpha-Beta ancestral fragment. Elife 9: 1–16, https://doi.org/10.7554/elife.64415.Search in Google Scholar
Macara, I.G., Baldarelli, R., Field, C.M., Glotzer, M., Hayashi, Y., Hsu, S.C., Kennedy, M.B., Kinoshita, M., Longtine, M., Low, C., et al.. (2002). Mammalian septins nomenclature. Mol. Biol. Cell 13: 4111–4113, https://doi.org/10.1091/mbc.e02-07-0438.Search in Google Scholar PubMed PubMed Central
Marcus, J., Bejerano-Sagie, M., Patterson, N., Bagchi, S., Verkhusha, V.V., Connolly, D., Goldberg, G.L., Golden, A., Sharma, V.P., Condeelis, J., et al.. (2019). Septin 9 isoforms promote tumorigenesis in mammary epithelial cells by increasing migration and ECM degradation through metalloproteinase secretion at focal adhesions. Oncogene 38: 5839–5859, https://doi.org/10.1038/s41388-019-0844-0.Search in Google Scholar PubMed PubMed Central
Mavrakis, M., Azou-Gros, Y., Tsai, F.-C., Alvarado, J., Bertin, A., Iv, F., Kress, A., Brasselet, S., Koenderink, G.H., and Lecuit, T. (2014). Septins promote F-actin ring formation by crosslinking actin filaments into curved bundles. Nat. Cell Biol. 16: 322–334, https://doi.org/10.1038/ncb2921.Search in Google Scholar PubMed
Mendonça, D.C., Guimarães, S.L., Pereira, H.D., Pinto, A.A., de Farias, M.A., de Godoy, A.S., Araujo, A.P.U., van Heel, M., Portugal, R.V., and Garratt, R.C. (2021). An atomic model for the human septin hexamer by cryo-EM. J. Mol. Biol. 433: 167096, https://doi.org/10.1016/j.jmb.2021.167096.Search in Google Scholar PubMed
Mendoza, M., Hyman, A.A., and Glotzer, M. (2002). GTP binding induces filament assembly of a recombinant septin. Curr. Biol. 12: 1858–1863, https://doi.org/10.1016/s0960-9822(02)01258-7.Search in Google Scholar PubMed
Menon, M.B., and Gaestel, M. (Eds.), (2017). Emerging Functions of septins. Frontiers Media, Lausanne.10.3389/978-2-88945-287-3Search in Google Scholar
Mino, A., Tanaka, K., Kamei, T., Umikawa, M., Fujiwara, T., and Takai, Y. (1998). Shs1p: a novel member of septin that interacts with spa2p, involved in polarized growth in saccharomyces cerevisiae. Biochem. Biophys. Res. Commun. 251: 732–736, https://doi.org/10.1006/bbrc.1998.9541.Search in Google Scholar PubMed
Mitchison, T.J. and Field, C.M. (2002). Cytoskeleton: what does GTP do for septins? Curr. Biol. 12: R788–R790, https://doi.org/10.1016/s0960-9822(02)01295-2.Search in Google Scholar PubMed
Nagaraj, S., Rajendran, A., Jackson, C.E., and Longtine, M.S. (2008). Role of nucleotide binding in septin-septin interactions and septin localization in Saccharomyces cerevisiae. Mol. Cell Biol. 28: 5120–5137, https://doi.org/10.1128/mcb.00786-08.Search in Google Scholar PubMed PubMed Central
Oh, Y. and Bi, E. (2011). Septin structure and function in yeast and beyond. Trends Cell Biol. 21: 141–148, https://doi.org/10.1016/j.tcb.2010.11.006.Search in Google Scholar PubMed PubMed Central
Ong, K., Wloka, C., Okada, S., Svitkina, T., and Bi, E. (2014). Architecture and dynamic remodelling of the septin cytoskeleton during the cell cycle. Nat. Commun. 5: 5698, https://doi.org/10.1038/ncomms6698.Search in Google Scholar PubMed PubMed Central
Pan, F., Malmberg, R.L., and Momany, M. (2007). Analysis of septins across kingdoms reveals orthology and new motifs. BMC Evol. Biol. 7: 103, https://doi.org/10.1186/1471-2148-7-103.Search in Google Scholar PubMed PubMed Central
Pinto, A.P.A., Pereira, H.M., Zeraik, A.E., Ciol, H., Ferreira, F.M., Brandão-Neto, J., DeMarco, R., Navarro, M.V.A.S., Risi, C., Galkin, V.E., et al.. (2017). Filaments and fingers: novel structural aspects of the single septin from Chlamydomonas reinhardtii. J. Biol. Chem. 292: 10899–10911, https://doi.org/10.1074/jbc.m116.762229.Search in Google Scholar PubMed PubMed Central
Renz, C., Johnsson, N., and Gronemeyer, T. (2013). An efficient protocol for the purification and labeling of entire yeast septin rods from E.coli for quantitative in vitro experimentation. BMC Biotechnol. 13: 60, https://doi.org/10.1186/1472-6750-13-60.Search in Google Scholar PubMed PubMed Central
Ribet, D., Boscaini, S., Cauvin, C., Siguier, M., Mostowy, S., Echard, A., and Cossart, P. (2017). SUMOylation of human septins is critical for septin filament bundling and cytokinesis. J. Cell Biol. 216: 4041–4052, https://doi.org/10.1083/jcb.201703096.Search in Google Scholar PubMed PubMed Central
Rosa, H.V.D., Leonardo, D.A., Brognara, G., Brandão-Neto, J., D’Muniz Pereira, H., Araújo, A.P.U., and Garratt, R.C. (2020). Molecular recognition at septin interfaces: the switches hold the key. J. Mol. Biol. 432: 5784–5801, https://doi.org/10.1016/j.jmb.2020.09.001.Search in Google Scholar PubMed
Rudack, T., Xia, F., Schlitter, J., Kötting, C., and Gerwert, K. (2012). The role of magnesium for geometry and charge in GTP hydrolysis, revealed by quantum mechanics/molecular mechanics simulations. Biophys. J. 103: 293–302, https://doi.org/10.1016/j.bpj.2012.06.015.Search in Google Scholar PubMed PubMed Central
Sanders, S.L. and Field, C.M. (1994). Cell division. Septins in common? Curr. Biol. 4: 907–910, https://doi.org/10.1016/s0960-9822(00)00201-3.Search in Google Scholar PubMed
Sellin, M.E., Holmfeldt, P., Stenmark, S., and Gullberg, M. (2011). Microtubules support a disk-like septin arrangement at the plasma membrane of mammalian cells. Mol. Biol. Cell 22: 4588–4601, https://doi.org/10.1091/mbc.e11-09-0754.Search in Google Scholar
Sheffield, P.J., Oliver, C.J., Kremer, B.E., Sheng, S., Shao, Z., and Macara, I.G. (2003). Borg/septin interactions and the assembly of mammalian septin heterodimers, trimers, and filaments. J. Biol. Chem. 278: 3483–3488, https://doi.org/10.1074/jbc.m209701200.Search in Google Scholar PubMed
Shen, Y.-R., Wang, H.-Y., Kuo, Y.-C., Shih, S.-C., Hsu, C.-H., Chen, Y.-R., Wu, S.-R., Wang, C.-Y., and Kuo, P.-L. (2017). SEPT12 phosphorylation results in loss of the septin ring/sperm annulus, defective sperm motility and poor male fertility. PLoS Genet. 13: e1006631, https://doi.org/10.1371/journal.pgen.1006631.Search in Google Scholar PubMed PubMed Central
Shuman, B. and Momany, M. (2022). Septins from protists to people. Front. Cell Dev. Biol. 9: 824850, https://doi.org/10.3389/fcell.2021.824850.Search in Google Scholar PubMed PubMed Central
Sirajuddin, M., Farkasovsky, M., Hauer, F., Kühlmann, D., Macara, I.G., Weyand, M., Stark, H., and Wittinghofer, A. (2007). Structural insight into filament formation by mammalian septins. Nature 449: 311–315, https://doi.org/10.1038/nature06052.Search in Google Scholar PubMed
Sirajuddin, M., Farkasovsky, M., Zent, E., Wittinghofer, A., and Fersht, A. (2009). GTP-induced conformational changes in septins and implications for function. Proc. Natl. Acad. Sci. U.S.A. 106: 16592–16597, https://doi.org/10.1073/pnas.0902858106.Search in Google Scholar PubMed PubMed Central
Smith, C., Dolat, L., Angelis, D., Forgacs, E., Spiliotis, E.T., and Galkin, V.E. (2015). Septin 9 exhibits polymorphic binding to F-actin and inhibits myosin and cofilin activity. J. Mol. Biol. 427: 3273–3284, https://doi.org/10.1016/j.jmb.2015.07.026.Search in Google Scholar PubMed PubMed Central
Soroor, F., Kim, M.S., Palander, O., Balachandran, Y., Collins, R.F., Benlekbir, S., Rubinstein, J., and Trimble, W.S. (2020). Revised subunit order of mammalian septin complexes explains their in-vitro polymerization properties. Mol. Biol. Cell 32: 289–300, https://doi.org/10.1091/mbc.e20-06-0398.Search in Google Scholar
Tokhtaeva, E., Capri, J., Marcus, E.A., Whitelegge, J.P., Khuzakhmetova, V., Bukharaeva, E., Deiss-Yehiely, N., Dada, L.A., Sachs, G., Fernandez-Salas, E., et al.. (2015). Septin dynamics are essential for exocytosis. J. Biol. Chem. 290: 5280–5297, https://doi.org/10.1074/jbc.m114.616201.Search in Google Scholar PubMed PubMed Central
Versele, M., Gullbrand, B., Shulewitz, M.J., Cid, V.J., Bahmanyar, S., Chen, R.E., Barth, P., Alber, T., and Thorner, J. (2004). Protein–protein interactions governing septin heteropentamer assembly and septin filament organization in Saccharomyces cerevisiae. Mol. Biol. Cell 15: 4568–4583, https://doi.org/10.1091/mbc.e04-04-0330.Search in Google Scholar PubMed PubMed Central
Versele, M. and Thorner, J. (2004). Septin collar formation in budding yeast requires GTP binding and direct phosphorylation by the PAK, Cla4. J. Cell Biol. 164: 701–715, https://doi.org/10.1083/jcb.200312070.Search in Google Scholar PubMed PubMed Central
Vetter, I.R. and Wittinghofer, A. (2001). The guanine nucleotide – binding switch in three dimensions. Science 294: 1299–1305, https://doi.org/10.1126/science.1062023.Search in Google Scholar PubMed
De Virgilio, C., DeMarini, D.J., and Pringle, J.R. (1996). SPR28, a sixth member of the septin gene family in Saccharomyces cerevisiae that is expressed specifically in sporulating cells. Microbiology 142: 2897–2905, https://doi.org/10.1099/13500872-142-10-2897.Search in Google Scholar PubMed
Vrabioiu, A.M., Gerber, S.A., Gygi, S.P., Field, C.M., and Mitchison, T.J. (2004). The majority of the Saccharomyces cerevisiae septin complexes do not exchange guanine nucleotides. J. Biol. Chem. 279: 3111–3118, https://doi.org/10.1074/jbc.m310941200.Search in Google Scholar
Wang, H.Y., Lin, C.H., Shen, Y.R., Chen, T.Y., Wang, C.Y., and Kuo, P.L. (2021). Protein kinase A-mediated Septin7 phosphorylation disrupts septin filaments and ciliogenesis. Cells 10: 1–16, https://doi.org/10.3390/cells10020361.Search in Google Scholar PubMed PubMed Central
Weems, A. and McMurray, M. (2017). The step-wise pathway of septin hetero-octamer assembly in budding yeast. Elife 6: e23689, https://doi.org/10.7554/elife.23689.Search in Google Scholar
Weems, A.D., Johnson, C.R., Argueso, J.L., and McMurray, M.A. (2014). Higher-order septin assembly is driven by GTP-promoted conformational changes: evidence from unbiased mutational analysis in Saccharomyces cerevisiae. Genetics 196: 711–727, https://doi.org/10.1534/genetics.114.161182.Search in Google Scholar PubMed PubMed Central
Weirich, C.S., Erzberger, J.P., and Barral, Y. (2008). The septin family of GTPases: architecture and dynamics. Nat. Rev. Mol. Cell Biol. 9: 478–489, https://doi.org/10.1038/nrm2407.Search in Google Scholar PubMed
Werner, B. and Yadav, S. (2022). Phosphoregulation of the septin cytoskeleton in neuronal development and disease. Cytoskeleton, [Preprint], https://doi.org/10.1002/cm.21728.Search in Google Scholar PubMed
Woods, B.L., Cannon, K.S., Vogt, E.J.D., Crutchley, J.M., and Gladfelter, A.S. (2021). Interplay of septin amphipathic helices in sensing membrane-curvature and filament bundling. Mol. Biol. Cell 32: 1–9, https://doi.org/10.1091/mbc.e20-05-0303.Search in Google Scholar PubMed PubMed Central
Zent, E., Vetter, I., and Wittinghofer, A. (2011). Structural and biochemical properties of Sept7, a unique septin required for filament formation. Biol. Chem. 392: 791–797, https://doi.org/10.1515/bc.2011.082.Search in Google Scholar
Zent, E. and Wittinghofer, A. (2014). Human septin isoforms and the GDP-GTP cycle. Biol. Chem. 395: 169–180, https://doi.org/10.1515/hsz-2013-0268.Search in Google Scholar PubMed
Zeraik, A.E., Pereira, H.M., Santos, Y.V., Brandão-Neto, J., Spoerner, M., Santos, M.S., Colnago, L.A., Garratt, R.C., Araújo, A.P.U., and DeMarco, R. (2014). Crystal structure of a Schistosoma mansoni septin reveals the phenomenon of strand slippage in septins dependent on the nature of the bound nucleotide. J. Biol. Chem. 289: 7799–7811, https://doi.org/10.1074/jbc.m113.525352.Search in Google Scholar PubMed PubMed Central
Zhang, B., Zhang, Y., Wang, Z.X., and Zheng, Y. (2000). The role of Mg2+ cofactor in the guanine nucleotide exchange and GTP hydrolysis reactions of Rho family GTP-binding proteins. J. Biol. Chem. 275: 25299–25307, https://doi.org/10.1074/jbc.m001027200.Search in Google Scholar
Zhang, J., Kong, C., Xie, H., McPherson, P.S., Grinstein, S., and Trimble, W.S. (1999). Phosphatidylinositol polyphosphate binding to the mammalian septin H5 is modulated by GTP. Curr. Biol. 9: 1458–1467, https://doi.org/10.1016/s0960-9822(00)80115-3.Search in Google Scholar PubMed
Ziman, M., O, J.M., Ouellette, L.A., Church, W.R., and Johnson, D.I. (1991). Mutational analysis of CDC42Sc, a Saccharomyces cerevisiae gene that encodes a putative GTP-binding protein involved in the control of cell polarity. Mol. Cell Biol. 11: 3537–3544, https://doi.org/10.1128/mcb.11.7.3537-3544.1991.Search in Google Scholar PubMed PubMed Central
© 2022 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Frontmatter
- Review
- A biochemical view on the septins, a less known component of the cytoskeleton
- Research Articles/Short Communications
- Protein Structure and Function
- Mathematical expressions describing enzyme velocity and inhibition at high enzyme concentration
- Cell Biology and Signaling
- MicroRNA-6838-5p suppresses the self-renewal and metastasis of human liver cancer stem cells through downregulating CBX4 expression and inactivating ERK signaling
- M2 macrophages-derived exosomal miR-3917 promotes the progression of lung cancer via targeting GRK6
- Extracellular stimulation of lung fibroblasts with arachidonic acid increases interleukin 11 expression through p38 and ERK signaling
- EVADR ceRNA transcript variants upregulate WNT and PI3K signaling pathways in SW480 and HCT116 cells by sponging miR-7 and miR-29b
Articles in the same Issue
- Frontmatter
- Review
- A biochemical view on the septins, a less known component of the cytoskeleton
- Research Articles/Short Communications
- Protein Structure and Function
- Mathematical expressions describing enzyme velocity and inhibition at high enzyme concentration
- Cell Biology and Signaling
- MicroRNA-6838-5p suppresses the self-renewal and metastasis of human liver cancer stem cells through downregulating CBX4 expression and inactivating ERK signaling
- M2 macrophages-derived exosomal miR-3917 promotes the progression of lung cancer via targeting GRK6
- Extracellular stimulation of lung fibroblasts with arachidonic acid increases interleukin 11 expression through p38 and ERK signaling
- EVADR ceRNA transcript variants upregulate WNT and PI3K signaling pathways in SW480 and HCT116 cells by sponging miR-7 and miR-29b