Lipid antigens in immunity
-
C. Marie Dowds
C. Marie Dowds studied Biosciences at the University of Kaiserslautern, Germany. After completion of her master’s degree in the group of Dr. Stefan Kins, she joined the laboratory of Dr. Zeissig at the Department of Internal Medicine, Kiel University (Kiel, Germany) as PhD student. In her PhD studies, Dowds investigates the role of NKT cells and lipid antigens in intestinal inflammation.
Sabin-Christin Kornell studied Biology at the Christian-Albrechts-University of Kiel and received her master’s degree in cell biology and physiology in 2013. In May 2013, Kornell started her PhD studies in Dr. Zeissig’s group at the Department of Internal Medicine, Kiel University, Germany (Kiel, Germany).
Richard S. Blumberg, MD, is Professor of Medicine at Harvard Medical School, Chief of the Division of Gastroenterology, Hepatology and Endoscopy at Brigham and Women’s Hospital, co-Director of the Biomedical Research Institute at Brigham and Women’s Hospital and co-Director of the Harvard Digestive Diseases Center. He received his medical degree from Jefferson Medical College (Philadelphia, PA), internship and residency at The New York Hospital, Cornell Medical Center followed by fellowship training in infectious diseases at Massachusetts General Hospital, gastroenterology at Brigham and Women’s Hospital and molecular immunology at Dana Farber Cancer Institute, all in Boston. His work focuses on the mechanisms that control homeostasis and inflammation at mucosal surfaces.
 und
Sebastian Zeissig
und
Sebastian Zeissig
Sebastian Zeissig, MD, received his medical degree from the Charité Berlin, Germany. After a postdoctoral fellowship at the same institution, Dr. Zeissig joined the laboratory of Richard S. Blumberg as postdoctoral fellow to investigate the role of CD1d-restricted NKT cells in infectious hepatitis and immunodeficiency. Since 2010, Dr. Zeissig is leading the Laboratory of Mucosal Immunology at the Department of Internal Medicine, Kiel University (Kiel, Germany), where he continues to investigate the role of lipid antigens in intestinal and hepatic immunity.

Abstract
Lipids are not only a central part of human metabolism but also play diverse and critical roles in the immune system. As such, they can act as ligands of lipid-activated nuclear receptors, control inflammatory signaling through bioactive lipids such as prostaglandins, leukotrienes, lipoxins, resolvins, and protectins, and modulate immunity as intracellular phospholipid- or sphingolipid-derived signaling mediators. In addition, lipids can serve as antigens and regulate immunity through the activation of lipid-reactive T cells, which is the topic of this review. We will provide an overview of the mechanisms of lipid antigen presentation, the biology of lipid-reactive T cells, and their contribution to immunity.
CD1 proteins survey subcellular compartments for lipids
Lipid antigens are presented by atypical MHC class I proteins of the CD1 family. In contrast to the polymorphic nature of classical MHC class I, CD1 genes exhibit little sequence diversity with few nonsynonymous single-nucleotide polymorphisms (Zimmer et al., 2009; Seshadri et al., 2013). All mammalian species investigated so far express at least one CD1 protein, suggesting conservation of the CD1 system in mammals (Kasmar et al., 2009). In humans, lipid antigens are presented by group 1 (CD1a, CD1b, and CD1c) and group 2 (CD1d) CD1 isoforms (Porcelli et al., 1989, 1992; Balk et al., 1991; Blumberg et al., 1991; Beckman et al., 1994; Kawano et al., 1997). Mice lack expression of group 1 CD1 and contain two highly homologous CD1d-encoding genes (Cd1d1 and Cd1d2), whereby the Cd1d2 gene contains a frameshift in the C57BL/6 strain that prevents cell surface expression of the encoded protein (Park et al., 1998).
T cells responding to lipid antigens are CD1a-, CD1b-, CD1c-, or CD1d-restricted and, in the case of CD1d, have been termed natural killer T (NKT) cells. Based on their T cell receptor (TCR) repertoire, (NK)T cells are further distinguished into invariant NKT (iNKT) cells expressing a semi-invariant TCR and non-invariant NKT cells with a more diverse TCR repertoire (see further below).
CD1 family members are considered atypical MHC class I proteins and share selected structural and functional characteristics of classical MHC class I and class II proteins. Thus, similar to MHC class I, CD1 proteins contain three extracellular domains (α1, α2, and α3), a transmembrane domain, and a C-terminal intracellular domain and are synthesized in the endoplasmic reticulum (ER) in a manner dependent on the ER chaperones calreticulin and calnexin as well as the thiol oxireductase ERp57 (Kang and Cresswell, 2002; Cohen et al., 2009). However, differences in the biosynthesis of CD1 and MHC class I exist with regard to the sequence of these interactions and their physiological implications (Kang and Cresswell, 2002). Thus, in contrast to MHC class I, calreticulin, calnexin, and ERp57 interact with the CD1d heavy chain before associating with β2-microglobulin (β2m) (Kang and Cresswell, 2002). As a consequence, the binding of β2m is less critical for folding of CD1d compared with MHC class I, which may explain the occurrence of functionally competent β2m-independent CD1d (Balk et al., 1994; Koh et al., 2008). CD1 isoform-specific differences also exist in that ER exit of CD1b but not CD1d is β2m-dependent (Balk et al., 1994; Sugita et al., 1997).
In the ER, CD1 loads cellular lipids common in this compartment such as glycerophospholipids (GPLs), including phosphatidylcholine (PC) and phosphatidylinositol (PI) (Park et al., 2004; Yuan et al., 2009). Although these findings were obtained with ER-retained forms of CD1d, the association of PC with secreted forms of CD1b and CD1c suggests that similar mechanisms likely apply to group 1 CD1 (Garcia-Alles et al., 2006; Haig et al., 2011). Loading of lipids onto CD1d in the ER is facilitated by lipid transfer molecules expressed in this compartment such as microsomal triglyceride transfer protein (MTP) and may contribute to ligand-induced stabilization of CD1 during biosynthesis (Brozovic et al., 2004; Dougan et al., 2005, 2007; Kaser et al., 2008; Odyniec et al., 2010; Zeissig et al., 2010). This concept is supported by the observation that mutations in the gene encoding for MTP (MTTP), as found in human abetalipoproteinemia (ABL), are associated with proteasomal degradation of CD1a, CD1b, and CD1c (Zeissig et al., 2010).
Following biosynthesis in the ER, CD1 traffics through the secretory pathway to the plasma membrane, where acquired lipids are presented to lipid-reactive T cells. Interestingly, the repertoire of lipids associated with engineered, ER-retained CD1d (ER-CD1d) differs from that observed with soluble CD1d (sCD1d), which lacks the transmembrane domain and is released at the cell surface following completion of trafficking through the secretory pathway. Thus, GPLs were found to be the dominant group of lipids associated with ER-CD1d, whereas both sphingolipids and GPLs were bound to sCD1d (Cox et al., 2009; Yuan et al., 2009; Muindi et al., 2010). This indicates that either unloaded CD1 proteins proceed to the Golgi complex, where they bind sphingolipids generated in this compartment or, more likely, that lipids loaded in the ER are replaced in the Golgi either spontaneously or facilitated by lipid transfer and editing proteins yet to be identified.
In addition to pathways shared with MHC class I, CD1 trafficking patterns partially overlap with those of MHC class II. Thus, CD1 is internalized after reaching the plasma membrane and traffics through the endolysosomal pathway in a manner dependent on the CD1 isoform. CD1a, CD1c, and human CD1d are recruited to early endosomes and the endocytic recycling compartment, whereas CD1b and mCD1d show additional trafficking into lysosomes (Cohen et al., 2009). CD1 isoform-specific trafficking is thereby determined by interactions of the cytoplasmic tail of CD1 with adaptor protein (AP) 2 and 3 leading to internalization through interaction with AP-2 (all CD1 isoforms except for CD1a) and subsequent lysosomal recruitment through interaction with AP-3 (CD1b and mCD1d) (Sugita et al., 1996, 1999, 2000; Chiu et al., 1999; Briken et al., 2000; Jayawardena-Wolf et al., 2001; Chiu et al., 2002). As a consequence of differences in trafficking patterns, CD1 isoforms survey different endolysosomal compartments, where cellular GPLs and sphingolipids loaded in the secretory pathway are exchanged against other endogenous (self) or exogenous (microbial, nutritional) lipids presented to CD1-restricted T cells after recycling of CD1 to the cell surface.
Endolysosomal lipid exchange is facilitated by low pH, lipid transfer and editing proteins, and enzymes involved in lipid processing and catabolism. Acidification within the endolysosomal system ensures optimal activity of hydrolytic enzymes, stabilizes CD1 in the absence of bound lipid (Odyniec et al., 2010), and facilitates lipid loading through direct effects on the conformation of CD1 (Ernst et al., 1998; Relloso et al., 2008). In addition, lipid transfer and editing proteins such as saposin family members and GM2 activator contribute to CD1 lipid loading in the endolysosomal system through extraction of lipids from membranes and transfer onto CD1 as well as potentially through prevention of an irreversible collapse of the CD1 binding cavity in the lipid-free intermediate state (Kang and Cresswell, 2004; Winau et al., 2004; Zhou et al., 2004a,b; Yuan et al., 2007; Garzon et al., 2013). In addition, cathepsin L expression in thymocytes is required for the activation and positive selection of iNKT cells and may act through the assistance in the conversion of prosaposin into saposins (Honey et al., 2002). Further, endolysosomal enzymes involved in the processing of glycolipid headgroups as well as the lipid tails play major roles in the generation of antigenic CD1 lipids. Thus, glycosidases including α- and β-galactosidase, α-mannosidase, and β-hexosaminidase are involved in the trimming of the headgroups of complex glycosphingolipids (GSLs) and bacterial glycosylphosphoinositides, which facilitates lipid headgroup recognition by the NKT-TCR (Sieling et al., 1995; Prigozy et al., 2001; Zhou et al., 2004a,b; de la Salle et al., 2005; Gadola et al., 2006). Deficiency in lysosomal glycolipid-processing enzymes is associated with storage disorders in humans and mice (Kolter and Sandhoff, 2010) and affects lipid antigen presentation through direct effects on the processing of CD1-restricted lipid antigens and indirect effects through interference with lysosomal function (Sieling et al., 1995; Prigozy et al., 2001; Zhou et al., 2004a,b; de la Salle et al., 2005; Gadola et al., 2006). In addition to glycosidases, CD1e, the sole known group 3 CD1 member, facilitates α-mannosidase-dependent processing of glycolipids and directly transfers diacylated phosphatidylinositol mannoside onto CD1b (de la Salle et al., 2005; Facciotti et al., 2011; Garcia-Alles et al., 2011; Cala-De Paepe et al., 2012). Finally, the processing of lipid tails, for example, by phospholipase A2 (PLA2) enzymes, occurs in various subcellular compartments including the endolysosomal system and modulates the antigenicity of CD1 lipids (Fox et al., 2009; Zeissig et al., 2012; Paduraru et al., 2013).
Together, broad and isoform-dependent trafficking of CD1 as well as the concerted action of lipid transfer and processing enzymes facilitate a survey of subcellular compartments for CD1 binding self-lipids and foreign lipids.
CD1 binds a broad spectrum of self-lipids
CD1 isoforms have two or four hydrophobic pockets (A′, C′, F′, and T′), which accommodate the hydrophobic tail of lipid ligands, whereas the lipid headgroup is presented to the NKT-TCR (Moody et al., 2005; De Libero et al., 2009; Girardi and Zajonc, 2012; Rossjohn et al., 2012). Volumes of the binding grooves vary significantly between CD1 isoforms with CD1a at the lower end (1200 Å3) and CD1b at the upper end of the spectrum (2400 Å3) (Moody et al., 2005; Cohen et al., 2009; De Libero et al., 2009). Consequently, CD1b has been shown to capture exogenous lipids with combined alkyl chain lengths of 70–86 carbons, whereas the total lipid tail bound by other CD1 isoforms is in the range of 36–46 carbons (Moody et al., 2005; Cohen et al., 2009). Lipids with up to four alkyl chains were found to be associated with CD1.
In accordance with few restrictions for CD1 binding, a large spectrum of self- and foreign lipids associates with CD1. Lipidomics studies of endogenous lipids associated with individual CD1 members revealed several hundred lipids with distinct mass-to-charge ratios bound to each CD1 isoform in a single cell line in vitro (Cox et al., 2009; Huang et al., 2011). Of those, 5–25% of lipids were CD1 isoform-specific (Huang et al., 2011).
With regard to CD1d, the most extensively studied CD1 member, the spectrum of associated lipids reflected that of the total cellular or compartmental abundance of lipids. Thus, GPLs and sphingolipids were found to be the major groups of lipids associated with CD1d (Cox et al., 2009; Yuan et al., 2009; Muindi et al., 2010; Haig et al., 2011). Within GPLs, abundant cellular lipids such as PC and phosphatidylethanolamine (PE) as well as less abundant lipids such as phosphatidylserine, PI, phosphatidylglycerol, and phosphatidic acid were bound to CD1d (Park et al., 2004; Cox et al., 2009; Shiratsuchi et al., 2009; Yuan et al., 2009; Haig et al., 2011). Among sphingolipids, both sphingomyelin and GSLs were found to be associated with CD1d (Cox et al., 2009; Yuan et al., 2009; Muindi et al., 2010; Haig et al., 2011). A detailed characterization of cellular and CD1d-bound GSLs revealed that the relative abundance of GSLs associated with CD1d did not solely reflect their cellular abundance (Muindi et al., 2010). This suggests that the subcellular localization of CD1d and potential lipid ligands as well as the abundance and activity of lipid processing and transfer proteins may contribute to the repertoire of CD1d-associated lipids. In accordance with the concept of compartmental effects on the CD1d lipidome, the spectrum of GPLs and sphingolipids associated with CD1d differed between CD1d molecules engineered to selectively traffic through the secretory pathway compared with those that exhibited additional endolysosomal trafficking (Yuan et al., 2009; Muindi et al., 2010). Thus, lysophospholipids were predominantly associated with CD1d proteins, which maintain the ability to undergo endolysosomal trafficking, whereas the ganglioside GM2 was only detected in CD1d engineered to selectively survey secretory compartments (Yuan et al., 2009; Muindi et al., 2010).
Similar studies on endogenous ligands have recently been performed for secreted forms of group 1 CD1, which selectively traffic through the secretory but not the endolysosomal pathway due to lack of the CD1 transmembrane domain and the intracellular C-terminus (Haig et al., 2011; Huang et al., 2011). These studies revealed a large overlap in the spectrum of lipids associated with group 1 and group 2 CD1 with some noteworthy isoform-specific distinctions. Thus, CD1b, despite having a significantly larger lipid binding groove than other CD1 isoforms, captures low-mass lipids such as diacylglycerols and deoxyceramides (Facciotti et al., 2011; Huang et al., 2011). These lipids act as scaffolds and facilitate subsequent parallel binding of lipids that would otherwise not fully occupy the CD1b-binding groove, thus contributing to optimal positioning of these lipids for T-cell recognition (Garcia-Alles et al., 2006; Facciotti et al., 2011; Huang et al., 2011). In addition, these and other lipids may act as spaceholders for subsequent exchange against lipids with long acyl chains, which fully occupy the CD1b groove (Huang et al., 2011).
Antigenicity is limited to a small subset of self-lipids
Group 1 CD1 expression is limited to professional antigen-presenting cells (APCs) including, in case of CD1c, a subset of B cells (Cohen et al., 2009). In contrast, CD1d is broadly expressed by professional and non-professional APCs including epithelial and endothelial cells (Blumberg et al., 1991; Geissmann et al., 2005; Cohen et al., 2009). As CD1d binds to a wide variety of self-lipids, it may provide a constant source of antigens for CD1d-restricted (NK)T cells, both in the thymus and the periphery. In accordance with this concept, autoreactivity, i.e., CD1-dependent activation by APCs in the absence of exogenous antigen, is a hallmark of (NK)T cells. However, the search for self-lipid antigens involved in (NK)T cell activation revealed that the majority of CD1d-binding lipids exhibit little or no antigenicity (Fox et al., 2009; Brennan et al., 2011; Pei et al., 2011; Facciotti et al., 2012; Zeissig et al., 2012). Noteworthy examples of antigenic CD1d self-lipids that activate iNKT cells include GSLs such as isoglobotrihexosylceramide (iGb3), β-glucosylceramide (β-GlcCer), β-galactosylceramide (β-GalCer), and lysosphingomyelin as well as phospholipids such as lysophosphatidylcholine (LPC), ether-bonded lysophosphatidylethanolamine (pLPE) and lysophosphatidic acid (eLPA) (Figure 1) (Zhou et al., 2004a,b; Fox et al., 2009; Brennan et al., 2011; Facciotti et al., 2012). In addition, CD1d self-lipids that exhibit antigenicity for non-invariant NKT cells include ester-bonded lysophosphatidylethanolamine (LPE) and the GSLs sulfatide, β-GlcCer and β-GalCer, whereby the lyso-derivatives of the GSLs showed increased antigenicity (Figure 1) (Jahng et al., 2004; Roy et al., 2008; Brennan et al., 2011; Rhost et al., 2012; Zeissig et al., 2012). The relative contribution of these antigenic lipids to (NK)T cell-positive selection in the thymus and activation in the periphery as well as the mechanisms underlying species-specific differences in self-antigen-mediated (NK)T cell activation are currently not clear and require further analysis. Similarly, the mechanisms that prevent autoimmunity in the presence of antigenic self-lipids require further study but may include the balance of antigenic and non-antigenic CD1d lipids.
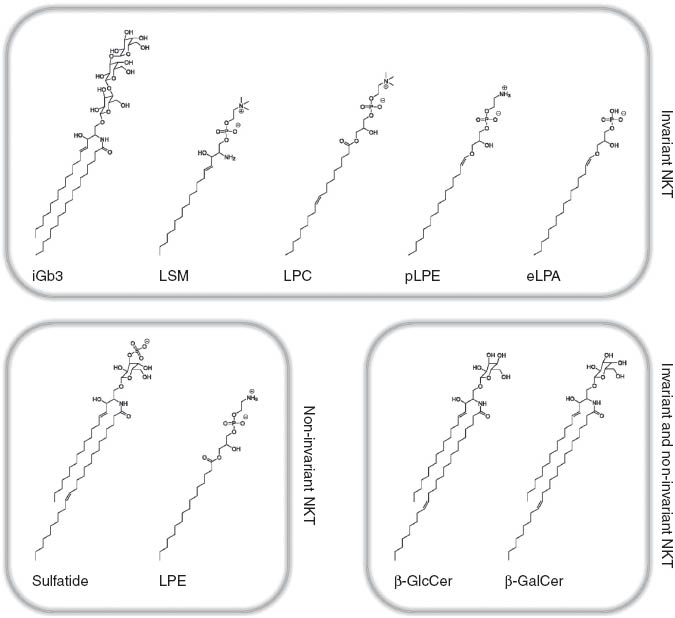
Self-antigens involved in the activation of invariant and non-invariant NKT cells.
Mechanistically, the discrepancy between the number of CD1d-binding lipids and those associated with (NK)T cell antigenicity is the consequence of a rather non-selective binding mode based on hydrophobic interactions between CD1d and the aliphatic lipid tail, whereas antigenicity follows more strict, but only partially understood structural requirements that allow for recognition of the CD1d-lipid complex by the NKT-TCR (De Libero et al., 2009; Girardi and Zajonc, 2012; Rossjohn et al., 2012). These structure-function relationships at the interface of the CD1d-lipid complex and the NKT-TCR have recently been the focus of excellent reviews and will only be briefly outlined here (De Libero et al., 2009; Girardi and Zajonc, 2012; Rossjohn et al., 2012). Thus, recognition of the CD1d-lipid complex by the NKT-TCR is based on interactions between the TCR and CD1d as well as the headgroup of lipids. Thereby, the molecular nature of the headgroup, its size, and its linkage to the lipid backbone affect TCR recognition (De Libero et al., 2009; Girardi and Zajonc, 2012; Rossjohn et al., 2012). Moreover, the length, composition, and saturation of lipid tails modulate positioning of lipids within CD1, thus regulating TCR recognition (De Libero et al., 2009; Girardi and Zajonc, 2012; Rossjohn et al., 2012). Remarkably, the spectrum of antigenic headgroups extends from more (iGb3) or less complex (β-GlcCer) carbohydrates and phospholipid headgroups (LPC and LPE) to the minimal sugar-free phosphate anion headgroup of eLPA (Zhou et al., 2004a,b; Brennan et al., 2011; Facciotti et al., 2012; Ly et al., 2013). In case of GSLs, mammalian β-glycosidic linkage confers lower TCR affinity compared with microbial α-glycosidic linkage because it requires an induced fit of the NKT-TCR (Pellicci et al., 2011; Yu et al., 2011). Similarly, complex GSL headgroups may require enzymatic processing and trimming for TCR recognition, whereas some adaptation to bulkier hexose headgroups such as that of iGb3 can also be achieved by an induced TCR fit (Florence et al., 2009). In contrast, the mechanisms that confer specificity to NKT-TCR recognition of some but not other phospholipids and the mechanistic basis of the observed increased potency of lyso-variants of phospholipids and sphingolipids are just beginning to emerge (Lopez-Sagaseta et al., 2012).
Finally, recent technical advances including group 1 CD1 tetramers and lipidome analysis of engineered group 1 CD1 proteins provide the basis for ongoing studies of the CD1a-, CD1b-, and CD1c-associated lipidome and the mechanisms that govern recognition by group 1 CD1 TCRs (Huang et al., 2011; Kasmar et al., 2011; Ly et al., 2013; Van Rhijn et al., 2013). These studies have revealed a significant overlap in the spectrum of lipids associated with group 1 and group 2 CD1 (Huang et al., 2011). Moreover, commonalities in self-lipid recognition by group 1 and 2 CD1-restricted (NK)T cells are likely to exist, given the recognition of particular self-antigens such as sulfatide across group 1 and group 2 CD1-restricted (NK)T cells (Shamshiev et al., 2002; Jahng et al., 2004). However, it is conceivable that the spectrum of antigenic lipids associated with group 1 CD1 exceeds that of CD1d given the diversity in the TCR repertoire of group 1 CD1-restricted T cells (Grant et al., 1999).
Environmental influences shape the CD1 self-lipid repertoire
Recent studies have revealed that the repertoire of CD1 self-lipids is not static but rather subject to continuous regulation in response to microbial, inflammatory, and likely metabolic stimuli. Such alterations may act in concert with microbiota- and inflammation-induced maturation of APCs, upregulation of CD1 expression, and CD1-dependent and CD1-independent (NK)T cell activation to contribute to CD1- and (NK)T cell-dependent immunity. Moreover, metabolic factors such as serum lipids and lipoproteins are involved in the regulation of CD1 expression and further contribute to the control of CD1-dependent immunity (Gogolak et al., 2007; Leslie et al., 2008).
Microbial infection and exposure to toll-like receptor (TLR) agonists affects cellular lipid metabolism and leads to alterations in the spectrum and antigenicity of CD1 lipids, particularly through the regulation of sphingolipid metabolism (De Libero et al., 2005; Paget et al., 2007; Salio et al., 2007; Muindi et al., 2010; Brennan et al., 2011). Recent studies have provided a comprehensive characterization of microbial-induced alterations in the sphingolipid self-antigen repertoire associated with CD1d and revealed β-GlcCer as a potent self-antigen for (NK)T cells, which is induced upon bacterial infection or TLR stimulation (Muindi et al., 2010; Brennan et al., 2011). In addition, it has recently been demonstrated that microbial-induced alterations in the endogenous CD1d lipid repertoire are not limited to bacterial infection. Thus, hepatitis B virus (HBV) infection is, among other broad alterations in the hepatocyte lipid profile, associated with increased abundance of PE. PE binds to CD1d but is non-antigenic and requires HBV-dependent induction of selected secretory PLA2 enzymes for hydrolysis into LPE (Figure 1), which is strongly antigenic for a subgroup of non-invariant NKT cells (Zeissig et al., 2012). Noteworthy, alterations in the abundance of several antigenic CD1d lipids, including β-GlcCer and lysophospholipids, are not restricted to specific microbial infections but represent common signatures of inflammatory and malignant diseases (Fuchs et al., 2012; Santos and Schulze, 2012). It is therefore likely that the CD1 self-lipid repertoire is under continuous metabolic and immune-mediated control, with broad implications for (NK)T cell-dependent immunity at steady state and during inflammatory, malignant, and metabolic disorders.
Microbial organisms contain potent CD1 lipid antigens
In addition to shaping the repertoire of CD1 self-lipids, environmental and particularly microbial exposure provides a wealth of exogenous lipid antigens, which contributes to antimicrobial responses of lipid-reactive T- and (NK)T cells in the context of local or systemic infections (Kinjo and Kronenberg, 2009; Brennan et al., 2013).
The structural complexity of microbial-derived CD1 lipid antigens considerably exceeds that of self-lipid antigens. Thus, seminal studies by Brenner, Porcelli, Moody, and others revealed that group 1 CD1 associates with a wide variety of mycobacterial phospholipids, glycolipids, mycolic acids, lipopeptides, mycoketides, and isoprenoids, which act as antigens for group 1 CD1-restricted T cells (Moody et al., 2005; Cohen et al., 2009). Similarly, various group 2 CD1 (CD1d)-binding, (NK)T cell-activating lipids have been described and include α-linked GSLs derived from Sphingomonas species (e.g., GSL-1; Figure 2) (Kinjo et al., 2005; Mattner et al., 2005; Sriram et al., 2005), diacylglycerol-containing glycolipids derived from Borrelia burgdorferi (e.g., Bb-GLIIc; Figure 2) (Kinjo et al., 2006), Streptococcus pneumoniae and group B Streptococcus (Kinjo et al., 2011), phospholipids derived from mycobacteria and Corynebacterium spp. [e.g., Corynebacterium glutamicum diphosphatidylglycerol (Cg-DPG); Figure 2] (Fischer et al., 2004; Tatituri et al., 2013), a cholesterol ester produced by Helicobacter pylori (PI-57; Figure 2) (Chang et al., 2011), and lipophosphoglycans derived from Leishmania donovani (Amprey et al., 2004), and Entamoeba histolytica (Lotter et al., 2009).
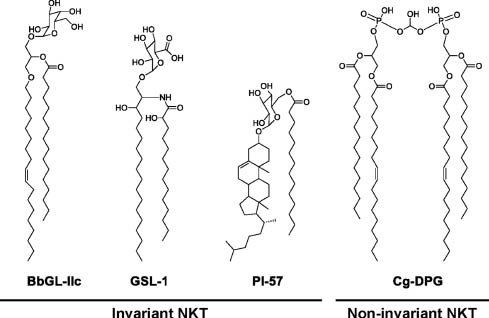
Selected microbial lipid antigens that activate invariant and non-invariant NKT cells.
Exogenous antigens typically associate with a particular CD1 isoform and load onto CD1 either at the plasma membrane or in the endolysosomal pathway. The specificity of lipid binding to a particular CD1 isoform relates both to the structural requirements of these interactions and differences in the compartmental distribution of CD1 isoforms and lipids. Thus, CD1a contains a shallow and laterally oriented F′ pocket, which, for example, facilitates binding and presentation of the peptide moiety of the lipopeptide didehydroxymycobactin (Moody et al., 2004; Zajonc et al., 2005). In contrast, binding of mycobacteria-associated long-chain mycolic acids to CD1b is facilitated by the large antigen binding groove of CD1b and specific trafficking of these lipids to late endosomes, where CD1b is located and where loading is facilitated by acidic pH (Moody et al., 2002; Relloso et al., 2008). In case of mycobacterial mannosyl phosphomycoketide, specific binding to CD1c is conferred by methyl branches found in foreign mycoketides but not mammalian self-lipids (Scharf et al., 2010; Ly et al., 2013). These examples of specific interactions between microbial lipids and CD1 illustrate the unique ability of CD1 to survey different subcellular compartments for exogenous, microbial lipid antigens in a CD1 isoform-dependent manner.
In the context of an infection, antigen-dependent and antigen-independent pathways contribute to the activation of lipid-reactive T cells and particularly CD1d-restricted (NK)T cells (see below). For these reasons, the contribution of an individual microbial-derived CD1 lipid antigen to immune-mediated control of a particular infection is often difficult to assess. However, studies using group 1 and group 2 CD1 tetramers loaded with microbial lipids could demonstrate the presence of (NK)T cells reactive or cross-reactive to these lipids in humans and mice (Fischer et al., 2004; Kinjo et al., 2005; Mattner et al., 2005; Sriram et al., 2005; Kinjo et al., 2006; Kasmar et al., 2011; Ly et al., 2013; Van Rhijn et al., 2013). Moreover, expansion of mycobacterial lipid-reactive group 1 CD1-restricted T cells in humans and mice following M. tuberculosis infection suggests that these T cells and the lipid antigens recognized participate in the antimicrobial immune response in vivo (Felio et al., 2009; Van Rhijn et al., 2013). Future studies in animal models will be critical to delineate the contribution of individual lipid antigens to antimicrobial immune responses through genetic and chemical manipulation of metabolic pathways associated with the generation of these lipids.
Lipid-reactive T cells
Endogenous and exogenous lipids presented by CD1 activate subsets of lipid-reactive T cells in a TCR-restricted manner. Thus, group 1 CD1 presents lipid antigens to CD1a-, CD1b-, and CD1c-restricted T cells, whereas group 2 CD1 presents lipids to CD1d-restricted (NK)T cells (Porcelli et al., 1989, 1992; Balk et al., 1991; Blumberg et al., 1991; Beckman et al., 1994; Kawano et al., 1997).
CD1d-restricted (NK)T cells
CD1d-restricted (NK)T cells are distinguished, based on their TCR expression, into invariant and non-invariant NKT cells, which differ with regard to the antigens recognized and the functions exerted. Invariant, or type I, (NK)T cells are characterized by the expression of a semi-invariant TCR composed of Vα14-Jα18 in mice and Vα24-Jα18 in humans, paired with a limited set of Vβ chains (Bendelac et al., 2007). iNKT cells recognize the marine sponge glycolipid α-GalCer and can be directly detected using CD1d-tetramers loaded with this lipid (Matsuda et al., 2000), which has provided unique insight into the phenotype and function of this (NK)T cell subset. iNKT cells exhibit co-expression of T-cell and NK-cell markers and are characterized by an effector memory phenotype (Bendelac et al., 2007). In accordance with these phenotypic characteristics, iNKT cells exhibit rapid, innate-like activation and secrete abundant amounts of TH1 [interferon (IFN) γ], TH2 [interleukin (IL) 4, 13, and 10], and TH17 cytokines (IL-17) with broad effects on other innate and adaptive immune cells and central roles in antimicrobial and cancer immunity as well as the regulation of autoimmunity and metabolism (Bendelac et al., 2007; Brennan et al., 2013).
Non-invariant, or type II, NKT cells are also CD1d-restricted but do not express the semi-invariant TCR. Compared with iNKT cells, less is known about non-invariant NKT cells due to the lack of specific markers for these cells. Initial studies on non-invariant NKT cells in MHC class II-deficient mice as well as subsequent studies on a subset of non-invariant NKT cells that recognize sulfatide revealed an oligoclonal TCR repertoire with predominant usage of Vβ8 paired with a limited number of different Vα chains (Vα8, Vα3, and Vα1) (Cardell et al., 1995; Park et al., 2001; Arrenberg et al., 2010). In addition to differences in the TCR repertoire between invariant and non-invariant NKT cells, the antigens recognized by these cells (see above), the mode of their recognition, and the phenotype and function of (NK)T cell subsets are distinct. Thus, the ternary complex of CD1d, sulfatide, and the non-invariant NKT-TCR revealed fundamental differences in interaction compared with the iNKT-CD1d complex (Girardi et al., 2012; Patel et al., 2012). Specifically, the type I NKT-TCR adopts a tilted and parallel docking mode over the F′-pocket of CD1d, whereas the type II NKT-TCR docked diagonally above the A′-pocket of CD1d (Girardi et al., 2012; Patel et al., 2012). Functionally, non-invariant NKT cells lack constitutive expression of the early activation marker CD69 and exhibit regulatory actions in part through suppression of iNKT cell activation (Terabe et al., 2000; Jahng et al., 2004; Halder et al., 2007; Berzofsky and Terabe, 2009; Arrenberg et al., 2010). However, LPE-reactive non-invariant NKT cells can support antimicrobial immunity both directly via IFN-γ secretion and indirectly via activation of iNKT cells and conventional T cells, suggesting the presence of functional heterogeneity even within the non-invariant NKT cell subset (Zeissig et al., 2012). Finally, additional complexity is provided by the recent discovery of atypical (NK)T cell subsets, which are α-GalCer/CD1d-tetramer-reactive, but do not express murine Vα14 or human Vα24 (Constantinides et al., 2011; Uldrich et al., 2011).
Group 1 CD1-restricted T cells
In contrast to (NK)T cells, group 1 CD1-restricted T cells do not exist in mice due to the lack of CD1a, CD1b, and CD1c. For this reason, functional studies of group 1 CD1-restricted T cells were initially limited to the analysis of T cell clones expanded from human peripheral blood. These studies revealed that group 1 CD1-restricted T cells express either αβ- or γδ-TCRs, can be grouped based on co-receptor expression into CD4+, CD8+, or double-negative T cells, and consistently secrete IFN-γ upon activation, whereas secretion of TH2 and TH17 cytokines exhibits significant CD1 isoform-dependent and inter-individual variation (Porcelli et al., 1989; Vincent et al., 2005; de Jong et al., 2010; de Lalla et al., 2011). Further, these cells resemble conventional MHC class I- and II-restricted T cells rather than iNKT cells in that they express a diverse TCR Vα and Vβ repertoire and, at least in peripheral blood, express markers characteristic of naive T cells (Grant et al., 1999; Vincent et al., 2005; de Jong et al., 2010; de Lalla et al., 2011). Similar to conventional MHC class I- and II-restricted T cells, the ratio of memory to naive group 1 CD1-restricted T cells increases from birth to adulthood with an increased percentage of cells expressing memory markers in peripheral tissues, suggesting antigen-dependent, activation-induced transition from naive to memory phenotype (de Jong et al., 2010; de Lalla et al., 2011).
Although studies of group 1 CD1-restricted T cell clones provided critical insight into the mechanisms of TCR-dependent recognition of foreign lipid antigens, it remained unclear whether the phenotype, function, and frequency of group 1 CD1-restricted T cells within these clonal populations reflected the situation found in vivo. The development of group 1 CD1 tetramers therefore represented a major breakthrough in the field because it provided the opportunity to directly detect group 1 CD1-restricted T cells among peripheral blood mononuclear cells (Kasmar et al., 2011; Ly et al., 2013; Van Rhijn et al., 2013). These studies revealed that T cells recognizing mycobacterial lipids in the context of CD1b and CD1c can be detected among polyclonal T cells in peripheral blood, whereas CD1b-restricted autoreactivity is not observed in contrast to CD1a (de Jong et al., 2010; de Lalla et al., 2011; Van Rhijn et al., 2013). Moreover, this work led to the recognition of an invariant CD1b-restricted germline-encoded, mycolyl lipid-reactive (GEM) T cell subset expressing a semi-invariant TCR composed of TRAV1-2 and TRAJ9 paired with a limited set of Vβ chains (Van Rhijn et al., 2013). Despite the expression of a semi-invariant TCR, GEM T cells are phenotypically and functionally distinct from iNKT cells in that they do not consistently express the NK-cell marker CD161, lack baseline expression of the activation marker CD69, and are increased in numbers in patients with tuberculosis in accordance with infection-induced expansion of GEM T cells (Van Rhijn et al., 2013). In addition, recently developed group 1 CD1 transgenic mice confirmed that group 1-restricted T cells resemble conventional T cells rather than iNKT cells and exhibit delayed immune responses and infection-induced expansion (Felio et al., 2009).
In conclusion, pioneering studies of lipid-reactive T cell clones and the recent development of group 1 CD1 tetramers and transgenic mice (see below) provided critical insight into the biology of CD1a-, CD1b-, and CD1c-restricted T cells. This work revealed that group 1 CD1-restricted T cells phenotypically and functionally resemble conventional T cells rather than iNKT cells and recognize a wide variety of mycobacterial lipid antigens. Future studies using tetramers and animal models will be important to obtain further insight into the frequency, TCR repertoire, and function of CD1a-, CD1b-, and CD1c-restricted T cells in vivo.
Antigen-dependent and antigen-independent pathways to activation of CD1-restricted (NK)T cells
Antigen-dependent and antigen-independent pathways to CD1-restricted, (NK)T cell-dependent immunity have been described (Figure 3). As such, a large number of microbial lipids can bind to CD1 and activate CD1-restricted T cells in vitro and in vivo, thus providing the basis for antigen-driven, CD1-restricted antimicrobial immune responses (Figure 3, left). In addition, CD1 expression can be modulated upon infection, which can either facilitate antigen presentation through upregulation of CD1 or contribute to immune evasion through interference with this process (Amprey et al., 2004; Shinya et al., 2004; Berntman et al., 2005; Cho et al., 2005; Hage et al., 2005; Lin et al., 2005; Renukaradhya et al., 2005; Sanchez et al., 2005; Skold et al., 2005; Chen et al., 2006; Raghuraman et al., 2006; Yuan et al., 2006; Donovan et al., 2007; Kawana et al., 2007; Raftery et al., 2008).
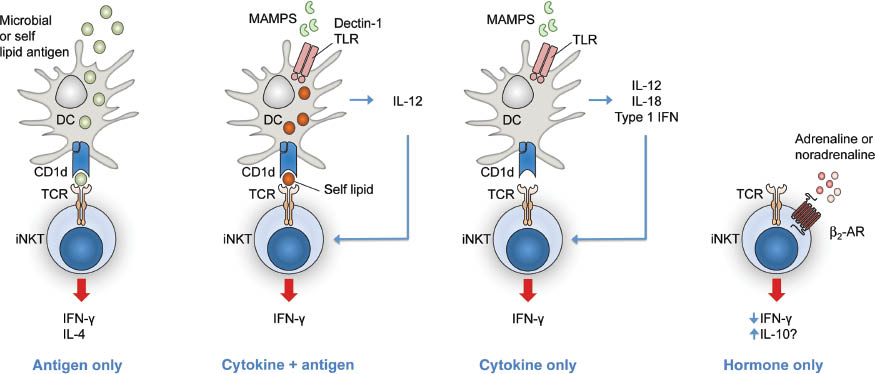
Direct and indirect mechanisms of iNKT cell activation or modulation.
β2-AR, β2-adrenergic receptor; DC, dendritic cell.
Moreover, microbial exposure can affect the CD1 self-lipid repertoire and its antigenicity, thereby contributing to protective antimicrobial immunity through (NK)T cell activation (De Libero et al., 2005; Paget et al., 2007; Salio et al., 2007; Muindi et al., 2010; Brennan et al., 2011; Zeissig et al., 2012).
In addition to direct, antigen-dependent activation of lipid-reactive T cells, indirect cytokine-driven mechanisms exist, which promote the activation of CD1d-restricted iNKT cells independent of microbial lipid antigens. Thus, recognition of microbe-associated microbial patterns (MAMPs) through pattern recognition receptors such as TLRs and dectin 1 on dendritic cells (DCs) is associated with production of TLR4-dependent IL-12 and IL-18, dectin 1-dependent IL-12, and TLR9-dependent IL-12 and type I IFN. These cytokines potently activate iNKT cells, which is further enhanced by CD1d-restricted presentation of self-lipid antigens (Figure 3, second from left) (Brigl et al., 2003, 2011; Mattner et al., 2005; Montoya et al., 2006; Brennan et al., 2011; Cohen et al., 2011). Depending on the context, cytokines can be sufficient for indirect stimulation of iNKT cells, even in the absence of CD1d signals (Figure 3, second from right) (Nagarajan and Kronenberg, 2007; Tyznik et al., 2008). In addition to these mechanisms, TLR engagement and APC- and iNKT cell-dependent cytokine secretion promote CD1d-restricted antigen presentation through differentiation and maturation of DCs (Fujii et al., 2003; Hermans et al., 2003, 2007; Hegde et al., 2007). Cytokine-mediated iNKT cell activation thus enables iNKT cell-dependent recognition of bacteria and viruses devoid of CD1d-restricted lipid antigens and provides a mechanistic explanation for broad, iNKT cell-dependent immune responses against a wide variety of bacterial, viral, fungal, and protozoan pathogens. Moreover, lipid antigen- and cytokine-dependent pathways of iNKT cell activation are not exclusive but act together in iNKT cell-mediated antimicrobial immune responses in vivo. Thus, even in the case of infection with microbes containing antigenic CD1d lipids, cytokine-mediated stimulation of iNKT cells critically contributes to the overall immune response (Brigl et al., 2003, 2011). Finally, neurotransmitters may provide a third pathway to iNKT cell activation in addition to antigen- and cytokine-dependent mechanisms (Figure 3, right) (Wong et al., 2011).
In conclusion, the activation of lipid-reactive (NK)T cells in infection is facilitated by regulation of CD1 expression, alterations in the CD1 self-lipid profile, modulation of APC function, and antigen- and cytokine-dependent iNKT cell activation.
Murine studies of lipid-reactive T cells in immunity, cancer, and metabolism
Our knowledge of the physiological and pathophysiological roles of lipid-reactive T cells is largely based on the study of CD1d-restricted (NK)T cells in mice. These studies revealed that (NK)T cells play critical roles in antimicrobial immunity, cancer immunosurveillance, the modulation of immune-mediated or autoimmune diseases, and the regulation of lipid and carbohydrate metabolism. Selective aspects of the functions of (NK)T cells and group 1 CD1-restricted T cells in immunity are discussed in the following sections. A more comprehensive description of this topic has been provided by several excellent, recent reviews (Berzofsky and Terabe, 2009; Cohen et al., 2009; Kinjo and Kronenberg, 2009; Vivier et al., 2012; Brennan et al., 2013).
(NK)T cells in antimicrobial immunity
Protective as well as pathogenic roles of (NK)T cells have been described in the immune response to a large number of bacterial, viral, fungal, and protozoan infections and in a subset of cases, direct, antigen-driven and indirect, cytokine-mediated (NK)T cell activation could be delineated (Tupin et al., 2007; Cohen et al., 2009). Thus, microbial-derived, CD1d-restricted lipid antigens implicated in antimicrobial immunity were found in Sphingomonas spp. (Kinjo et al., 2005; Mattner et al., 2005; Sriram et al., 2005), B. burgdorferi (Kinjo et al., 2006), S. pneumoniae, group B Streptococcus (Kinjo et al., 2011), Mycobacterium tuberculosis (Fischer et al., 2004; Tatituri et al., 2013), H. pylori (Chang et al., 2011), L. donovani (Amprey et al., 2004), and E. histolytica (Lotter et al., 2009). In contrast, TLR-induced cytokine-driven responses mediate iNKT cell activation in response to bacteria and viruses devoid of lipid antigens and also contribute to immunity against bacteria containing CD1d-restricted lipid antigens (Brigl et al., 2003, 2011; Mattner et al., 2005; Montoya et al., 2006; Nagarajan and Kronenberg, 2007; Tyznik et al., 2008). Another example of combined direct and indirect effects on (NK)T cell activation is HBV infection, where early HBV-induced alterations in the self-lipid repertoire are recognized by a subset of non-invariant NKT cells, which consequently leads to broad, IL-12-dependent activation of other immune cells including iNKT cells (Zeissig et al., 2012).
Although a vast number of studies aimed at the delineation of the roles of invariant and non-invariant NKT cells in antimicrobial immunity, these studies were based on comparative analysis of CD1d- and Jα18-deficient mice and may need reassessment given the recent discovery of unanticipated, broad defects in the TCR repertoire of Jα18 knockout mice (Bedel et al., 2012). CD1d knockout mice (Cd1d1-/-Cd1d2-/-) lack both invariant and non-invariant NKT cells due to loss of CD1d-restricted positive selection in the thymus (Chen et al., 1997; Gapin et al., 2001; Benlagha et al., 2005). In contrast, Jα18 knockout mice exhibit a deletion of the Jα segment of the semi-invariant iNKTCR (Traj18-/-) and were developed to selectively delete iNKT cells (Cui et al., 1997). However, a recent study revealed that Jα18 knockout mice exhibit an unanticipated loss of a significant fraction of Jα regions associated with lack of about 60% of TCRα diversity (Bedel et al., 2012). Impaired immunity in Traj18-/- mice may therefore relate to lack of iNKT cells as well as defects in the conventional T cell repertoire, which limits conclusions derived from the study of these mice. For these reasons, the relative contributions of invariant and non-invariant NKT cells to antimicrobial immunity are largely unclear at present.
Finally, in accordance with the critical role of (NK)T cells in antimicrobial immunity, immune evasion strategies exist, particularly for viral agents. Thus, HIV infection is associated with internalization and trans-Golgi retention of CD1d in a manner dependent on the HIV Nef and gp120 proteins (Cho et al., 2005; Hage et al., 2005; Chen et al., 2006). Moreover, HIV infection is associated with (NK)T cell exhaustion through the modulation of PD-1 expression and PD-1-independent pathways (Moll et al., 2009). Similar observations of evasion of CD1d-restricted immunity have been made for other viruses such as lymphocytic choriomeningitis virus, vesicular stomatitis virus, herpes simplex virus, Kaposi sarcoma-associated herpesvirus and bacteria including Leishmania and Chlamydia species (Lin et al., 2005; Sanchez et al., 2005; Raftery et al., 2006; Yuan et al., 2006; Kawana et al., 2007; Renukaradhya et al., 2008).
(NK)T cells in cancer immunosurveillance
α-GalCer, a marine sponge-derived GSL, was originally identified in a screen for tumor-modulating compounds, where it was associated with protection in the B16 melanoma model (Kobayashi et al., 1995; Motoki et al., 1996). α-GalCer was later demonstrated to act through the activation of iNKT cells and to suppress tumor development in an IFN-γ- and IL-12-dependent manner (Kawano et al., 1997; Kitamura et al., 1999). In addition, related ceramide-based lipids such as β-mannosylceramide elicit similar anti-oncogenic actions but act through independent iNKT cell-mediated pathways dependent on TNF-α and nitric oxide (O’Konek et al., 2011). Even in the absence of exogenous, stimulatory lipid antigens, iNKT cells contribute to natural immunosurveillance in solid and hematopoietic cancers including fibrosarcoma induced by methylcholanthrene (Smyth et al., 2000; Crowe et al., 2002, 2005), adenocarcinomas, sarcomas, and hematopoietic tumors induced by loss of p53 (Swann et al., 2009), colorectal cancer (Yoshioka et al., 2012), hepatocellular carcinoma (Anson et al., 2012), and neuroblastoma (Song et al., 2009). Studies in neuroblastoma also revealed that iNKT cells may not only interfere with tumor growth through direct tumor cell cytotoxicity but can act indirectly through killing of tumor-associated, oncogenic macrophages (Song et al., 2009). Although initial cancer studies were performed in Jα18 knockout mice, with limitations as described above, CD1d-deficient mice exhibited a similar phenotype, and the transfer of total (NK)T cells or purified iNKT cells was associated with protection thus supporting a role of iNKT cells in anticancer immunity (Crowe et al., 2002, 2005; Song et al., 2009; Izhak et al., 2013). One remarkable exception to the rule of anti-oncogenic effects of iNKT cells is the model of azoxymethane/oxazolone-induced colitis-associated colorectal cancer, which is mediated by iNKT cells in a process dependent on IL-13 secretion, induction of CD11bhi Gr1low macrophages, and transforming growth factor-β (TGF-β)-dependent suppression of tumor immunosurveillance (Schiechl et al., 2011).
In contrast to protective roles of iNKT cells, non-invariant NKT cells were suggested to contribute to tumor growth in an oncogenic manner in models of fibrosarcoma as well as mammary, colorectal, and renal cell carcinoma (Terabe et al., 2000, 2003; Izhak et al., 2013). These actions were mediated by IL-13-induced expression of TGF-β by CD11b+ Gr-1+ cells and suppression of tumor-specific CD8+ T cells (Terabe et al., 2000, 2003; Izhak et al., 2013). In the majority of these studies, the protective effects of CD1d deletion were ascribed to the loss of non-invariant NKT cells, as tumor development in Jα18-deficient mice resembled that observed in wildtype mice. Although the defects in the conventional TCR repertoire may have affected tumor cell growth, recent studies based on selective activation of invariant and non-invariant NKT cell subsets and reconstitution of iNKT cells in Jα18-deficient mice revealed similar results and support the concept of opposing roles of NKT cell subsets in cancer (Izhak et al., 2013). Noteworthy, invariant and non-invariant NKT cell subsets cross-regulate each other not only in terms of function but also frequency, thus adding another layer of complexity to the role of (NK)T cells in malignancy (Izhak et al., 2013).
Although the aforementioned studies were performed in mice, indirect evidence suggests similar roles of (NK)T cells in human malignancy. Thus, iNKT cell infiltration in colorectal cancer and neuroblastoma correlates with increased survival (Metelitsa et al., 2004; Tachibana et al., 2005), whereas low levels of circulating iNKT cells predict poor clinical outcomes in head and neck squamous cell carcinoma (Molling et al., 2007). Moreover, iNKT cells obtained from human cancer patients exhibit impaired proliferative responses, decreased secretion of IFN-γ, and an increased ratio of regulatory IL-4 to anti-oncogenic IFN-γ (Tahir et al., 2001; Yanagisawa et al., 2002; Dhodapkar et al., 2003; Bricard et al., 2009).
Given the protective roles of iNKT cells in oncogenesis, therapeutic modulation of (NK)T cells has been investigated in human malignancy. Direct injection of α-GalCer into patients with solid tumors proved largely unsuccessful due to limited iNKT cell responses (Giaccone et al., 2002). Subsequent approaches included the transfer of α-GalCer-pulsed autologous DCs, which was also associated with considerable, inter-individual variation in the response observed (Ishikawa et al., 2005; Motohashi et al., 2006, 2009; Uchida et al., 2008; Nagato et al., 2012). Therefore, combination therapy may be required for efficacious (NK)T cell-based immunotherapy in cancer. In accordance with this concept, a subset of patients with multiple myeloma exhibited tumor regression upon treatment with α-GalCer-loaded dendritic cells in combination with lenalidomide (Richter et al., 2013).
Together, these studies demonstrate a critical role of (NK)T cells in tumor development and warrant further optimization of (NK)T cell-based immunotherapy in human malignancy.
(NK)T cells in the regulation of immune-mediated diseases
(NK)T cells were suggested to contribute to a variety of human immune-mediated diseases and their murine models including type I diabetes, multiple sclerosis, rheumatoid arthritis, systemic lupus erythematosus, autoimmune hepatitis, primary biliary cirrhosis, sarcoidosis, asthma, and inflammatory bowel disease (IBD). Owing to the complexity of this field and the controversies that have arisen (Subleski et al., 2011), we will focus on the role of (NK)T cells in IBD, where recent studies have provided critical insight into the regulation of mucosal immune responses by (NK)T cells at the interface between the host and the environment.
We and others could demonstrate that (NK)T cells play critical roles in human IBD and mouse models of intestinal inflammation (Saubermann et al., 2000; Heller et al., 2002; Fuss et al., 2004; Zeissig et al., 2007; Schiechl et al., 2011; Camelo et al., 2012; Olszak et al., 2012). Thus, oxazolone colitis, a model of human ulcerative colitis (UC), is dependent on CD1d and CD1d-restricted (NK)T cells (Heller et al., 2002; Schiechl et al., 2011; Olszak et al., 2012). Similar findings were made for human UC, where key pathogenic cytokines such as IL-13 originate from (NK)T cells and contribute to impaired epithelial barrier function and sustained intestinal inflammation (Fuss et al., 2004; Heller et al., 2005). Although studies in oxazolone colitis reported a central, pathogenic role of iNKT cells (Heller et al., 2002), this work was based on Jα18-deficient mice and is associated with the respective limitations outlined above (Bedel et al., 2012). In contrast, pathogenic IL-13 in human UC originates from α-GalCer/CD1d-tetramer-negative T cells and thus presumably non-invariant NKT cells (Fuss et al., 2004). In accordance with a predominant role of non-invariant NKT cells in intestinal inflammation, transgenic expression of a non-invariant TCR led to spontaneous intestinal inflammation in mice, particularly in the presence of overexpression of CD1d (Liao et al., 2012). The biological relevance of these findings is highlighted by the fact that genetic polymorphisms associated with susceptibility to IBD relate to genes enriched in expression by NKT cells (Jostins et al., 2012). Further, a variety of genes with critical roles in lipid metabolism have been highlighted by expression analysis and genome-wide association studies in IBD and may at least partially contribute to disease susceptibility through modulation of the activation of lipid-reactive T cells (Jostins et al., 2012; Planell et al., 2013). Finally, a randomized, placebo-controlled, multicenter study recently revealed the efficacy of oral PC in UC (Karner et al., 2012). Given that PC binds to CD1d but does not activate human (NK)T cells (Fox et al., 2009), interference with CD1d-restricted mucosal (NK)T cell activation may contribute to the therapeutic efficacy of PC, a concept that awaits verification.
It has recently been demonstrated that intestinal inflammation in oxazolone colitis is not dependent on microbial antigens as germfree mice exhibited increased rather than decreased intestinal inflammation (Olszak et al., 2012). These findings suggest that a yet to be identified self-antigen may initiate and mediate chronic, (NK)T cell-dependent intestinal inflammation. Moreover, these studies revealed that exposure to the intestinal microbiota during early but not adult life leads to a persistent decrease in intestinal iNKT cell numbers and protection from (NK)T cell-mediated intestinal inflammation during adult life (Olszak et al., 2012). The microbiota therefore plays critical roles in the modulation of (NK)T cells at mucosal surfaces, a phenomenon similarly observed in the lung (Olszak et al., 2012). Future studies will be required to delineate the cellular origin and the nature of presumed self-antigens, which initiate (NK)T cell-dependent inflammation at mucosal surfaces. Similarly, antigen-dependent and antigen-independent environmental modulators of postnatal, mucosal (NK)T cell development await further characterization.
(NK)T cells in the regulation of lipid and carbohydrate metabolism
(NK)T cells are affected by dysregulation of lipid metabolism and, conversely, also actively control lipid and carbohydrate metabolism, thus reflecting a bidirectional process of regulation. Moreover, (NK)T cells modulate disorders at the interface of metabolism and inflammation such as obesity, metabolic syndrome, and non-alcoholic fatty liver disease (NAFLD). Thus, obesity is associated with reduced numbers of iNKT cells in peripheral blood, liver, and adipose tissue in humans and mice, whereas weight loss leads to increasing numbers of peripheral iNKT cells (Guebre-Xabier et al., 2000; Lynch et al., 2009, 2012; Kotas et al., 2011; Ji et al., 2012). Moreover, NAFLD is associated with reduced CD1d expression by hepatocytes and impaired hepatocyte-dependent iNKT cell activation (Yang et al., 2007).
In accordance with bidirectional regulation of (NK)T cells and lipid metabolism, CD1d and Jα18 deficiency in mice, independent of low- or high-fat diet, is associated with adipocyte hypertrophy, increased leptin as well as decreased adiponectin levels, and insulin resistance (Lynch et al., 2012; Schipper et al., 2012). On a high fat diet, CD1d- and Jα18-deficient mice also exhibit increased weight gain and hepatic steatosis (Lynch et al., 2012). In further support of a mechanistic link between iNKT cells and the regulation of glucose and lipid metabolism, transfer of iNKT cells or α-GalCer treatment was associated with reduced adipocyte size, decreased leptin and increased adiponectin levels, increased M2 macrophage polarization, anti-inflammatory IL-10 and IL-4 signaling, and improved insulin sensitivity (Ji et al., 2012; Lynch et al., 2012). Importantly, both hepatocytes and adipocytes express CD1d and can activate iNKT cells in a CD1d-restricted manner (Yang et al., 2007; Schipper et al., 2012). This raises the possibility that obesity-induced alterations in the adipocyte and hepatocyte CD1d self-lipid repertoire may regulate metabolism and obesity-associated inflammation via modulation of iNKT cell activation. Although differences in the extent and outcome of iNKT cell-mediated metabolic modulation were observed between studies (Mantell et al., 2011; Wu et al., 2012), these discrepancies may relate to differences in the intestinal microbiota, which is modulated by host metabolism and regulates iNKT cell function (Olszak et al., 2012; Tremaroli and Backhed, 2012; Wingender et al., 2012).
Together, these studies demonstrate a tight connection between metabolism and immunity and reveal a central role of lipid-reactive T cells in the bidirectional regulation of this process.
Murine studies of group 1 CD1-restricted T cells in immunity
Although a large number of studies in rodent models characterized the role of CD1d-restricted (NK)T cells in vivo, similar studies of CD1a-, CD1b-, and CD1c-restricted T cells were prevented by the lack of group 1 CD1 expression in mice. Our understanding of the function of group 1 CD1-restricted T cells is therefore largely based on the analysis of human T cell clones. These studies revealed the recognition of various mycobacterial lipid antigens by group 1 CD1-restricted T cells, suggesting a potential role of these cells in immunity to mycobacteria and particularly to M. tuberculosis (Moody et al., 2005; Cohen et al., 2009).
The recent development of mice with transgenic expression of human group 1 CD1 provided a crucial step toward the characterization of CD1a-, CD1b-, and CD1c-restricted T cells in vivo (Felio et al., 2009). Initial studies in these mice demonstrated that group 1 CD1-restricted T cells both phenotypically and functionally resemble conventional T cells rather than iNKT cells and exhibit delayed immune responses (Felio et al., 2009). In accordance with previous ex vivo and in vitro studies, which suggested a central role of CD1a, CD1b, and CD1c in the recognition of M. tuberculosis, infection of transgenic mice induced group 1 CD1-restricted T cell responses directed against M. tuberculosis with T cell-dependent recognition of previously characterized M. tuberculosis-derived lipids (Felio et al., 2009). Interestingly, preliminary studies revealed that clearance of M. tuberculosis did not differ between mice with and without group 1 CD1 expression. Further studies are therefore required to delineate the contribution of group 1 CD1 to the immune response against M. tuberculosis and other microorganisms and to define potential roles of CD1a-, CD1b-, and CD1c-restricted T cells in the modulation of autoimmunity, cancer immunosurveillance, and the regulation of metabolic function.
The biological role of lipid-reactive T cells in human immunity
Murine studies revealed a critical role of CD1d-restricted (NK)T cells in antimicrobial immunity and demonstrated susceptibility of CD1d-deficient mice to a vast number of microorganisms (Tupin et al., 2007; Brennan et al., 2013). However, similar observations of broad immunodeficiency were made in mice deficient in TLRs, TLR adaptors, and NK cells, whereas patients carrying mutations in these pathways exhibited few, selected immune defects (Orange, 2002; Zhang et al., 2007; von Bernuth et al., 2008). These studies therefore suggested that the immune response in inbred mice may not fully recapitulate human immunity – a finding recently confirmed in a systematic approach (Casanova and Abel, 2007; Seok et al., 2013).
To define the role of CD1 in human immunity, patients with the rare Mendelian disorder abetalipoproteinemia (ABL) were studied. ABL is characterized by mutations in MTP, an ER-resident lipid transfer protein involved in the generation of apolipoprotein B (ApoB)-containing lipoprotein particles (Wetterau et al., 1992; Berriot-Varoqueaux et al., 2000; Abumrad and Davidson, 2012). Due to deficiency in MTP, ABL patients exhibit defects in the secretion of chylomicrons and very low-density lipoprotein, which is associated with hypolipidemia and severe neurological defects (Berriot-Varoqueaux et al., 2000; Abumrad and Davidson, 2012). In addition to its role in lipoprotein metabolism, MTP can directly transfer phospholipids onto CD1d and is critical for the function of group 1 and group 2 CD1 (Brozovic et al., 2004; Dougan et al., 2005, 2007; Kaser et al., 2008). Accordingly, these studies revealed broad defects in group 1 and group 2 CD1 function in ABL (Zeissig et al., 2010). Group 1 CD1, particularly CD1a and CD1c, underwent proteasomal degradation in patients with ABL, possibly as a consequence of impaired ligand-dependent CD1 stabilization, a fate similarly observed for apolipoprotein B (ApoB) (Berriot-Varoqueaux et al., 2000; Zeissig et al., 2010). CD1d escaped proteasomal degradation and showed normal cell surface expression but was unable to load and present exogenous lipid antigens and did not elicit (NK)T cell autoreactivity. Consequently, iNKT cells were either not detectable or showed severe phenotypic alterations in ABL. In contrast to CD1, MHC class I- and class II-restricted antigen presentation and conventional T- and B-cell function were unaltered. ABL is thus characterized by selective and severe defects in CD1 and provides a unique opportunity to study the biological relevance of CD1-restricted (NK)T cells.
Although fewer than 100 patients with ABL have been clinically characterized, several cases of heart failure due to lesions resembling chronic interstitial myocarditis, fatal pulmonary infections, and rare malignancies have been described in ABL (Berriot-Varoqueaux et al., 2000). Given the central role of (NK)T cells in immunity to pulmonary and cardiotropic microbial pathogens as well as cancer immunity in mice, these findings support the concept of immune-related defects due to deficiency in MTP and CD1 (Zeissig et al., 2010; Zeissig and Blumberg, 2012). However, the spectrum of immune defects in ABL is remarkably restricted, which contrasts with observations in CD1d-deficient mice. Although such conclusions are limited to frequently encountered microorganisms given the low incidence of ABL, they reflect findings made for human and murine TLR and NK cell deficiency as outlined above (Orange, 2002; Zhang et al., 2007; von Bernuth et al., 2008). Together, these observations suggest that CD1-restricted (NK)T cells have critical and indispensable contributions to human and murine immunity but that mechanisms that compensate for the loss of functional CD1 prevent broad immunodeficiency, at least in humans.
Concluding remarks
Significant insight into the role of lipid-reactive T cells has been gained over the past 25 years. As such, antigen-dependent and antigen-independent pathways to the activation of lipid-reactive T cells were described, a large number of self and microbial lipid antigens were identified, and the roles of lipid-reactive T cells in immunity were characterized. Despite these significant advances, however, further studies are required to characterize the dynamics of the CD1 lipidome in health and disease, to gain further insight into the physiological role of group 1 CD1-restricted T cells in vivo, to delineate the functions of invariant and non-invariant NKT cells, and to define the mechanisms of (NK)T cell activation in autoimmune diseases and cancer and their modulation by environmental influences such as the microbiota.
About the authors

C. Marie Dowds studied Biosciences at the University of Kaiserslautern, Germany. After completion of her master’s degree in the group of Dr. Stefan Kins, she joined the laboratory of Dr. Zeissig at the Department of Internal Medicine, Kiel University (Kiel, Germany) as PhD student. In her PhD studies, Dowds investigates the role of NKT cells and lipid antigens in intestinal inflammation.
Sabin-Christin Kornell studied Biology at the Christian-Albrechts-University of Kiel and received her master’s degree in cell biology and physiology in 2013. In May 2013, Kornell started her PhD studies in Dr. Zeissig’s group at the Department of Internal Medicine, Kiel University, Germany (Kiel, Germany).
Richard S. Blumberg, MD, is Professor of Medicine at Harvard Medical School, Chief of the Division of Gastroenterology, Hepatology and Endoscopy at Brigham and Women’s Hospital, co-Director of the Biomedical Research Institute at Brigham and Women’s Hospital and co-Director of the Harvard Digestive Diseases Center. He received his medical degree from Jefferson Medical College (Philadelphia, PA), internship and residency at The New York Hospital, Cornell Medical Center followed by fellowship training in infectious diseases at Massachusetts General Hospital, gastroenterology at Brigham and Women’s Hospital and molecular immunology at Dana Farber Cancer Institute, all in Boston. His work focuses on the mechanisms that control homeostasis and inflammation at mucosal surfaces.
Sebastian Zeissig, MD, received his medical degree from the Charité Berlin, Germany. After a postdoctoral fellowship at the same institution, Dr. Zeissig joined the laboratory of Richard S. Blumberg as postdoctoral fellow to investigate the role of CD1d-restricted NKT cells in infectious hepatitis and immunodeficiency. Since 2010, Dr. Zeissig is leading the Laboratory of Mucosal Immunology at the Department of Internal Medicine, Kiel University (Kiel, Germany), where he continues to investigate the role of lipid antigens in intestinal and hepatic immunity.
We apologize that due to constraints in the length of this manuscript and the number of citations, we had to focus on selected aspects of CD1 biology and could only highlight few, selected studies. Experimental studies by the authors were supported by the Deutsche Forschungsgemeinschaft (DFG) (ZE 814/1-1, 2-1, 4-1; DFG Cluster ‘Inflammation at Interfaces’) and the European Commission (Marie-Curie International Reintegration Grant, FP7) (S.Z.); NIH grants DK044319, DK051362, DK053056, and DK088199; the Harvard Digestive Diseases Center (HDDC) (DK0034854) (R.S.B.).
References
Abumrad, N.A. and Davidson, N.O. (2012). Role of the gut in lipid homeostasis. Physiol. Rev. 92, 1061–1085.10.1152/physrev.00019.2011Suche in Google Scholar PubMed PubMed Central
Amprey, J.L., Im, J.S., Turco, S.J., Murray, H.W., Illarionov, P.A., Besra, G.S., Porcelli, S.A., and Spath, G.F. (2004). A subset of liver NK T cells is activated during Leishmania donovani infection by CD1d-bound lipophosphoglycan. J. Exp. Med. 200, 895–904.10.1084/jem.20040704Suche in Google Scholar PubMed PubMed Central
Anson, M., Crain-Denoyelle, A.M., Baud, V., Chereau, F., Gougelet, A., Terris, B., Yamagoe, S., Colnot, S., Viguier, M., Perret, C., et al. (2012). Oncogenic β-catenin triggers an inflammatory response that determines the aggressiveness of hepatocellular carcinoma in mice. J. Clin. Invest. 122, 586–599.10.1172/JCI43937Suche in Google Scholar PubMed PubMed Central
Arrenberg, P., Halder, R., Dai, Y., Maricic, I., and Kumar, V. (2010). Oligoclonality and innate-like features in the TCR repertoire of type II NKT cells reactive to a β-linked self-glycolipid. Proc. Natl. Acad. Sci. USA 107, 10984–10989.10.1073/pnas.1000576107Suche in Google Scholar PubMed PubMed Central
Balk, S.P., Ebert, E.C., Blumenthal, R.L., McDermott, F.V., Wucherpfennig, K.W., Landau, S.B., and Blumberg, R.S. (1991). Oligoclonal expansion and CD1 recognition by human intestinal intraepithelial lymphocytes. Science 253, 1411–1415.10.1126/science.1716785Suche in Google Scholar PubMed
Balk, S.P., Burke, S., Polischuk, J.E., Frantz, M.E., Yang, L., Porcelli, S., Colgan, S.P., and Blumberg, R.S. (1994). Beta 2-microglobulin-independent MHC class Ib molecule expressed by human intestinal epithelium. Science 265, 259–262.10.1126/science.7517575Suche in Google Scholar PubMed
Beckman, E.M., Porcelli, S.A., Morita, C.T., Behar, S.M., Furlong, S.T., and Brenner, M.B. (1994). Recognition of a lipid antigen by CD1-restricted αβ+ T cells. Nature 372, 691–694.10.1038/372691a0Suche in Google Scholar PubMed
Bedel, R., Matsuda, J.L., Brigl, M., White, J., Kappler, J., Marrack, P., and Gapin, L. (2012). Lower TCR repertoire diversity in Traj18-deficient mice. Nat. Immunol. 13, 705–706.10.1038/ni.2347Suche in Google Scholar PubMed PubMed Central
Bendelac, A., Savage, P.B., and Teyton, L. (2007). The biology of NKT cells. Annu. Rev. Immunol. 25, 297–336.10.1146/annurev.immunol.25.022106.141711Suche in Google Scholar PubMed
Benlagha, K., Wei, D.G., Veiga, J., Teyton, L., and Bendelac, A. (2005). Characterization of the early stages of thymic NKT cell development. J. Exp. Med. 202, 485–942.10.1084/jem.20050456Suche in Google Scholar PubMed PubMed Central
Berntman, E., Rolf, J., Johansson, C., Anderson, P., and Cardell, S.L. (2005). The role of CD1d-restricted NK T lymphocytes in the immune response to oral infection with Salmonella typhimurium. Eur. J. Immunol. 35, 2100–2109.10.1002/eji.200425846Suche in Google Scholar PubMed
Berriot-Varoqueaux, N., Aggerbeck, L.P., Samson-Bouma, M., and Wetterau, J.R. (2000). The role of the microsomal triglygeride transfer protein in abetalipoproteinemia. Annu. Rev. Nutr. 20, 663–697.10.1146/annurev.nutr.20.1.663Suche in Google Scholar PubMed
Berzofsky, J.A. and Terabe, M. (2009). The contrasting roles of NKT cells in tumor immunity. Curr. Mol. Med. 9, 667–672.10.2174/156652409788970706Suche in Google Scholar PubMed PubMed Central
Blumberg, R.S., Terhorst, C., Bleicher, P., McDermott, F.V., Allan, C.H., Landau, S.B., Trier, J.S., and Balk, S.P. (1991). Expression of a nonpolymorphic MHC class I-like molecule, CD1D, by human intestinal epithelial cells. J. Immunol. 147, 2518–2524.10.4049/jimmunol.147.8.2518Suche in Google Scholar
Brennan, P.J., Tatituri, R.V., Brigl, M., Kim, E.Y., Tuli, A., Sanderson, J.P., Gadola, S.D., Hsu, F.F., Besra, G.S., and Brenner, M.B. (2011). Invariant natural killer T cells recognize lipid self antigen induced by microbial danger signals. Nat. Immunol. 12, 1202–1211.10.1038/ni.2143Suche in Google Scholar PubMed PubMed Central
Brennan, P.J., Brigl, M., and Brenner, M.B. (2013). Invariant natural killer T cells: an innate activation scheme linked to diverse effector functions. Nat. Rev. Immunol. 13, 101–117.10.1038/nri3369Suche in Google Scholar PubMed
Bricard, G., Cesson, V., Devevre, E., Bouzourene, H., Barbey, C., Rufer, N., Im, J.S., Alves, P.M., Martinet, O., Halkic, N., et al. (2009). Enrichment of human CD4+ Vα24/Vβ11 invariant NKT cells in intrahepatic malignant tumors. J. Immunol. 182, 5140–5151.10.4049/jimmunol.0711086Suche in Google Scholar PubMed
Brigl, M., Bry, L., Kent, S.C., Gumperz, J.E., and Brenner, M.B. (2003). Mechanism of CD1d-restricted natural killer T cell activation during microbial infection. Nat. Immunol. 4, 1230–1237.10.1038/ni1002Suche in Google Scholar PubMed
Brigl, M., Tatituri, R.V., Watts, G.F., Bhowruth, V., Leadbetter, E.A., Barton, N., Cohen, N.R., Hsu, F.F., Besra, G.S., and Brenner, M.B. (2011). Innate and cytokine-driven signals, rather than microbial antigens, dominate in natural killer T cell activation during microbial infection. J. Exp. Med. 208, 1163–1177.10.1084/jem.20102555Suche in Google Scholar PubMed PubMed Central
Briken, V., Jackman, R.M., Watts, G.F., Rogers, R.A., and Porcelli, S.A. (2000). Human CD1b and CD1c isoforms survey different intracellular compartments for the presentation of microbial lipid antigens. J. Exp. Med. 192, 281–288.10.1084/jem.192.2.281Suche in Google Scholar PubMed PubMed Central
Brozovic, S., Nagaishi, T., Yoshida, M., Betz, S., Salas, A., Chen, D., Kaser, A., Glickman, J., Kuo, T., Little, A., et al. (2004). CD1d function is regulated by microsomal triglyceride transfer protein. Nat. Med. 10, 535–539.10.1038/nm1043Suche in Google Scholar PubMed
Cala-De Paepe, D., Layre, E., Giacometti, G., Garcia-Alles, L.F., Mori, L., Hanau, D., de Libero, G., de la Salle, H., Puzo, G., and Gilleron, M. (2012). Deciphering the role of CD1e protein in mycobacterial phosphatidyl-myo-inositol mannosides (PIM) processing for presentation by CD1b to T lymphocytes. J. Biol. Chem. 287, 31494–31502.10.1074/jbc.M112.386300Suche in Google Scholar
Camelo, A., Barlow, J.L., Drynan, L.F., Neill, D.R., Ballantyne, S.J., Wong, S.H., Pannell, R., Gao, W., Wrigley, K., Sprenkle, J., et al. (2012). Blocking IL-25 signalling protects against gut inflammation in a type-2 model of colitis by suppressing nuocyte and NKT derived IL-13. J. Gastroenterol. 47, 1198–1211.10.1007/s00535-012-0591-2Suche in Google Scholar
Cardell, S., Tangri, S., Chan, S., Kronenberg, M., Benoist, C., and Mathis, D. (1995). CD1-restricted CD4+ T cells in major histocompatibility complex class II-deficient mice. J. Exp. Med. 182, 993–1004.10.1084/jem.182.4.993Suche in Google Scholar
Casanova, J.L. and Abel, L. (2007). Primary immunodeficiencies: a field in its infancy. Science 317, 617–619.10.1126/science.1142963Suche in Google Scholar
Chang, Y.J., Kim, H.Y., Albacker, L.A., Lee, H.H., Baumgarth, N., Akira, S., Savage, P.B., Endo, S., Yamamura, T., Maaskant, J., et al. (2011). Influenza infection in suckling mice expands an NKT cell subset that protects against airway hyperreactivity. J. Clin. Invest. 121, 57–69.10.1172/JCI44845Suche in Google Scholar
Chen, N., McCarthy, C., Drakesmith, H., Li, D., Cerundolo, V., McMichael, A.J., Screaton, G.R., and Xu, X.N. (2006). HIV-1 down-regulates the expression of CD1d via Nef. Eur. J. Immunol. 36, 278–286.10.1002/eji.200535487Suche in Google Scholar
Chen, Y.H., Chiu, N.M., Mandal, M., Wang, N., and Wang, C.R. (1997). Impaired NK1+ T cell development and early IL-4 production in CD1-deficient mice. Immunity 6, 459–467.10.1016/S1074-7613(00)80289-7Suche in Google Scholar
Chiu, Y.H., Jayawardena, J., Weiss, A., Lee, D., Park, S.H., Dautry-Varsat, A., and Bendelac, A. (1999). Distinct subsets of CD1d-restricted T cells recognize self-antigens loaded in different cellular compartments. J. Exp. Med. 189, 103–110.10.1084/jem.189.1.103Suche in Google Scholar PubMed PubMed Central
Chiu, Y.H., Park, S.H., Benlagha, K., Forestier, C., Jayawardena-Wolf, J., Savage, P.B., Teyton, L., and Bendelac, A. (2002). Multiple defects in antigen presentation and T cell development by mice expressing cytoplasmic tail-truncated CD1d. Nat. Immunol. 3, 55–60.10.1038/ni740Suche in Google Scholar PubMed
Cho, S., Knox, K.S., Kohli, L.M., He, J.J., Exley, M.A., Wilson, S.B., and Brutkiewicz, R.R. (2005). Impaired cell surface expression of human CD1d by the formation of an HIV-1 Nef/CD1d complex. Virology 337, 242–252.10.1016/j.virol.2005.04.020Suche in Google Scholar PubMed
Cohen, N.R., Garg, S., and Brenner, M.B. (2009). Antigen presentation by CD1 lipids, T cells, and NKT cells in microbial immunity. Adv. Immunol. 102, 1–94.10.1016/S0065-2776(09)01201-2Suche in Google Scholar
Cohen, N.R., Tatituri, R.V., Rivera, A., Watts, G.F., Kim, E.Y., Chiba, A., Fuchs, B.B., Mylonakis, E., Besra, G.S., Levitz, S.M., et al. (2011). Innate recognition of cell wall β-glucans drives invariant natural killer T cell responses against fungi. Cell Host Microbe 10, 437–450.10.1016/j.chom.2011.09.011Suche in Google Scholar PubMed PubMed Central
Constantinides, M.G., Picard, D., Savage, A.K., and Bendelac, A. (2011). A naive-like population of human CD1d-restricted T cells expressing intermediate levels of promyelocytic leukemia zinc finger. J. Immunol. 187, 309–315.10.4049/jimmunol.1100761Suche in Google Scholar PubMed PubMed Central
Cox, D., Fox, L., Tian, R., Bardet, W., Skaley, M., Mojsilovic, D., Gumperz, J., and Hildebrand, W. (2009). Determination of cellular lipids bound to human CD1d molecules. PLoS One 4, e5325.10.1371/journal.pone.0005325Suche in Google Scholar PubMed PubMed Central
Crowe, N.Y., Smyth, M.J., and Godfrey, D.I. (2002). A critical role for natural killer T cells in immunosurveillance of methylcholanthrene-induced sarcomas. J. Exp. Med. 196, 119–127.10.1084/jem.20020092Suche in Google Scholar PubMed PubMed Central
Crowe, N.Y., Coquet, J.M., Berzins, S.P., Kyparissoudis, K., Keating, R., Pellicci, D.G., Hayakawa, Y., Godfrey, D.I., and Smyth, M.J. (2005). Differential antitumor immunity mediated by NKT cell subsets in vivo. J. Exp. Med. 202, 1279–1288.10.1084/jem.20050953Suche in Google Scholar PubMed PubMed Central
Cui, J., Shin, T., Kawano, T., Sato, H., Kondo, E., Toura, I., Kaneko, Y., Koseki, H., Kanno, M., and Taniguchi, M. (1997). Requirement for Vα14 NKT cells in IL-12-mediated rejection of tumors. Science 278, 1623–1626.10.1126/science.278.5343.1623Suche in Google Scholar PubMed
de Jong, A., Pena-Cruz, V., Cheng, T.Y., Clark, R.A., Van Rhijn, I., and Moody, D.B. (2010). CD1a-autoreactive T cells are a normal component of the human αβ T cell repertoire. Nat. Immunol. 11, 1102–1109.10.1038/ni.1956Suche in Google Scholar PubMed PubMed Central
de la Salle, H., Mariotti, S., Angenieux, C., Gilleron, M., Garcia-Alles, L.F., Malm, D., Berg, T., Paoletti, S., Maitre, B., Mourey, L., et al. (2005). Assistance of microbial glycolipid antigen processing by CD1e. Science 310, 1321–1324.10.1126/science.1115301Suche in Google Scholar PubMed
de Lalla, C., Lepore, M., Piccolo, F.M., Rinaldi, A., Scelfo, A., Garavaglia, C., Mori, L., De Libero, G., Dellabona, P., and Casorati, G. (2011). High-frequency and adaptive-like dynamics of human CD1 self-reactive T cells. Eur. J. Immunol. 41, 602–610.10.1002/eji.201041211Suche in Google Scholar PubMed
De Libero, G., Moran, A.P., Gober, H.J., Rossy, E., Shamshiev, A., Chelnokova, O., Mazorra, Z., Vendetti, S., Sacchi, A., Prendergast, M.M., et al. (2005). Bacterial infections promote T cell recognition of self-glycolipids. Immunity 22, 763–772.10.1016/j.immuni.2005.04.013Suche in Google Scholar
De Libero, G., Collmann, A., and Mori, L. (2009). The cellular and biochemical rules of lipid antigen presentation. Eur. J. Immunol. 39, 2648–2656.10.1002/eji.200939425Suche in Google Scholar
Dhodapkar, M.V., Geller, M.D., Chang, D.H., Shimizu, K., Fujii, S., Dhodapkar, K.M., and Krasovsky, J. (2003). A reversible defect in natural killer T cell function characterizes the progression of premalignant to malignant multiple myeloma. J. Exp. Med. 197, 1667–1676.10.1084/jem.20021650Suche in Google Scholar
Donovan, M.J., Jayakumar, A., and McDowell, M.A. (2007). Inhibition of groups 1 and 2 CD1 molecules on human dendritic cells by Leishmania species. Parasite Immunol. 29, 515–524.10.1111/j.1365-3024.2007.00970.xSuche in Google Scholar
Dougan, S.K., Salas, A., Rava, P., Agyemang, A., Kaser, A., Morrison, J., Khurana, A., Kronenberg, M., Johnson, C., Exley, M., Hussain, M.M., and Blumberg, R.S. (2005). Microsomal triglyceride transfer protein lipidation and control of CD1d on antigen-presenting cells. J. Exp. Med. 202, 529–539.10.1084/jem.20050183Suche in Google Scholar
Dougan, S.K., Rava, P., Hussain, M.M., and Blumberg, R.S. (2007). MTP regulated by an alternate promoter is essential for NKT cell development. J. Exp. Med. 204, 533–545.10.1084/jem.20062006Suche in Google Scholar
Ernst, W.A., Maher, J., Cho, S., Niazi, K.R., Chatterjee, D., Moody, D.B., Besra, G.S., Watanabe, Y., Jensen, P.E., Porcelli, S.A., et al. (1998). Molecular interaction of CD1b with lipoglycan antigens. Immunity 8, 331–340.10.1016/S1074-7613(00)80538-5Suche in Google Scholar
Facciotti, F., Cavallari, M., Angenieux, C., Garcia-Alles, L.F., Signorino-Gelo, F., Angman, L., Gilleron, M., Prandi, J., Puzo, G., Panza, L., et al. (2011). Fine tuning by human CD1e of lipid-specific immune responses. Proc. Natl. Acad. Sci. USA 108, 14228–14233.10.1073/pnas.1108809108Suche in Google Scholar PubMed PubMed Central
Facciotti, F., Ramanjaneyulu, G.S., Lepore, M., Sansano, S., Cavallari, M., Kistowska, M., Forss-Petter, S., Ni, G., Colone, A., Singhal, A., et al. (2012). Peroxisome-derived lipids are self antigens that stimulate invariant natural killer T cells in the thymus. Nat. Immunol. 13, 474–480.10.1038/ni.2245Suche in Google Scholar PubMed
Felio, K., Nguyen, H., Dascher, C.C., Choi, H.J., Li, S., Zimmer, M.I., Colmone, A., Moody, D.B., Brenner, M.B., and Wang, C.R. (2009). CD1-restricted adaptive immune responses to Mycobacteria in human group 1 CD1 transgenic mice. J. Exp. Med. 206, 2497–2509.10.1084/jem.20090898Suche in Google Scholar PubMed PubMed Central
Fischer, K., Scotet, E., Niemeyer, M., Koebernick, H., Zerrahn, J., Maillet, S., Hurwitz, R., Kursar, M., Bonneville, M., Kaufmann, S.H., et al. (2004). Mycobacterial phosphatidylinositol mannoside is a natural antigen for CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA 101, 10685–10690.10.1073/pnas.0403787101Suche in Google Scholar PubMed PubMed Central
Florence, W.C., Xia, C., Gordy, L.E., Chen, W., Zhang, Y., Scott-Browne, J., Kinjo, Y., Yu, K.O., Keshipeddy, S., Pellicci, D.G., et al. (2009). Adaptability of the semi-invariant natural killer T-cell receptor towards structurally diverse CD1d-restricted ligands. EMBO J. 28, 3781.10.1038/emboj.2009.348Suche in Google Scholar PubMed PubMed Central
Fox, L.M., Cox, D.G., Lockridge, J.L., Wang, X., Chen, X., Scharf, L., Trott, D.L., Ndonye, R.M., Veerapen, N., Besra, G.S., et al. (2009). Recognition of lyso-phospholipids by human natural killer T lymphocytes. PLoS Biol. 7, e1000228.10.1371/journal.pbio.1000228Suche in Google Scholar PubMed PubMed Central
Fuchs, B., Muller, K., Paasch, U., and Schiller, J. (2012). Lysophospholipids: potential markers of diseases and infertility? Mini Rev. Med. Chem. 12, 74–86.Suche in Google Scholar
Fujii, S., Shimizu, K., Smith, C., Bonifaz, L., and Steinman, R.M. (2003). Activation of natural killer T cells by alpha-galactosylceramide rapidly induces the full maturation of dendritic cells in vivo and thereby acts as an adjuvant for combined CD4 and CD8 T cell immunity to a coadministered protein. J. Exp. Med. 198, 267–279.10.1084/jem.20030324Suche in Google Scholar PubMed PubMed Central
Fuss, I.J., Heller, F., Boirivant, M., Leon, F., Yoshida, M., Fichtner-Feigl, S., Yang, Z., Exley, M., Kitani, A., Blumberg, R.S., et al. (2004). Nonclassical CD1d-restricted NK T cells that produce IL-13 characterize an atypical Th2 response in ulcerative colitis. J. Clin. Invest. 113, 1490–1497.10.1172/JCI19836Suche in Google Scholar PubMed PubMed Central
Gadola, S.D., Silk, J.D., Jeans, A., Illarionov, P.A., Salio, M., Besra, G.S., Dwek, R., Butters, T.D., Platt, F.M., and Cerundolo, V. (2006). Impaired selection of invariant natural killer T cells in diverse mouse models of glycosphingolipid lysosomal storage diseases. J. Exp. Med. 203, 2293–2303.10.1084/jem.20060921Suche in Google Scholar PubMed PubMed Central
Gapin, L., Matsuda, J.L., Surh, C.D., and Kronenberg, M. (2001). NKT cells derive from double-positive thymocytes that are positively selected by CD1d. Nat. Immunol. 2, 971–978.10.1038/ni710Suche in Google Scholar PubMed
Garcia-Alles, L.F., Versluis, K., Maveyraud, L., Vallina, A.T., Sansano, S., Bello, N.F., Gober, H.J., Guillet, V., de la Salle, H., Puzo, G., et al. (2006). Endogenous phosphatidylcholine and a long spacer ligand stabilize the lipid-binding groove of CD1b. EMBO J. 25, 3684–3692.10.1038/sj.emboj.7601244Suche in Google Scholar PubMed PubMed Central
Garcia-Alles, L.F., Giacometti, G., Versluis, C., Maveyraud, L., de Paepe, D., Guiard, J., Tranier, S., Gilleron, M., Prandi, J., Hanau, D., et al. (2011). Crystal structure of human CD1e reveals a groove suited for lipid-exchange processes. Proc. Natl. Acad. Sci. USA 108, 13230–13235.10.1073/pnas.1105627108Suche in Google Scholar PubMed PubMed Central
Garzon, D., Anselmi, C., Bond, P.J., and Faraldo-Gomez, J.D. (2013). Dynamics of the antigen-binding grooves in CD1 proteins: reversible hydrophobic collapse in the lipid-free state. J. Biol. Chem. 288, 19528–19536.10.1074/jbc.M113.470179Suche in Google Scholar PubMed PubMed Central
Geissmann, F., Cameron, T.O., Sidobre, S., Manlongat, N., Kronenberg, M., Briskin, M.J., Dustin, M.L., and Littman, D.R. (2005). Intravascular immune surveillance by CXCR6+ NKT cells patrolling liver sinusoids. PLoS Biol. 3, e113.10.1371/journal.pbio.0030113Suche in Google Scholar PubMed PubMed Central
Giaccone, G., Punt, C.J., Ando, Y., Ruijter, R., Nishi, N., Peters, M., von Blomberg, B.M., Scheper, R.J., van der Vliet, H.J., van den Eertwegh, A.J., et al. (2002). A phase I study of the natural killer T-cell ligand α-galactosylceramide (KRN7000) in patients with solid tumors. Clin. Cancer Res. 8, 3702–3709.Suche in Google Scholar
Girardi, E. and Zajonc, D.M. (2012). Molecular basis of lipid antigen presentation by CD1d and recognition by natural killer T cells. Immunol. Rev. 250, 167–179.10.1111/j.1600-065X.2012.01166.xSuche in Google Scholar PubMed PubMed Central
Girardi, E., Maricic, I., Wang, J., Mac, T.T., Iyer, P., Kumar, V., and Zajonc, D.M. (2012). Type II natural killer T cells use features of both innate-like and conventional T cells to recognize sulfatide self antigens. Nat. Immunol. 13, 851–856.10.1038/ni.2371Suche in Google Scholar PubMed PubMed Central
Gogolak, P., Rethi, B., Szatmari, I., Lanyi, A., Dezso, B., Nagy, L., and Rajnavolgyi, E. (2007). Differentiation of CD1a- and CD1a+ monocyte-derived dendritic cells is biased by lipid environment and PPARγ. Blood 109, 643–652.10.1182/blood-2006-04-016840Suche in Google Scholar PubMed
Grant, E.P., Degano, M., Rosat, J.P., Stenger, S., Modlin, R.L., Wilson, I.A., Porcelli, S.A., and Brenner, M.B. (1999). Molecular recognition of lipid antigens by T cell receptors. J. Exp. Med. 189, 195–205.10.1084/jem.189.1.195Suche in Google Scholar PubMed PubMed Central
Guebre-Xabier, M., Yang, S., Lin, H.Z., Schwenk, R., Krzych, U., and Diehl, A.M. (2000). Altered hepatic lymphocyte subpopulations in obesity-related murine fatty livers: potential mechanism for sensitization to liver damage. Hepatology 31, 633–640.10.1002/hep.510310313Suche in Google Scholar PubMed
Hage, C.A., Kohli, L.L., Cho, S., Brutkiewicz, R.R., Twigg, H.L., 3rd, and Knox, K.S. (2005). Human immunodeficiency virus gp120 downregulates CD1d cell surface expression. Immunol. Lett. 98, 131–135.10.1016/j.imlet.2004.10.025Suche in Google Scholar PubMed
Haig, N.A., Guan, Z., Li, D., McMichael, A., Raetz, C.R., and Xu, X.N. (2011). Identification of self-lipids presented by CD1c and CD1d proteins. J. Biol. Chem. 286, 37692–37701.10.1074/jbc.M111.267948Suche in Google Scholar PubMed PubMed Central
Halder, R.C., Aguilera, C., Maricic, I., and Kumar, V. (2007). Type II NKT cell-mediated anergy induction in type I NKT cells prevents inflammatory liver disease. J. Clin. Invest. 117, 2302–2312.10.1172/JCI31602Suche in Google Scholar PubMed PubMed Central
Hegde, S., Chen, X., Keaton, J.M., Reddington, F., Besra, G.S., and Gumperz, J.E. (2007). NKT cells direct monocytes into a DC differentiation pathway. J. Leukoc. Biol. 81, 1224–1235.10.1189/jlb.1206718Suche in Google Scholar PubMed
Heller, F., Fuss, I.J., Nieuwenhuis, E.E., Blumberg, R.S., and Strober, W. (2002). Oxazolone colitis, a Th2 colitis model resembling ulcerative colitis, is mediated by IL-13-producing NK-T cells. Immunity 17, 629–638.10.1016/S1074-7613(02)00453-3Suche in Google Scholar
Heller, F., Florian, P., Bojarski, C., Richter, J., Christ, M., Hillenbrand, B., Mankertz, J., Gitter, A.H., Burgel, N., Fromm, M., et al. (2005). Interleukin-13 is the key effector Th2 cytokine in ulcerative colitis that affects epithelial tight junctions, apoptosis, and cell restitution. Gastroenterology 129, 550–564.10.1016/j.gastro.2005.05.002Suche in Google Scholar
Hermans, I.F., Silk, J.D., Gileadi, U., Salio, M., Mathew, B., Ritter, G., Schmidt, R., Harris, A.L., Old, L., and Cerundolo, V. (2003). NKT cells enhance CD4+ and CD8+ T cell responses to soluble antigen in vivo through direct interaction with dendritic cells. J. Immunol. 171, 5140–5147.10.4049/jimmunol.171.10.5140Suche in Google Scholar
Hermans, I.F., Silk, J.D., Gileadi, U., Masri, S.H., Shepherd, D., Farrand, K.J., Salio, M., and Cerundolo, V. (2007). Dendritic cell function can be modulated through cooperative actions of TLR ligands and invariant NKT cells. J. Immunol. 178, 2721–2729.10.4049/jimmunol.178.5.2721Suche in Google Scholar
Honey, K., Benlagha, K., Beers, C., Forbush, K., Teyton, L., Kleijmeer, M.J., Rudensky, A.Y., and Bendelac, A. (2002). Thymocyte expression of cathepsin L is essential for NKT cell development. Nat. Immunol. 3, 1069–1074.10.1038/ni844Suche in Google Scholar
Huang, S., Cheng, T.Y., Young, D.C., Layre, E., Madigan, C.A., Shires, J., Cerundolo, V., Altman, J.D., and Moody, D.B. (2011). Discovery of deoxyceramides and diacylglycerols as CD1b scaffold lipids among diverse groove-blocking lipids of the human CD1 system. Proc. Natl. Acad. Sci. USA 108, 19335–19340.10.1073/pnas.1112969108Suche in Google Scholar
Ishikawa, A., Motohashi, S., Ishikawa, E., Fuchida, H., Higashino, K., Otsuji, M., Iizasa, T., Nakayama, T., Taniguchi, M., and Fujisawa, T. (2005). A phase I study of alpha-galactosylceramide (KRN7000)-pulsed dendritic cells in patients with advanced and recurrent non-small cell lung cancer. Clin. Cancer Res. 11, 1910–1917.10.1158/1078-0432.CCR-04-1453Suche in Google Scholar
Izhak, L., Ambrosino, E., Kato, S., Parish, S.T., O’Konek, J.J., Weber, H., Xia, Z., Venzon, D., Berzofsky, J.A., and Terabe, M. (2013). Delicate balance among three types of T cells in concurrent regulation of tumor immunity. Cancer Res. 73, 1514–1523.10.1158/0008-5472.CAN-12-2567Suche in Google Scholar
Jahng, A., Maricic, I., Aguilera, C., Cardell, S., Halder, R.C., and Kumar, V. (2004). Prevention of autoimmunity by targeting a distinct, noninvariant CD1d-reactive T cell population reactive to sulfatide. J. Exp. Med. 199, 947–957.10.1084/jem.20031389Suche in Google Scholar
Jayawardena-Wolf, J., Benlagha, K., Chiu, Y.H., Mehr, R., and Bendelac, A. (2001). CD1d endosomal trafficking is independently regulated by an intrinsic CD1d-encoded tyrosine motif and by the invariant chain. Immunity 15, 897–908.10.1016/S1074-7613(01)00240-0Suche in Google Scholar
Ji, Y., Sun, S., Xu, A., Bhargava, P., Yang, L., Lam, K.S., Gao, B., Lee, C.H., Kersten, S., and Qi, L. (2012). Activation of natural killer T cells promotes M2 Macrophage polarization in adipose tissue and improves systemic glucose tolerance via interleukin-4 (IL-4)/STAT6 protein signaling axis in obesity. J. Biol. Chem. 287, 13561–13571.10.1074/jbc.M112.350066Suche in Google Scholar
Jostins, L., Ripke, S., Weersma, R.K., Duerr, R.H., McGovern, D.P., Hui, K.Y., Lee, J.C., Schumm, L.P., Sharma, Y., Anderson, C.A., et al. (2012). Host-microbe interactions have shaped the genetic architecture of inflammatory bowel disease. Nature 491, 119–124.10.1038/nature11582Suche in Google Scholar
Kang, S.J. and Cresswell, P. (2002). Calnexin, calreticulin, and ERp57 cooperate in disulfide bond formation in human CD1d heavy chain. J. Biol. Chem. 277, 44838–44844.10.1074/jbc.M207831200Suche in Google Scholar
Kang, S.J. and Cresswell, P. (2004). Saposins facilitate CD1d-restricted presentation of an exogenous lipid antigen to T cells. Nat. Immunol. 5, 175–181.10.1038/ni1034Suche in Google Scholar
Karner, M., Kocjan, A., Stein, J., von Boyen, G., Uebel, P., Schmidt, C., Kupcinskas, L., Dina, I., Hanemann, A., Zuelch, F., et al. (2012). Modified release phosphatidylcholine LT-02 in active ulcerative colitis – a randomized, placebo-controlled multicentre study. Gastroenterology 142, S–211.10.1016/S0016-5085(12)60790-2Suche in Google Scholar
Kaser, A., Hava, D.L., Dougan, S.K., Chen, Z., Zeissig, S., Brenner, M.B., and Blumberg, R.S. (2008). Microsomal triglyceride transfer protein regulates endogenous and exogenous antigen presentation by group 1 CD1 molecules. Eur. J. Immunol. 38, 2351–2359.10.1002/eji.200738102Suche in Google Scholar PubMed PubMed Central
Kasmar, A., Van Rhijn, I., and Moody, D.B. (2009). The evolved functions of CD1 during infection. Curr. Opin Immunol. 21, 397–403.10.1016/j.coi.2009.05.022Suche in Google Scholar PubMed PubMed Central
Kasmar, A.G., van Rhijn, I., Cheng, T.Y., Turner, M., Seshadri, C., Schiefner, A., Kalathur, R.C., Annand, J.W., de Jong, A., Shires, J., et al. (2011). CD1b tetramers bind αβ T cell receptors to identify a mycobacterial glycolipid-reactive T cell repertoire in humans. J. Exp. Med. 208, 1741–1747.10.1084/jem.20110665Suche in Google Scholar PubMed PubMed Central
Kawano, T., Cui, J., Koezuka, Y., Toura, I., Kaneko, Y., Motoki, K., Ueno, H., Nakagawa, R., Sato, H., Kondo, E., et al. (1997). CD1d-restricted and TCR-mediated activation of vα14 NKT cells by glycosylceramides. Science 278, 1626–1629.10.1126/science.278.5343.1626Suche in Google Scholar PubMed
Kawana, K., Quayle, A.J., Ficarra, M., Ibana, J.A., Shen, L., Kawana, Y., Yang, H., Marrero, L., Yavagal, S., Greene, S.J., et al. (2007). CD1d degradation in Chlamydia trachomatis-infected epithelial cells is the result of both cellular and chlamydial proteasomal activity. J. Biol. Chem. 282, 7368–7375.10.1074/jbc.M610754200Suche in Google Scholar PubMed
Kinjo, Y. and Kronenberg, M. (2009). Detection of microbes by natural killer T cells. Adv. Exp. Med. Biol. 633, 17–26.10.1007/978-0-387-79311-5_3Suche in Google Scholar PubMed
Kinjo, Y., Wu, D., Kim, G., Xing, G.W., Poles, M.A., Ho, D.D., Tsuji, M., Kawahara, K., Wong, C.H., and Kronenberg, M. (2005). Recognition of bacterial glycosphingolipids by natural killer T cells. Nature 434, 520–525.10.1038/nature03407Suche in Google Scholar PubMed
Kinjo, Y., Tupin, E., Wu, D., Fujio, M., Garcia-Navarro, R., Benhnia, M.R., Zajonc, D.M., Ben-Menachem, G., Ainge, G.D., Painter, G.F., et al. (2006). Natural killer T cells recognize diacylglycerol antigens from pathogenic bacteria. Nat. Immunol. 7, 978–986.10.1038/ni1380Suche in Google Scholar PubMed
Kinjo, Y., Illarionov, P., Vela, J.L., Pei, B., Girardi, E., Li, X., Li, Y., Imamura, M., Kaneko, Y., Okawara, A., et al. (2011). Invariant natural killer T cells recognize glycolipids from pathogenic Gram-positive bacteria. Nat. Immunol. 12, 966–974.10.1038/ni.2096Suche in Google Scholar PubMed PubMed Central
Kitamura, H., Iwakabe, K., Yahata, T., Nishimura, S., Ohta, A., Ohmi, Y., Sato, M., Takeda, K., Okumura, K., Van Kaer, L., et al. (1999). The natural killer T (NKT) cell ligand α-galactosylceramide demonstrates its immunopotentiating effect by inducing interleukin (IL)-12 production by dendritic cells and IL-12 receptor expression on NKT cells. J. Exp. Med. 189, 1121–1128.10.1084/jem.189.7.1121Suche in Google Scholar PubMed PubMed Central
Kobayashi, E., Motoki, K., Uchida, T., Fukushima, H., and Koezuka, Y. (1995). KRN7000, a novel immunomodulator, and its antitumor activities. Oncol. Res. 7, 529–534.Suche in Google Scholar
Koh, Y.I., Kim, H.Y., Meyer, E.H., Pichavant, M., Akbari, O., Yasumi, T., Savage, P.B., Dekruyff, R.H., and Umetsu, D.T. (2008). Activation of nonclassical CD1d-restricted NK T cells induces airway hyperreactivity in β2-microglobulin-deficient mice. J. Immunol. 181, 4560–4569.10.4049/jimmunol.181.7.4560Suche in Google Scholar PubMed PubMed Central
Kolter, T. and Sandhoff, K. (2010). Lysosomal degradation of membrane lipids. FEBS Lett. 584, 1700–1712.10.1016/j.febslet.2009.10.021Suche in Google Scholar PubMed
Kotas, M.E., Lee, H.Y., Gillum, M.P., Annicelli, C., Guigni, B.A., Shulman, G.I., and Medzhitov, R. (2011). Impact of CD1d deficiency on metabolism. PLoS One 6, e25478.10.1371/journal.pone.0025478Suche in Google Scholar PubMed PubMed Central
Leslie, D.S., Dascher, C.C., Cembrola, K., Townes, M.A., Hava, D.L., Hugendubler, L.C., Mueller, E., Fox, L., Roura-Mir, C., Moody, D.B., et al. (2008). Serum lipids regulate dendritic cell CD1 expression and function. Immunology 125, 289–301.10.1111/j.1365-2567.2008.02842.xSuche in Google Scholar PubMed PubMed Central
Liao, C.M., Zimmer, M.I., Shanmuganad, S., Yu, H.T., Cardell, S.L., and Wang, C.R. (2012). dysregulation of CD1d-restricted type II natural killer T cells leads to spontaneous development of colitis in mice. Gastroenterology 142, 326–34 e1–2.10.1053/j.gastro.2011.10.030Suche in Google Scholar PubMed PubMed Central
Lin, Y., Roberts, T.J., Spence, P.M., and Brutkiewicz, R.R. (2005). Reduction in CD1d expression on dendritic cells and macrophages by an acute virus infection. J. Leukoc. Biol. 77, 151–158.10.1189/jlb.0704399Suche in Google Scholar PubMed
Lopez-Sagaseta, J., Sibener, L.V., Kung, J.E., Gumperz, J., and Adams, E.J. (2012). Lysophospholipid presentation by CD1d and recognition by a human natural killer T-cell receptor. EMBO J. 31, 2047–2059.10.1038/emboj.2012.54Suche in Google Scholar PubMed PubMed Central
Lotter, H., Gonzalez-Roldan, N., Lindner, B., Winau, F., Isibasi, A., Moreno-Lafont, M., Ulmer, A.J., Holst, O., Tannich, E., and Jacobs, T. (2009). Natural killer T cells activated by a lipopeptidophosphoglycan from Entamoeba histolytica are critically important to control amebic liver abscess. PLoS Pathog 5, e1000434.10.1371/journal.ppat.1000434Suche in Google Scholar PubMed PubMed Central
Ly, D., Kasmar, A.G., Cheng, T.Y., de Jong, A., Huang, S., Roy, S., Bhatt, A., van Summeren, R.P., Altman, J.D., Jacobs, W.R., Jr., et al. (2013). CD1c tetramers detect ex vivo T cell responses to processed phosphomycoketide antigens. J. Exp. Med. 210, 729–741.10.1084/jem.20120624Suche in Google Scholar PubMed PubMed Central
Lynch, L., O’Shea, D., Winter, D.C., Geoghegan, J., Doherty, D.G., and O’Farrelly, C. (2009). Invariant NKT cells and CD1d+ cells amass in human omentum and are depleted in patients with cancer and obesity. Eur. J. Immunol. 39, 1893–1901.10.1002/eji.200939349Suche in Google Scholar PubMed
Lynch, L., Nowak, M., Varghese, B., Clark, J., Hogan, A.E., Toxavidis, V., Balk, S.P., O’Shea, D., O’Farrelly, C., and Exley, M.A. (2012). Adipose tissue invariant NKT cells protect against diet-induced obesity and metabolic disorder through regulatory cytokine production. Immunity 37, 574–587.10.1016/j.immuni.2012.06.016Suche in Google Scholar PubMed PubMed Central
Mantell, B.S., Stefanovic-Racic, M., Yang, X., Dedousis, N., Sipula, I.J., and O’Doherty, R.M. (2011). Mice lacking NKT cells but with a complete complement of CD8+ T-cells are not protected against the metabolic abnormalities of diet-induced obesity. PLoS One 6, e19831.10.1371/journal.pone.0019831Suche in Google Scholar PubMed PubMed Central
Matsuda, J.L., Naidenko, O.V., Gapin, L., Nakayama, T., Taniguchi, M., Wang, C.R., Koezuka, Y., and Kronenberg, M. (2000). Tracking the response of natural killer T cells to a glycolipid antigen using CD1d tetramers. J. Exp. Med. 192, 741–754.10.1084/jem.192.5.741Suche in Google Scholar PubMed PubMed Central
Mattner, J., Debord, K.L., Ismail, N., Goff, R.D., Cantu, C., 3rd, Zhou, D., Saint-Mezard, P., Wang, V., Gao, Y., Yin, N., et al. (2005). Exogenous and endogenous glycolipid antigens activate NKT cells during microbial infections. Nature 434, 525–529.10.1038/nature03408Suche in Google Scholar PubMed
Metelitsa, L.S., Wu, H.W., Wang, H., Yang, Y., Warsi, Z., Asgharzadeh, S., Groshen, S., Wilson, S.B., and Seeger, R.C. (2004). Natural killer T cells infiltrate neuroblastomas expressing the chemokine CCL2. J. Exp. Med. 199, 1213–1221.10.1084/jem.20031462Suche in Google Scholar PubMed PubMed Central
Moll, M., Kuylenstierna, C., Gonzalez, V.D., Andersson, S.K., Bosnjak, L., Sonnerborg, A., Quigley, M.F., and Sandberg, J.K. (2009). Severe functional impairment and elevated PD-1 expression in CD1d-restricted NKT cells retained during chronic HIV-1 infection. Eur. J. Immunol. 39, 902–911.10.1002/eji.200838780Suche in Google Scholar PubMed PubMed Central
Molling, J.W., Langius, J.A., Langendijk, J.A., Leemans, C.R., Bontkes, H.J., van der Vliet, H.J., von Blomberg, B.M., Scheper, R.J., and van den Eertwegh, A.J. (2007). Low levels of circulating invariant natural killer T cells predict poor clinical outcome in patients with head and neck squamous cell carcinoma. J. Clin. Oncol. 25, 862–868.10.1200/JCO.2006.08.5787Suche in Google Scholar PubMed
Montoya, C.J., Jie, H.B., Al-Harthi, L., Mulder, C., Patino, P.J., Rugeles, M.T., Krieg, A.M., Landay, A.L., and Wilson, S.B. (2006). Activation of plasmacytoid dendritic cells with TLR9 agonists initiates invariant NKT cell-mediated cross-talk with myeloid dendritic cells. J. Immunol. 177, 1028– 1039.10.4049/jimmunol.177.2.1028Suche in Google Scholar PubMed
Moody, D.B., Briken, V., Cheng, T.Y., Roura-Mir, C., Guy, M.R., Geho, D.H., Tykocinski, M.L., Besra, G.S., and Porcelli, S.A. (2002). Lipid length controls antigen entry into endosomal and nonendosomal pathways for CD1b presentation. Nat. Immunol. 3, 435–442.10.1038/ni780Suche in Google Scholar PubMed
Moody, D.B., Young, D.C., Cheng, T.Y., Rosat, J.P., Roura-Mir, C., O’Connor, P.B., Zajonc, D.M., Walz, A., Miller, M.J., Levery, S.B., et al. (2004). T cell activation by lipopeptide antigens. Science 303, 527–531.10.1126/science.1089353Suche in Google Scholar PubMed
Moody, D.B., Zajonc, D.M., and Wilson, I.A. (2005). Anatomy of CD1-lipid antigen complexes. Nat. Rev. Immunol. 5, 387–399.10.1038/nri1605Suche in Google Scholar PubMed
Motohashi, S., Ishikawa, A., Ishikawa, E., Otsuji, M., Iizasa, T., Hanaoka, H., Shimizu, N., Horiguchi, S., Okamoto, Y., Fujii, S., et al. (2006). A phase I study of in vitro expanded natural killer T cells in patients with advanced and recurrent non-small cell lung cancer. Clin. Cancer Res. 12, 6079–6086.10.1158/1078-0432.CCR-06-0114Suche in Google Scholar PubMed
Motohashi, S., Nagato, K., Kunii, N., Yamamoto, H., Yamasaki, K., Okita, K., Hanaoka, H., Shimizu, N., Suzuki, M., Yoshino, I., et al. (2009). A phase I-II study of alpha-galactosylceramide-pulsed IL-2/GM-CSF-cultured peripheral blood mononuclear cells in patients with advanced and recurrent non-small cell lung cancer. J. Immunol. 182, 2492–2501.10.4049/jimmunol.0800126Suche in Google Scholar PubMed
Motoki, K., Maeda, K., Ueno, H., Kobayashi, E., Uchida, T., Fukushima, H., and Koezuka, Y. (1996). Antitumor activities of combined treatment with a novel immunomodulator, (2S,3S,4R)-1-O-(α-d-galactopyranosyl)-2-(N-hexacosanoylamino)-1,3,4-octadecanetriol (KRN7000), and radiotherapy in tumor-bearing mice. Oncol. Res. 8, 155–162.Suche in Google Scholar
Muindi, K., Cernadas, M., Watts, G.F., Royle, L., Neville, D.C., Dwek, R.A., Besra, G.S., Rudd, P.M., Butters, T.D., and Brenner, M.B. (2010). Activation state and intracellular trafficking contribute to the repertoire of endogenous glycosphingolipids presented by CD1d [corrected]. Proc. Natl. Acad. Sci. USA 107, 3052–3057.10.1073/pnas.0915056107Suche in Google Scholar PubMed PubMed Central
Nagarajan, N.A. and Kronenberg, M. (2007). Invariant NKT cells amplify the innate immune response to lipopolysaccharide. J. Immunol. 178, 2706–2713.10.4049/jimmunol.178.5.2706Suche in Google Scholar PubMed
Nagato, K., Motohashi, S., Ishibashi, F., Okita, K., Yamasaki, K., Moriya, Y., Hoshino, H., Yoshida, S., Hanaoka, H., Fujii, S., et al. (2012). Accumulation of activated invariant natural killer T cells in the tumor microenvironment after α-galactosylceramide-pulsed antigen presenting cells. J. Clin. Immunol. 32, 1071–1081.10.1007/s10875-012-9697-9Suche in Google Scholar
Odyniec, A.N., Barral, D.C., Garg, S., Tatituri, R.V., Besra, G.S., and Brenner, M.B. (2010). Regulation of CD1 antigen-presenting complex stability. J. Biol. Chem. 285, 11937–11947.10.1074/jbc.M109.077933Suche in Google Scholar
O’Konek, J.J., Illarionov, P., Khursigara, D.S., Ambrosino, E., Izhak, L., Castillo, B.F., 2nd, Raju, R., Khalili, M., Kim, H.Y., Howell, A.R., et al. (2011). Mouse and human iNKT cell agonist β-mannosylceramide reveals a distinct mechanism of tumor immunity. J. Clin. Invest. 121, 683–694.10.1172/JCI42314Suche in Google Scholar
Olszak, T., An, D., Zeissig, S., Vera, M.P., Richter, J., Franke, A., Glickman, J.N., Siebert, R., Baron, R.M., Kasper, D.L., et al. (2012). Microbial exposure during early life has persistent effects on natural killer T cell function. Science 336, 489–493.10.1126/science.1219328Suche in Google Scholar
Orange, J.S. (2002). Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 4, 1545–1558.10.1016/S1286-4579(02)00038-2Suche in Google Scholar
Paduraru, C., Bezbradica, J.S., Kunte, A., Kelly, R., Shayman, J.A., Veerapen, N., Cox, L.R., Besra, G.S., and Cresswell, P. (2013). Role for lysosomal phospholipase A2 in iNKT cell-mediated CD1d recognition. Proc. Natl. Acad. Sci. USA 110, 5097–5102.10.1073/pnas.1302923110Suche in Google Scholar PubMed PubMed Central
Paget, C., Mallevaey, T., Speak, A.O., Torres, D., Fontaine, J., Sheehan, K.C., Capron, M., Ryffel, B., Faveeuw, C., Leite de Moraes, M., et al. (2007). Activation of invariant NKT cells by toll-like receptor 9-stimulated dendritic cells requires type I interferon and charged glycosphingolipids. Immunity 27, 597–609.10.1016/j.immuni.2007.08.017Suche in Google Scholar PubMed
Park, S.H., Roark, J.H., and Bendelac, A. (1998). Tissue-specific recognition of mouse CD1 molecules. J. Immunol. 160, 3128–3134.10.4049/jimmunol.160.7.3128Suche in Google Scholar
Park, S.H., Weiss, A., Benlagha, K., Kyin, T., Teyton, L., and Bendelac, A. (2001). The mouse CD1d-restricted repertoire is dominated by a few autoreactive T cell receptor families. J. Exp. Med. 193, 893–904.10.1084/jem.193.8.893Suche in Google Scholar PubMed PubMed Central
Park, J.J., Kang, S.J., De Silva, A.D., Stanic, A.K., Casorati, G., Hachey, D.L., Cresswell, P., and Joyce, S. (2004). Lipid-protein interactions: biosynthetic assembly of CD1 with lipids in the endoplasmic reticulum is evolutionarily conserved. Proc. Natl. Acad. Sci. USA 101, 1022–1026.10.1073/pnas.0307847100Suche in Google Scholar PubMed PubMed Central
Patel, O., Pellicci, D.G., Gras, S., Sandoval-Romero, M.L., Uldrich, A.P., Mallevaey, T., Clarke, A.J., Le Nours, J., Theodossis, A., Cardell, S.L., et al. (2012). Recognition of CD1d-sulfatide mediated by a type II natural killer T cell antigen receptor. Nat. Immunol. 13, 857–863.10.1038/ni.2372Suche in Google Scholar PubMed
Pei, B., Speak, A.O., Shepherd, D., Butters, T., Cerundolo, V., Platt, F.M., and Kronenberg, M. (2011). Diverse endogenous antigens for mouse NKT cells: self-antigens that are not glycosphingolipids. J. Immunol. 186, 1348–1360.10.4049/jimmunol.1001008Suche in Google Scholar PubMed PubMed Central
Pellicci, D.G., Clarke, A.J., Patel, O., Mallevaey, T., Beddoe, T., Le Nours, J., Uldrich, A.P., McCluskey, J., Besra, G.S., Porcelli, S.A., et al. (2011). Recognition of β-linked self glycolipids mediated by natural killer T cell antigen receptors. Nat. Immunol. 12, 827–833.10.1038/ni.2076Suche in Google Scholar PubMed PubMed Central
Planell, N., Lozano, J.J., Mora-Buch, R., Masamunt, M.C., Jimeno, M., Ordás, I., Esteller, M., Ricart, E., Piqué, J.M., Panés, J., et al. (2013). Transcriptional analysis of the intestinal mucosa of patients with ulcerative colitis in remission reveals lasting epithelial cell alterations. Gut 62, 967–976.10.1136/gutjnl-2012-303333Suche in Google Scholar PubMed
Porcelli, S., Brenner, M.B., Greenstein, J.L., Balk, S.P., Terhorst, C., and Bleicher, P.A. (1989). Recognition of cluster of differentiation 1 antigens by human CD4-CD8-cytolytic T lymphocytes. Nature 341, 447–450.10.1038/341447a0Suche in Google Scholar PubMed
Porcelli, S., Morita, C.T., and Brenner, M.B. (1992). CD1b restricts the response of human CD4–8-T lymphocytes to a microbial antigen. Nature 360, 593–597.10.1038/360593a0Suche in Google Scholar PubMed
Prigozy, T.I., Naidenko, O., Qasba, P., Elewaut, D., Brossay, L., Khurana, A., Natori, T., Koezuka, Y., Kulkarni, A., and Kronenberg, M. (2001). Glycolipid antigen processing for presentation by CD1d molecules. Science 291, 664–667.10.1126/science.291.5504.664Suche in Google Scholar PubMed
Raftery, M.J., Winau, F., Kaufmann, S.H., Schaible, U.E., and Schonrich, G. (2006). CD1 antigen presentation by human dendritic cells as a target for herpes simplex virus immune evasion. J. Immunol. 177, 6207–6214.10.4049/jimmunol.177.9.6207Suche in Google Scholar PubMed
Raftery, M.J., Winau, F., Giese, T., Kaufmann, S.H., Schaible, U.E., and Schonrich, G. (2008). Viral danger signals control CD1d de novo synthesis and NKT cell activation. Eur. J. Immunol. 38, 668–679.10.1002/eji.200737233Suche in Google Scholar PubMed
Raghuraman, G., Geng, Y., and Wang, C.R. (2006). IFN-beta-mediated up-regulation of CD1d in bacteria-infected APCs. J. Immunol. 177, 7841–7848.10.4049/jimmunol.177.11.7841Suche in Google Scholar PubMed
Relloso, M., Cheng, T.Y., Im, J.S., Parisini, E., Roura-Mir, C., DeBono, C., Zajonc, D.M., Murga, L.F., Ondrechen, M.J., Wilson, I.A., et al. (2008). pH-dependent interdomain tethers of CD1b regulate its antigen capture. Immunity 28, 774–786.10.1016/j.immuni.2008.04.017Suche in Google Scholar PubMed PubMed Central
Renukaradhya, G.J., Webb, T.J., Khan, M.A., Lin, Y.L., Du, W., Gervay-Hague, J., and Brutkiewicz, R.R. (2005). Virus-induced inhibition of CD1d1-mediated antigen presentation: reciprocal regulation by p38 and ERK. J. Immunol. 175, 4301–4308.10.4049/jimmunol.175.7.4301Suche in Google Scholar PubMed
Renukaradhya, G.J., Khan, M.A., Shaji, D., and Brutkiewicz, R.R. (2008). Vesicular stomatitis virus matrix protein impairs CD1d-mediated antigen presentation through activation of the p38 MAPK pathway. J. Virol. 82, 12535–12542.10.1128/JVI.00881-08Suche in Google Scholar PubMed PubMed Central
Rhost, S., Lofbom, L., Rynmark, B.M., Pei, B., Mansson, J.E., Teneberg, S., Blomqvist, M., and Cardell, S.L. (2012). Identification of novel glycolipid ligands activating a sulfatide-reactive, CD1d-restricted, type II natural killer T lymphocyte. Eur. J. Immunol. 42, 2851–2860.10.1002/eji.201142350Suche in Google Scholar PubMed
Richter, J., Neparidze, N., Zhang, L., Nair, S., Monesmith, T., Sundaram, R., Miesowicz, F., Dhodapkar, K.M., and Dhodapkar, M.V. (2013). Clinical regressions and broad immune activation following combination therapy targeting human NKT cells in myeloma. Blood 121, 423–430.10.1182/blood-2012-06-435503Suche in Google Scholar PubMed PubMed Central
Rossjohn, J., Pellicci, D.G., Patel, O., Gapin, L., and Godfrey, D.I. (2012). Recognition of CD1d-restricted antigens by natural killer T cells. Nat. Rev. Immunol. 12, 845–857.10.1038/nri3328Suche in Google Scholar PubMed PubMed Central
Roy, K.C., Maricic, I., Khurana, A., Smith, T.R., Halder, R.C., and Kumar, V. (2008). Involvement of secretory and endosomal compartments in presentation of an exogenous self-glycolipid to type II NKT cells. J. Immunol. 180, 2942–2950.10.4049/jimmunol.180.5.2942Suche in Google Scholar PubMed
Salio, M., Speak, A.O., Shepherd, D., Polzella, P., Illarionov, P.A., Veerapen, N., Besra, G.S., Platt, F.M., and Cerundolo, V. (2007). Modulation of human natural killer T cell ligands on TLR-mediated antigen-presenting cell activation. Proc. Natl. Acad. Sci. USA 104, 20490–20495.10.1073/pnas.0710145104Suche in Google Scholar PubMed PubMed Central
Sanchez, D.J., Gumperz, J.E., and Ganem, D. (2005). Regulation of CD1d expression and function by a herpesvirus infection. J. Clin. Invest. 115, 1369–1378.10.1172/JCI200524041Suche in Google Scholar
Santos, C.R. and Schulze, A. (2012). Lipid metabolism in cancer. FEBS J. 279, 2610–2623.10.1111/j.1742-4658.2012.08644.xSuche in Google Scholar PubMed
Saubermann, L.J., Beck, P., De Jong, Y.P., Pitman, R.S., Ryan, M.S., Kim, H.S., Exley, M., Snapper, S., Balk, S.P., Hagen, S.J., et al. (2000). Activation of natural killer T cells by α-galactosylceramide in the presence of CD1d provides protection against colitis in mice. Gastroenterology 119, 119–128.10.1053/gast.2000.9114Suche in Google Scholar PubMed
Scharf, L., Li, N.S., Hawk, A.J., Garzon, D., Zhang, T., Fox, L.M., Kazen, A.R., Shah, S., Haddadian, E.J., Gumperz, J.E., et al. (2010). The 2.5 Å structure of CD1c in complex with a mycobacterial lipid reveals an open groove ideally suited for diverse antigen presentation. Immunity 33, 853–862.10.1016/j.immuni.2010.11.026Suche in Google Scholar PubMed PubMed Central
Schiechl, G., Bauer, B., Fuss, I., Lang, S.A., Moser, C., Ruemmele, P., Rose-John, S., Neurath, M.F., Geissler, E.K., Schlitt, H.J., et al. (2011). Tumor development in murine ulcerative colitis depends on MyD88 signaling of colonic F4/80+CD11bhighGr1low macrophages. J. Clin. Invest. 121, 1692–1708.10.1172/JCI42540Suche in Google Scholar PubMed PubMed Central
Schipper, H.S., Rakhshandehroo, M., van de Graaf, S.F., Venken, K., Koppen, A., Stienstra, R., Prop, S., Meerding, J., Hamers, N., Besra, G., et al. (2012). Natural killer T cells in adipose tissue prevent insulin resistance. J. Clin. Invest. 122, 3343–3354.10.1172/JCI62739Suche in Google Scholar PubMed PubMed Central
Seok, J., Warren, H.S., Cuenca, A.G., Mindrinos, M.N., Baker, H.V., Xu, W., Richards, D.R., McDonald-Smith, G.P., Gao, H., Hennessy, L., et al. (2013). Genomic responses in mouse models poorly mimic human inflammatory diseases. Proc. Natl. Acad. Sci. USA 110, 3507–3512.10.1073/pnas.1222878110Suche in Google Scholar PubMed PubMed Central
Seshadri, C., Shenoy, M., Wells, R.D., Hensley-McBain, T., Andersen-Nissen, E., McElrath, M.J., Cheng, T.Y., Moody, D.B., and Hawn, T.R. (2013). Human CD1a deficiency is common and genetically regulated. J. Immunol. 191, 1586–1593.10.4049/jimmunol.1300575Suche in Google Scholar PubMed PubMed Central
Shamshiev, A., Gober, H.J., Donda, A., Mazorra, Z., Mori, L., and De Libero, G. (2002). Presentation of the same glycolipid by different CD1 molecules. J. Exp. Med. 195, 1013–1021.10.1084/jem.20011963Suche in Google Scholar PubMed PubMed Central
Shinya, E., Owaki, A., Shimizu, M., Takeuchi, J., Kawashima, T., Hidaka, C., Satomi, M., Watari, E., Sugita, M., and Takahashi, H. (2004). Endogenously expressed HIV-1 nef down-regulates antigen-presenting molecules, not only class I MHC but also CD1a, in immature dendritic cells. Virology 326, 79–89.10.1016/j.virol.2004.06.004Suche in Google Scholar PubMed
Shiratsuchi, T., Schneck, J., Kawamura, A., and Tsuji, M. (2009). Human CD1 dimeric proteins as indispensable tools for research on CD1-binding lipids and CD1-restricted T cells. J. Immunol. Methods 345, 49–59.10.1016/j.jim.2009.04.002Suche in Google Scholar PubMed PubMed Central
Sieling, P.A., Chatterjee, D., Porcelli, S.A., Prigozy, T.I., Mazzaccaro, R.J., Soriano, T., Bloom, B.R., Brenner, M.B., Kronenberg, M., Brennan, P.J., et al. (1995). CD1-restricted T cell recognition of microbial lipoglycan antigens. Science 269, 227–230.10.1126/science.7542404Suche in Google Scholar PubMed
Skold, M., Xiong, X., Illarionov, P.A., Besra, G.S., and Behar, S.M. (2005). Interplay of cytokines and microbial signals in regulation of CD1d expression and NKT cell activation. J. Immunol. 175, 3584–3593.10.4049/jimmunol.175.6.3584Suche in Google Scholar PubMed
Smyth, M.J., Thia, K.Y., Street, S.E., Cretney, E., Trapani, J.A., Taniguchi, M., Kawano, T., Pelikan, S.B., Crowe, N.Y., and Godfrey, D.I. (2000). Differential tumor surveillance by natural killer (NK) and NKT cells. J. Exp. Med. 191, 661–668.10.1084/jem.191.4.661Suche in Google Scholar
Song, L., Asgharzadeh, S., Salo, J., Engell, K., Wu, H.W., Sposto, R., Ara, T., Silverman, A.M., DeClerck, Y.A., Seeger, R.C., et al. (2009). Valpha24-invariant NKT cells mediate antitumor activity via killing of tumor-associated macrophages. J. Clin. Invest. 119, 1524–1536.10.1172/JCI37869Suche in Google Scholar
Sriram, V., Du, W., Gervay-Hague, J., and Brutkiewicz, R.R. (2005). Cell wall glycosphingolipids of Sphingomonas paucimobilis are CD1d-specific ligands for NKT cells. Eur. J. Immunol. 35, 1692–1701.10.1002/eji.200526157Suche in Google Scholar
Subleski, J.J., Jiang, Q., Weiss, J.M., and Wiltrout, R.H. (2011). The split personality of NKT cells in malignancy, autoimmune and allergic disorders. Immunotherapy 3, 1167–1184.10.2217/imt.11.117Suche in Google Scholar
Sugita, M., Jackman, R.M., van Donselaar, E., Behar, S.M., Rogers, R.A., Peters, P.J., Brenner, M.B., and Porcelli, S.A. (1996). Cytoplasmic tail-dependent localization of CD1b antigen-presenting molecules to MIICs. Science 273, 349–352.10.1126/science.273.5273.349Suche in Google Scholar
Sugita, M., Porcelli, S.A., and Brenner, M.B. (1997). Assembly and retention of CD1b heavy chains in the endoplasmic reticulum. J. Immunol. 159, 2358–2365.10.4049/jimmunol.159.5.2358Suche in Google Scholar
Sugita, M., Grant, E.P., van Donselaar, E., Hsu, V.W., Rogers, R.A., Peters, P.J., and Brenner, M.B. (1999). Separate pathways for antigen presentation by CD1 molecules. Immunity 11, 743–752.10.1016/S1074-7613(00)80148-XSuche in Google Scholar
Sugita, M., van Der Wel, N., Rogers, R.A., Peters, P.J., and Brenner, M.B. (2000). CD1c molecules broadly survey the endocytic system. Proc. Natl. Acad. Sci. USA 97, 8445–8450.10.1073/pnas.150236797Suche in Google Scholar PubMed PubMed Central
Swann, J.B., Uldrich, A.P., van Dommelen, S., Sharkey, J., Murray, W.K., Godfrey, D.I., and Smyth, M.J. (2009). Type I natural killer T cells suppress tumors caused by p53 loss in mice. Blood 113, 6382–6385.10.1182/blood-2009-01-198564Suche in Google Scholar PubMed PubMed Central
Tachibana, T., Onodera, H., Tsuruyama, T., Mori, A., Nagayama, S., Hiai, H., and Imamura, M. (2005). Increased intratumor Vα24-positive natural killer T cells: a prognostic factor for primary colorectal carcinomas. Clin. Cancer Res. 11, 7322–7327.10.1158/1078-0432.CCR-05-0877Suche in Google Scholar PubMed
Tahir, S.M., Cheng, O., Shaulov, A., Koezuka, Y., Bubley, G.J., Wilson, S.B., Balk, S.P., and Exley, M.A. (2001). Loss of IFN-γ production by invariant NK T cells in advanced cancer. J. Immunol. 167, 4046–4050.10.4049/jimmunol.167.7.4046Suche in Google Scholar PubMed
Tatituri, R.V., Watts, G.F., Bhowruth, V., Barton, N., Rothchild, A., Hsu, F.F., Almeida, C.F., Cox, L.R., Eggeling, L., Cardell, S., et al. (2013). Recognition of microbial and mammalian phospholipid antigens by NKT cells with diverse TCRs. Proc. Natl. Acad. Sci. USA 110, 1827–1832.10.1073/pnas.1220601110Suche in Google Scholar PubMed PubMed Central
Terabe, M., Matsui, S., Noben-Trauth, N., Chen, H., Watson, C., Donaldson, D.D., Carbone, D.P., Paul, W.E., and Berzofsky, J.A. (2000). NKT cell-mediated repression of tumor immunosurveillance by IL-13 and the IL-4R-STAT6 pathway. Nat. Immunol. 1, 515–520.10.1038/82771Suche in Google Scholar PubMed
Terabe, M., Matsui, S., Park, J.M., Mamura, M., Noben-Trauth, N., Donaldson, D.D., Chen, W., Wahl, S.M., Ledbetter, S., Pratt, B., et al. (2003). Transforming growth factor β production and myeloid cells are an effector mechanism through which CD1d-restricted T cells block cytotoxic T lymphocyte-mediated tumor immunosurveillance: abrogation prevents tumor recurrence. J. Exp. Med. 198, 1741–1752.10.1084/jem.20022227Suche in Google Scholar PubMed PubMed Central
Tremaroli, V. and Backhed, F. (2012). Functional interactions between the gut microbiota and host metabolism. Nature 489, 242–249.10.1038/nature11552Suche in Google Scholar PubMed
Tupin, E., Kinjo, Y., and Kronenberg, M. (2007). The unique role of natural killer T cells in the response to microorganisms. Nat. Rev. Microbiol. 5, 405–417.10.1038/nrmicro1657Suche in Google Scholar PubMed
Tyznik, A.J., Tupin, E., Nagarajan, N.A., Her, M.J., Benedict, C.A., and Kronenberg, M. (2008). Cutting edge: the mechanism of invariant NKT cell responses to viral danger signals. J. Immunol. 181, 4452–4456.10.4049/jimmunol.181.7.4452Suche in Google Scholar PubMed PubMed Central
Uchida, T., Horiguchi, S., Tanaka, Y., Yamamoto, H., Kunii, N., Motohashi, S., Taniguchi, M., Nakayama, T., and Okamoto, Y. (2008). Phase I study of alpha-galactosylceramide-pulsed antigen presenting cells administration to the nasal submucosa in unresectable or recurrent head and neck cancer. Cancer Immunol. Immunother 57, 337–345.10.1007/s00262-007-0373-5Suche in Google Scholar PubMed
Uldrich, A.P., Patel, O., Cameron, G., Pellicci, D.G., Day, E.B., Sullivan, L.C., Kyparissoudis, K., Kjer-Nielsen, L., Vivian, J.P., Cao, B., et al. (2011). A semi-invariant Vα10+ T cell antigen receptor defines a population of natural killer T cells with distinct glycolipid antigen-recognition properties. Nat. Immunol. 12, 616–623.10.1038/ni.2051Suche in Google Scholar PubMed PubMed Central
Van Rhijn, I., Kasmar, A., de Jong, A., Gras, S., Bhati, M., Doorenspleet, M.E., de Vries, N., Godfrey, D.I., Altman, J.D., de Jager, W., et al. (2013). A conserved human T cell population targets mycobacterial antigens presented by CD1b. Nat. Immunol. 14, 706–713.10.1038/ni.2630Suche in Google Scholar PubMed PubMed Central
Vincent, M.S., Xiong, X., Grant, E.P., Peng, W., and Brenner, M.B. (2005). CD1a-, b-, and c-restricted TCRs recognize both self and foreign antigens. J. Immunol. 175, 6344–6351.10.4049/jimmunol.175.10.6344Suche in Google Scholar PubMed
Vivier, E., Ugolini, S., Blaise, D., Chabannon, C., and Brossay, L. (2012). Targeting natural killer cells and natural killer T cells in cancer. Nat. Rev. Immunol. 12, 239–252.10.1038/nri3174Suche in Google Scholar PubMed PubMed Central
von Bernuth, H., Picard, C., Jin, Z., Pankla, R., Xiao, H., Ku, C.L., Chrabieh, M., Mustapha, I.B., Ghandil, P., Camcioglu, Y., et al. (2008). Pyogenic bacterial infections in humans with MyD88 deficiency. Science 321, 691–696.10.1126/science.1158298Suche in Google Scholar PubMed PubMed Central
Wetterau, J.R., Aggerbeck, L.P., Bouma, M.E., Eisenberg, C., Munck, A., Hermier, M., Schmitz, J., Gay, G., Rader, D.J., and Gregg, R.E. (1992). Absence of microsomal triglyceride transfer protein in individuals with abetalipoproteinemia. Science 258, 999–1001.10.1126/science.1439810Suche in Google Scholar PubMed
Winau, F., Schwierzeck, V., Hurwitz, R., Remmel, N., Sieling, P.A., Modlin, R.L., Porcelli, S.A., Brinkmann, V., Sugita, M., Sandhoff, K., et al. (2004). Saposin C is required for lipid presentation by human CD1b. Nat. Immunol. 5, 169–174.10.1038/ni1035Suche in Google Scholar PubMed
Wingender, G., Stepniak, D., Krebs, P., Lin, L., McBride, S., Wei, B., Braun, J., Mazmanian, S.K., and Kronenberg, M. (2012). Intestinal microbes affect phenotypes and functions of invariant natural killer T cells in mice. Gastroenterology 143, 418–428.10.1053/j.gastro.2012.04.017Suche in Google Scholar PubMed PubMed Central
Wong, C.H., Jenne, C.N., Lee, W.Y., Leger, C., and Kubes, P. (2011). Functional innervation of hepatic iNKT cells is immunosuppressive following stroke. Science 334, 101–105.10.1126/science.1210301Suche in Google Scholar PubMed
Wu, L., Parekh, V.V., Gabriel, C.L., Bracy, D.P., Marks-Shulman, P.A., Tamboli, R.A., Kim, S., Mendez-Fernandez, Y.V., Besra, G.S., Lomenick, J.P., et al. (2012). Activation of invariant natural killer T cells by lipid excess promotes tissue inflammation, insulin resistance, and hepatic steatosis in obese mice. Proc. Natl. Acad. Sci. USA 109, E1143–E1152.10.1073/pnas.1200498109Suche in Google Scholar PubMed PubMed Central
Yanagisawa, K., Seino, K., Ishikawa, Y., Nozue, M., Todoroki, T., and Fukao, K. (2002). Impaired proliferative response of Vα24 NKT cells from cancer patients against α-galactosylceramide. J. Immunol. 168, 6494–6499.10.4049/jimmunol.168.12.6494Suche in Google Scholar PubMed
Yang, L., Jhaveri, R., Huang, J., Qi, Y., and Diehl, A.M. (2007). Endoplasmic reticulum stress, hepatocyte CD1d and NKT cell abnormalities in murine fatty livers. Lab. Invest. 87, 927–937.10.1038/labinvest.3700603Suche in Google Scholar PubMed
Yoshioka, K., Ueno, Y., Tanaka, S., Nagai, K., Onitake, T., Hanaoka, R., Watanabe, H., and Chayama, K. (2012). Role of natural killer T cells in the mouse colitis-associated colon cancer model. Scand. J. Immunol. 75, 16–26.10.1111/j.1365-3083.2011.02607.xSuche in Google Scholar PubMed
Yu, E.D., Girardi, E., Wang, J., and Zajonc, D.M. (2011). Cutting edge: structural basis for the recognition of β-linked glycolipid antigens by invariant NKT cells. J. Immunol. 187, 2079–2083.10.4049/jimmunol.1101636Suche in Google Scholar PubMed PubMed Central
Yuan, W., Dasgupta, A., and Cresswell, P. (2006). Herpes simplex virus evades natural killer T cell recognition by suppressing CD1d recycling. Nat. Immunol. 7, 835–842.10.1038/ni1364Suche in Google Scholar PubMed
Yuan, W., Qi, X., Tsang, P., Kang, S.J., Illarionov, P.A., Besra, G.S., Gumperz, J., and Cresswell, P. (2007). Saposin B is the dominant saposin that facilitates lipid binding to human CD1d molecules. Proc. Natl. Acad. Sci. USA 104, 5551–5556.10.1073/pnas.0700617104Suche in Google Scholar PubMed PubMed Central
Yuan, W., Kang, S.J., Evans, J.E., and Cresswell, P. (2009). Natural lipid ligands associated with human CD1d targeted to different subcellular compartments. J. Immunol. 182, 4784–4791.10.4049/jimmunol.0803981Suche in Google Scholar PubMed PubMed Central
Zajonc, D.M., Crispin, M.D., Bowden, T.A., Young, D.C., Cheng, T.Y., Hu, J., Costello, C.E., Rudd, P.M., Dwek, R.A., Miller, M.J., et al. (2005). Molecular mechanism of lipopeptide presentation by CD1a. Immunity 22, 209–219.10.1016/j.immuni.2004.12.009Suche in Google Scholar PubMed
Zeissig, S. and Blumberg, R.S. (2012). Primary immunodeficiency associated with defects in CD1 and CD1-restricted T cells. Ann. NY Acad Sci 1250, 14–24.10.1111/j.1749-6632.2011.06380.xSuche in Google Scholar PubMed
Zeissig, S., Kaser, A., Dougan, S.K., Nieuwenhuis, E.E., and Blumberg, R.S. (2007). Role of NKT cells in the digestive system. III. Role of NKT cells in intestinal immunity. Am. J. Physiol. Gastrointest. Liver Physiol. 293, G1101–G1105.Suche in Google Scholar
Zeissig, S., Dougan, S.K., Barral, D.C., Junker, Y., Chen, Z., Kaser, A., Ho, M., Mandel, H., McIntyre, A., Kennedy, S.M., et al. (2010). Primary deficiency of microsomal triglyceride transfer protein in human abetalipoproteinemia is associated with loss of CD1 function. J. Clin. Invest. 120, 2889–2899.10.1172/JCI42703Suche in Google Scholar PubMed PubMed Central
Zeissig, S., Murata, K., Sweet, L., Publicover, J., Hu, Z., Kaser, A., Bosse, E., Iqbal, J., Hussain, M.M., Balschun, K., et al. (2012). Hepatitis B virus-induced lipid alterations contribute to natural killer T cell-dependent protective immunity. Nat. Med. 18, 1060–1068.10.1038/nm.2811Suche in Google Scholar PubMed PubMed Central
Zhang, S.Y., Jouanguy, E., Ugolini, S., Smahi, A., Elain, G., Romero, P., Segal, D., Sancho-Shimizu, V., Lorenzo, L., Puel, A., et al. (2007). TLR3 deficiency in patients with herpes simplex encephalitis. Science 317, 1522–1527.10.1126/science.1139522Suche in Google Scholar PubMed
Zhou, D., Cantu, C., 3rd, Sagiv, Y., Schrantz, N., Kulkarni, A.B., Qi, X., Mahuran, D.J., Morales, C.R., Grabowski, G.A., Benlagha, K., et al. (2004a). Editing of CD1d-bound lipid antigens by endosomal lipid transfer proteins. Science 303, 523–527.10.1126/science.1092009Suche in Google Scholar PubMed PubMed Central
Zhou, D., Mattner, J., Cantu, C., 3rd, Schrantz, N., Yin, N., Gao, Y., Sagiv, Y., Hudspeth, K., Wu, Y.P., Yamashita, T., et al. (2004b). Lysosomal glycosphingolipid recognition by NKT cells. Science 306, 1786–1789.10.1126/science.1103440Suche in Google Scholar PubMed
Zimmer, M.I., Nguyen, H.P., Wang, B., Xu, H., Colmone, A., Felio, K., Choi, H.J., Zhou, P., Alegre, M.L., and Wang, C.R. (2009). Polymorphisms in CD1d affect antigen presentation and the activation of CD1d-restricted T cells. Proc. Natl. Acad. Sci. USA 106, 1909–1914.10.1073/pnas.0808476106Suche in Google Scholar PubMed PubMed Central
©2014 by Walter de Gruyter Berlin Boston
Artikel in diesem Heft
- Masthead
- Masthead
- Reviews
- Stressed to death – mechanisms of ER stress-induced cell death
- Granule proteases of hematopoietic cells, a family of versatile inflammatory mediators – an update on their cleavage specificity, in vivo substrates, and evolution
- Unraveling RNA-mediated networks: new insights from new technologies
- Lipid antigens in immunity
- Research Articles/Short Communications
- Protein Structure and Function
- Differential contributions of the latch in Thermotoga maritima reverse gyrase to the binding of single-stranded DNA before and after ATP hydrolysis
- Proteolysis
- Clinical value of protein expression of kallikrein-related peptidase 7 (KLK7) in ovarian cancer
- Amyloid β peptide cleavage by kallikrein 7 attenuates fibril growth and rescues neurons from Aβ-mediated toxicity in vitro
Artikel in diesem Heft
- Masthead
- Masthead
- Reviews
- Stressed to death – mechanisms of ER stress-induced cell death
- Granule proteases of hematopoietic cells, a family of versatile inflammatory mediators – an update on their cleavage specificity, in vivo substrates, and evolution
- Unraveling RNA-mediated networks: new insights from new technologies
- Lipid antigens in immunity
- Research Articles/Short Communications
- Protein Structure and Function
- Differential contributions of the latch in Thermotoga maritima reverse gyrase to the binding of single-stranded DNA before and after ATP hydrolysis
- Proteolysis
- Clinical value of protein expression of kallikrein-related peptidase 7 (KLK7) in ovarian cancer
- Amyloid β peptide cleavage by kallikrein 7 attenuates fibril growth and rescues neurons from Aβ-mediated toxicity in vitro