Updates on the classification and numbers of marine fungi
-
Mark S. Calabon
Mark S. Calabon obtained a PhD (Biological Sciences) degree from the Center of Excellence in Fungal Research, Mae Fah Luang University (Thailand). He is an assistant professor in the University of the Philippines Visayas. His research focuses on the taxonomy, phylogeny, and classification of aquatic fungi.

 ,
E.B. Gareth Jones
,
E.B. Gareth Jones
Prof. E.B. Gareth Jones is a widely cited author on marine fungi, their ecology, physiology, and systematics and has co-edited several books and journal special issues. He has reported on marine fungi from around the world, in particular Asia, especially those found on mangroves. He supervised over 100 Ph.D. and a highly cited scientist.
,
Ka-Lai Pang
Ka-Lai Pang obtained his BSc and PhD degrees from the City University of Hong Kong in 1998 and 2001, respectively. Prof. Pang studies the biology of marine fungi and fungus-like organisms and endophytic fungi associated with mangrove plants and macroalgae.
,
Mohamed A. Abdel-Wahab
Mohamed A. Abdel-Wahab is a professor of marine mycology, and his research interest includes ecology and taxonomy of marine fungi, natural products from aquatic and endophytic fungi, and ecology and physiology of thraustochytrids.
,
Jing Jin
,
Bandarupalli Devadatha
,
Resurreccion B. Sadaba
,
Carlo Chris Apurillo
and
Kevin D. Hyde
Prof. Kevin D. Hyde is the director of the Center of Excellence in Fungal Research, Mae Fah Luang University. As a prolific researcher, internationally renowned mycologist, and most respected educator, he has been recognized as a highly cited scientist and distinguished mycologist by different organizations for his works in various disciplines of mycology. At present, he supervises 100+ graduate students, and is a proud mentor of his previous students, now collaborators, who became leading mycologists as educators and researchers.

Abstract
This article revises the documented diversity of known marine fungi from nine phyla, 33 classes, 107 orders, 273 families, 767 genera and 1898 species reported worldwide. A member of the Aphelidiomycota, Pseudaphelidium drebesii, is reported for the first time from the marine environment, on a diatom. Likewise, the phylum Mortierellomycota is listed following taxonomic changes, as are six subclasses: Chaetothyriomycetidae, Savoryellomycetidae, Sclerococcomycetidae, Agaricostilbomycetidae, Auriculariomycetidae and Aphelidiomycotina. Thirty-three orders and 105 family names with marine species are added to the checklist, along with 641 species in 228 genera, bringing the total to 1898. New additions of species added to the list of marine fungi are highlighted. Four new combinations are proposed: Stigmatodiscus mangrovei, Diaporthe krabiensis and Diaporthe xylocarpi, while the hyphomycete Humicola alopallonella is referred to the genus Halosphaeriopsis, as Halosphaeriopsis alopallonella.
1 Introduction
The classification of marine fungi has been continuously updated with the incorporation of molecular data in taxonomic studies. This resulted in an increase in the estimated numbers of fungi from marine environments and has been shown in series of papers on the classification of higher orders of marine fungi: 530 (Jones et al. 2009), 624 (Abdel-Wahab and Bahkali 2012), 1112 (Jones et al. 2015), 1257 (Jones et al. 2019). Since Jones et al. (2019) listed the marine fungi documented at that time, a further 641 species, 231 genera, 58 families, 14 orders, two classes and two phyla have been added. Some of these are due to nomenclature changes while the Microsporidia were not included in the earlier publications. In this manuscript we highlight the major changes that have occurred in the past four years.
Table 1 is a snapshot of marine fungi assigned to higher order taxa of the fungi, which illustrates the dominance of the Ascomycota in marine habitats. This dominance is reflected in every ecological habitat surveyed: mangroves (Devadatha et al. 2021), salt marshes (Calabon et al. 2021), and planktonic marine fungi (Hassett et al. 2020). However, parasites of marine animals are dominated by microsporidia (Bojko and Stentiford 2022; Pang et al. 2021).
Taxonomic classification of marine fungi.
| Phylum | Species | Genus | Family | Order | Class |
|---|---|---|---|---|---|
| Ascomycota | 1561 | 590 | 176 | 63 | 10 |
| Basidiomycota | 158 | 86 | 49 | 27 | 10 |
| Microsporidia | 128 | 56 | 26 | 7 | 3 |
| Chytridiomycota | 29 | 18 | 12 | 5 | 5 |
| Mucoromycota | 17 | 12 | 6 | 1 | 1 |
| Olpidiomycota | 2 | 2 | 1 | 1 | 1 |
| Aphelidiomycota | 1 | 1 | 1 | 1 | 1 |
| Blastocladiomycota | 1 | 1 | 1 | 1 | 1 |
| Mortierellomycota | 1 | 1 | 1 | 1 | 1 |
| Total | 1898 | 767 | 273 | 107 | 33 |
2 Selected notes on marine fungi
2.1 Phyla and subclasses
Since the last checklist of marine fungi in 2019, numerous novel species, genera, families, orders, and subclasses have been discovered. This can be accounted for by the sampling of a wider range of substrata (e.g., algae, sea grasses, seawater), sampling of deep-sea locations, and innovation in sampling, such as Sanger dideoxy chain-termination sequencing, high throughput sequencing (HTS) platforms 454-Pyrosequencing and Illumina (Abdel-Wahab et al. 2021; Calabon et al. 2021; Poli et al. 2020a; Vargas-Gastélum and Riquelme 2020; Varrella et al. 2021; Walker and Robicheau 2021; Wang et al. 2014, 2019). Significant changes have been made to the higher classification of the fungi as the result of a number of phylogenetic studies based on a wider range of loci sequenced and divergence time estimates (Hongsanan et al. 2020a,b; Hyde 2022; Hyde et al. 2020b; Phukhamsakda et al. 2022; Senanayake et al. 2022). Higher order classifications have been proposed by various authors from Hibbett et al. (2007) and Hyde et al. (2017) to Tedersoo et al. (2018).
Since 2019, the phylum Aphelidiomycota has been added to our survey of marine fungi while the Mortierallales has been raised in rank to Mortierellomycota (Tedersoo et al. 2018). We add six subclasses which include marine fungi: Chaetothyriomycetidae (Doweld 2001), Savoryellomycetidae (Hongsanan et al. 2017), Sclerococcomycetidae (Réblová et al. 2017), Agaricostilbomycetidae (Swann et al. 2001), Auriculariomycetidae (Jülich 1982), and Aphelidiomycotina (Tedersoo et al. 2018).
Currently, marine microsporidia total 128 species in 56 genera, parasitic primarily on crustacea, fish, less frequently on annelid worms (such as the genera Capitella, Echiurida), rarely striped dolphin (Stenella coeruleoalba), and the California sea-lion (Zalophus californanus). Microsporidia are a particular problem as the cause of diseases in aquaculture. A survey of 150 pre-stocked ponds in Thailand found the prevalence of the microsporidian Enterocytozoon hepatopenaei to be 49 %, with the resultant economic loss $32,000 per hectare (Shinn 2016). They are worldwide in distribution with Sapir et al. (2014) reporting on a new microsporidian Nematocenator marisprofundi from the deep sea, that infects benthic nematodes in methane seeps on the Pacific Ocean floor.
2.2 Orders
2.2.1 Eremithallales (Dothideomycetes, Ascomycota)
Eremithallales was introduced for a single species Eremithallus costaricensis (Lücking et al. 2008), with a divergence time estimated as 238 MYA (stem age, Hongsanan et al. 2020b). The order comprised one family Eremithallaceae which has now been synonymized under Melaspileaceae, and Eremithallus costaricensis is included in the genus Melaspilea (Ertz and Diederich 2015). One marine ascomycete has been referred to Melaspilea, namely M. mangrovei (Vrijmoed et al. 1996), but this has not been confirmed by molecular data.
2.2.2 Golubeviales (Exobasidiomycetes, Basidiomycota)
The order was introduced for the genus Golubeva with G. pallescens as the type species (Wang et al. 2015). In a multigene phylogeny and taxonomic revision of yeasts in the Ustilaginomycotina, two species in the genus Tilletiopsis (T. albescens, T. pallescens) grouped separately from the Tilletiopsis clade and these were referred to new genera, Robbauera and Golubeva, respectively. These genera served as new families and orders in the Ustilaginomycotina. Golubeva is the type genus for the Golubeviaceae, Golubeviales (Wang et al. 2015), a new name for the checklist.
2.2.3 Lobulomycetales (Lobulomycetes, Chytridiomycota)
Lobulomycetales was introduced by Simmons et al. (2009) based on genetic analyses, morphology, and ultrastructural data, with Lobulomyces (Lobulomycetaceae) and L. angularis as the type genus and species, respectively. Features of the order are a polycentric or monocentric thallus, multiple rhizoidal axes, spherical motile zoospores with or without a flagellar plug, ultrastructural details of the kinestosome, growing on chitin in soil and freshwater/seawater. Two families are assigned to the order Lobulomycetaceae and (Alogomycetaceae). Alogomyces tanneri (Alogomycetaceae) is known from marine habitats; based on environmental DNA samples from the deep ocean (Le Calvez et al. 2009; Yan et al. 2020).
2.2.4 Monoblastiales (Dothideomycetes, Ascomycota)
The genus Heleiosa was introduced by Kohlmeyer et al. (1996), for the type species H. barbatula, growing on leaves of Juncus roemerianus in salt marshes, and referred to the Dothideomycetes incertae sedis. Subsequently it was attributed to Monoblastiaceae (Monoblastiales) which includes both lichenized and non-lichenized ascomycetes (Wijayawardene et al. 2022). Monoblastiales was introduced by Hyde et al. (2013), and H. barbatula was referred to this order by Hongsanan et al. (2020b).
2.2.5 Muyocopronales (Dothideomycetes, Ascomycota)
Muyocopronaceae was invalidly introduced by Luttrell (1951) and later reintroduced by Hyde et al. (2013) while the order Muyocopronales was introduced by Mapook et al. (2016). Ellisiodothis inquinans (Muyocopronaceae) was reported from Spartina alterniflora in Buenos Aires, Argentina by Gessner and Kohlmeyer (1976), but there is no further documentation.
2.2.6 Myrmecridiales (Sordariomycetes, Ascomycota)
The order Myrmecridiales (Sordariomycetes) includes one salt marsh fungus Myrmecridium schulzeri, an asexual morph with subhyaline, obovoid or fusiform, 1-celled, conidia that can be distinguished from Ramichloridium by having hyaline mycelium, pale brown to hyaline conidiogenous cells with pimple-like denticles (Arzanlou et al. 2007). Myrmecridium schulzeri is recorded for the first time as saprobic on Distichlis spicata (saltgrass) in Buenos Aires, Argentina (Calabon et al. 2021; Elíades et al. 2007).
2.2.7 Neophaeothecales (Dothideomycetes, Ascomycota)
Phaeotheca salicorniae was introduced by Crous et al. (2016) and transferred to the new genus Neophaeotheca by Abdollahzadeh et al. (2020) and the type for the new family and order Neophaeothecaceae, and Neophaeothecales, respectively. Neophaeotheca salicorniae and Neophaeotheca triangularis are black yeasts reported from hypersaline waters in salterns (Chung et al. 2019; Gunde-Cimerman et al. 2000).
2.2.8 Pleurotheciales (Sordariomycetes, Ascomycota)
Two Phaeoisaria species are found in marine habitats: Ph. sedimenticola isolated from surface marine sediment in the inter-tidal zone, Shandong, China (Cheng et al. 2014) and Ph. clematidis in mangrove litter (Devadatha et al. 2021; Maria and Sridhar 2003). Pleurotheciaceae was introduced by Réblová et al. (2016) in Pleurotheciales (Sordariomycetes) and is included for the first time in the checklist of marine fungi.
2.2.9 Sakaguchiales (Cystobasidiomycetes, Basidiomycota)
In our 2019 checklist of marine fungi (Jones et al. 2019), we listed the Basidiomycota genus Sakaguchia in the Sakaguchiaceae and order Erythrobasdiales. Zhao et al. (2017) introduced the order Sakaguchiales and this is accepted in this revision.
2.2.10 Sclerococcales (Eurotiomycetes, Ascomycota)
Within the Eurotiomycetes, the discovery of an older name for the genus Dactylospora (Sclerococcum) has led to the introduction of a new subclass Sclerococcomycetidae for the order and family Sclerococcales, and Sclerococcaceae, respectively (Réblová et al. 2017). Four apothecial marine fungi are included in this change: Sclerococcum haliotrephum, S. mangrovei, S. vrijmoediae from mangrove wood (Jones et al. 1999; Kohlmeyer and Kohlmeyer 1965; Pang et al. 2014) and S. canariense from driftwood (Kohlmeyer and Volkmann-Kohlmeyer 1998).
2.2.11 Sporidesmiales (Sordariomycetes, Ascomycota)
Sporidesmiales was introduced by Crous et al. (2018b) with a single family (Sporidesmiaceae) and genus (Sporidesmium) saprobic on woody debris in terrestrial and aquatic habitats or mycoparasites of fungi. The divergence time for Sporidesmiales is estimated as 81 MYA (Hyde et al. 2020b), which is evidence of a family status but ranking it as an order may need revision following further study. One marine species (Sporidesmium salinum) has been reported on submerged wood at Millport, Isle of Man, UK as an asexual morph (Jones 1963). No further reports of the species are known and further collections are required to obtain sequence data and determine its taxonomic affiliation.
2.2.12 Stigmatodiscales (Dothideomycetes, Ascomycota)
Asterodiscus tamaricis was introduced by Voglmayr et al. (2016) from material collected on dead twigs of Tamarix tetrandra (Austria) but later transferred to Stigmatodiscus, S. tamaricis, by Voglmayr and Amengual (2018). A second species, Asterodiscus mangrovei was described by Dayarathne et al. (2020a) from decaying submerged wood at a mangrove strand in Thailand. Voglmayr et al. (2016) referred Stigmatodiscus to the new family and order (Stigmatodiscaceae, Stigmatodiscales) and recorded for the first time for the marine fungi checklist. In the present study, we transfer Asterodiscus mangrovei to Stigmatodiscus as S. mangrovei.
Stigmatodiscus mangrovei (Dayar., E.B.G. Jones et K.D. Hyde) M.S. Calabon, E.B.G Jones et K.L. Pang, comb. nov.
Mycobank number: MB848521.
≡ Asterodiscus mangrovei Dayar., E.B.G. Jones et K.D. Hyde, Mycosphere 11(1): 86 (2020).
Sexual morph: Descriptions and illustrations refer to Dayarathne et al. (2020a). Asexual morph: Not observed.
Distribution: THAILAND, Krabi Province, on decaying wood at a mangrove strand (Dayarathne et al. 2020a).
Notes: Holotype MFLU 16–1216. LSU (MN017861), SSU (MN017926), ITS (MN047095) sequence data are available.
2.2.13 Xenospadicoidales (Sordariomycetes, Ascomycota)
Xenospadicoidales, is a monotypic order in the class Sordariomycetes and based on the family Xenospadicoidaceae introduced by Hernández-Restrepo et al. (2017). It comprises eight genera of which Lentomitella has been reported from the marine environment, although generally known from wood in terrestrial and freshwater habitats (Ding et al. 2011). Members of Xenospadicoidales were reported from various marine substrata (Ding et al. 2011).
2.3 Families
Many of the new family names listed in this study are due to additional species reported from the marine environment.
2.3.1 Amorosiaceae
Amorosiaceae (Pleosporales) was introduced by Thambugala and Hyde (in Thambugala et al. 2015) for the monotypic hyphomycete genus Amorosia (type species A. littorali) isolated from intertidal sediment, associated with Rhizophora, in the Bahamas (Mantle et al. 2006). Conidia are solitary, dry, lateral, elongate-clavate, pale brown to brown, 1–3-septate, 3–4-septate when mature, constricted at the septa, smooth-walled, and are lacking any mucilaginous sheath or appendages. Previously assigned to Sporomiaceae, Thambugala et al. (2015), in a multi-gene phylogenetic analysis, showed it grouped with Angustimassarina in a well-supported clade, in between a clade comprising Decaisnella formosa, Thyridaria macrostomoides and a new genus, Lignosphaeria and Sporormiaceae. They therefore introduced a new family for the two genera Amorosia and Angustimassarina. It is the source of three new chlorinated compounds: two propenylphenol derivatives, chlorophenol A and B (1 and 2), and one benzofuran derivative, chlorophenol C (3) and other compounds (Ren et al. 2022).
2.3.2 Cordieritidaceae
Fryar et al. (2019) introduced the genus Annabella with A. australiensis as the type species collected on decaying decorticated branches of Avicennia marina in South Australia and referred it to the Cordieritidaceae (Helotiales) in the subclass Leotiomycetidae (Leotiomycetes). This is the first marine species record of this subclass introduced by Kirk et al. (2008) and the family Cordieritidaceae. Annabella australiensis differs from other taxa within Cordieritidaceae in having hyaline to yellowish perithecioid apothecia, contrasting with the usually dark-coloured apothecia of other members of the family (Fryar et al. 2019). Due to this absence of perithecoid pigment, Annabella australiensis differs from other taxa in the family. Ascospores are hyaline, vary in shape from ellipsoid to fusoid to oblong, are aseptate, and lack appendages or sheaths.
2.3.3 Cyphellophoraceae
Cyphellophoraceae, typified by Cyphellophora laciniata, was introduced by Réblová et al. (2013). Presently, only one species, Cyphellophora europaea, has been reported from deep-sea hydrothermal vents, deep-sea, and deep subsurface sediments using culture-based approaches (Xu et al. 2017).
2.3.4 Cylindrosympodiaceae
Shen et al. (2020) introduced the family Cylindrosympodiaceae in the Venturiales (Dothideomycetes) to accommodate the type species Cylindrosympodium variabile. A number of genera are assigned to the family: Cylindrosympodium, Pseudoanungitea, Sympodiella, Tothia and Septonema, with Septonema strictum (=Taeniolella stricta) reported from marine habitats (Sridhar and Maria 2003, 2006). Septonema was previously assigned to the Mytilinidiaceae and is polyphyletic with many species awaiting sequencing (Crous et al. 2007b).
2.3.5 Lachnaceae
In the 2019 checklist, Lachnum spartinae was classified under Hyaloscyphaceae, but here we correct this and refer it to the new family Lachnaceae introduced by Raitviir (2004).
2.3.6 Linocarpaceae
Konta et al. (2017) introduced Linocarpaceae, typified by Linocarpon pandani, to accommodate Linocarpon and Neolinocarpon. In marine habitats, two species have been recorded, Linocarpon livistonae and L. pandani (Hawksworth 2000; Hyde 1988a,b, 1989; Hyde and Alias 2000). This is a new family introduction to the marine fungi checklist.
2.3.7 Lophiotremataceae
Lophiotrema rubi is recorded from collections made from algae and seagrass (Gnavi et al. 2017; Panno et al. 2013) and is referred in the present work to family Lophiotremataceae introduced by Hirayama and Tanaka (2011), based on morphological characters and molecular phylogenetic data using LSU and SSU sequence data. This is a new family introduction to the marine fungi checklist.
2.3.8 Myrotheciomycetaceae
Jones et al. (2019) referred marine Emericellopsis species to the Hypocreales incertae sedis, but Crous et al. (2018b) introduced the family Myrotheciomycetaceae (Hypocreales, Sordariomycetes) for this genus, and this is followed here. Seven novel Emericellopsis species have been introduced in the last decade from a variety of substrata: E. atlantica on Stelletta normani in Ireland, E. cladophorae on the green alga Cladophora, E. phycophila both from Portugal (Gonçalves et al. 2020; Hagestad et al. 2021). This brings the number of marine Emericellopsis to twelve. Hagestad et al. (2021) sequenced the genome of an Emericellopsis sp. TS7 from a marine origin.
2.3.9 Nigrogranaceae
Nigrogranaceae was introduced by Jaklitsch and Voglmayr (2016) for the genus Nigrograna and two novel species Ni. hydei and Ni. obtusispora, based on phylogenetic analyses of a combined ITS-TEF1α sequence dataset. The genus includes two marine species: Ni. samueliana on decaying wood of Avicennia marina, and Ni. rhizophorae, on submerged propagule of Rhizophora sp. They are recorded for the first time from mangrove habitats (Hyde et al. 2020b)
2.3.10 Phaeoseptaceae
Phaeoseptaceae (Pleosporales) was introduced by Hyde et al. (2018) for Phaeoseptum which was previously placed in Halotthiaceae (Zhang et al. 2013). Phaeoseptum is a monotypic genus and includes two marine species: P. carolshearerianum on decaying wood of Avicennia marina, and P. manglicola on decaying wood of Avicennia marina and Suaeda monoica (Dayarathne et al. 2020a). This is the first record of the family in the marine fungi checklist.
2.3.11 Pleomassariaceae
Pleomassariaceae (Pleosporales) was introduced with Pleomassaria as the type by Barr (1979) and to include fungi characterized by distoseptate, dark brown ascospores and ascomatal walls which are thickened at sides and thin at the base. The family includes Splanchnonema with the taxon S. britzelmayrianum reported for the first time from the marine environment. A new family record for the marine fungi checklist.
2.3.12 Pseudoastrosphaeriellaceae
Phookamsak et al. (2015) introduced the family Pseudoastrosphaeriellaceae to accommodate a single genus Pseudoastrosphaeriella for taxa originally described in the genus Astrosphaeriella (Hawksworth and Boise 1985). Hyde et al. (2017) accommodated the two marine species of Carinispora in Pseudoastrosphaeriellaceae based on morphology and phylogenetic analyses. Later, Phookamsak et al. (2019) introduced a third genus, Pseudoastrosphaeriellopsis with one new species from mangroves in India. Currently the family includes three genera and 10 species, of which three taxa were described from marine habitats.
2.3.13 Saccotheciaceae
Jones et al. (2019) referred Aureobasidium pullulans (collected in the marine environment) to the family Dothioraceae, while Thambugala et al. (2014) included it in the new family Aureobasidiaceae, a family already introduced by Ciferri (1958). However, an earlier name for Aureobasidiaceae is Saccotheciaceae, established by Bonorden (1864) to accommodate Saccothecium, and is considered as a distinct family in Dothidelaes. Saccotheciaceae can be distinguished from Dothideaceae by immersed to erumpent, uniloculate ascostromata and aseptate to multi-septate, hyaline ascospores (Hongsanan et al. 2020a). Currently seven genera are accepted in Saccotheciaceae with three marine species: Aureobasidium leucospermi, A. pullulans, and A. melanogenum.
2.3.14 Stigmatodiscaceae
Voglmayr et al. (2016) established the family Stigmatodiscaceae to accommodate two genera: Asterodiscus and Stigmatodiscus. However, Voglmayr and Amengual (2018) synonymized the two genera based on indistinguishable morphology and molecular characteristics. Currently, Stigmatodiscaceae contains Stigmatodiscus with seven new species. Stigmatodiscus touroultii was described from Salvadora angustifolia, a saline tree (Poncet et al. 2022). Dayarathne et al. (2020a) described Asterodiscus mangrovei from mangroves in Thailand. In the present study, Asterodiscus mangrovei is transferred to Stigmatodiscus as S. mangrovei (see notes in Stigmatodiscales).
2.3.15 Striatiguttulaceae
Zhang et al. (2019b) introduced two new genera Longicorpus and Striatiguttula in the new pleosporalean family Striatiguttulaceae, collected on mangrove plants. A multi-gene dataset (LSU, SSU, TEF-1α, RPB2) showed that the two genera formed a monophyletic clade (Striatiguttulaceae) and a new lineage in the Pleosporales, and close to Ligninsphaeriaceae, Pseudoastrosphaeriellaceae, Testudinaceae, and Tetraplosphaeriaceae. Two new species of Striatiguttula nypae and S. phoenicis were collected on Nypa fruticans and Phoenix paludosa, respectively, and characterized by black, immersed ascomata with a short papilla, and ascospores that are hyaline to brown, fusiform or ellipsoidal, 1–3-septate, striate, with paler end cells and surrounded by a mucilaginous sheath. Longicorpus is based on Trematosphaeria striataspora introduced by Hyde (1988a) and later transferred to Astrosphaeriella striataspora (Hyde 1992) but formed a distinct well supported clade in Striatiguttulaceae, and characterized by black ascomata that are immersed to erumpent, clypeate, with a long neck, and ascospores hyaline to brown, fusiform, upper middle cell slightly swollen towards the central septum, and the end cells paler and smaller, striate, surrounded by a mucilaginous sheath. Longicorpus striataspora was found on two mangrove palm species, Nypa fruticans and Phoenix paludosa. A divergence time estimate analysis indicated the crown age to be 39 (20–63) MYA and stem age 60 (35–91) MYA for Striatiguttulaceae (Zhang et al. 2019b).
2.3.16 Teratosphaeriaceae
Crous et al. (2007a) introduced Teratosphaeriaceae to accommodate Teratosphaeria (with a Readeriella-like asexual morph) and 11 asexual genera. The family comprises around 60 genera and represents one of the largest families in Dothideomycetes (Crous et al. 2011c; Wijayawardene et al. 2022). The members of the family are saprobes, plant and human pathogens, rock-inhabiting, and endophytes (Crous et al. 2009a,b, 2011a,b; Egidi et al. 2014; Quaedvlieg et al. 2014). Hortaea werneckii (Teratosphaeriaceae) is a very common taxon in marine habitats (Abdel-Wahab et al. 2014).
2.3.17 Thyridariaceae
Thyridariaceae was introduced by Hyde et al. (2013) with Thyridaria incrustans as the type genus and species, in the Pleosporales, Dothideomycetes. Thyridaria previously had been referred to some six different families. Three genera in Thyridariaceae have marine species: Thyridariella, Parathyridaria and Pseudothyridariella. Devadatha et al. (2018) introduced the new genus Thyridariella with two new marine species (T. mangrovei, T. mahakoshae) on decaying wood of Avicennia marina from India. Based on phylogeny of 18S, 28S, ITS rRNA, RPB2, and TEF-1α genes, Thyridariella formed a sister relationship with a clade comprising Thyridaria and Parathyridaria in the Thyridariaceae. Thyridariella has ascomata with ostiolar necks thickened laterally, hyaline, and centrally constricted muriform ascospores with a single longitudinal septum in each segment and surrounded by a mucilaginous sheath (Devadatha et al. 2018). Parathyridaria differs in having pale to grayish brown, rarely muriform ascospores and one half of the spores is slightly larger than the other half (Jaklitsch and Voglmayr 2016). Thyridaria differs from Thyridariella mainly in ascospore morphology, i.e., phragmosporus in Thyridaria without a sheath, while muriform in Thyridariella with a sheath (Devadatha et al. 2018; Jaklitsch and Voglmayr 2016). Thyridariella mahakoshae was subsequently transferred to the new genus Pseudothyridariella by Mapook et al. (2020) as Ps. mahakoshae. Three further marine species were referred to Thyridariaceae with the introduction of the genus Parathyridariella with Pa. dematiacea as the type species, found on the green alga Flabellia petiolata and the seagrass Posidonia oceanica. Parathyridaria tyrrhenica was described from the brown alga Padina pavonica and Flabellia petiolata; and Pa. flabelliae from F. petiolata (Poli et al. 2020b). Unfortunately, all were described from mycelial cultures without sexual and asexual morphology.
2.3.18 Trichomeriaceae
Chomnunti et al. (2012) in a phylogenetic study, showed that the Trichomerium species (type species T. coffeicola), form a monophyletic clade within Chaetothyriales and established the new family Trichomeriaceae. Members of Trichomerium are generally foliar epiphytes (sooty moulds), mostly occurring on the surface of living leaves as ascostromata with setae with dark brown mycelia (Chomnunti et al. 2012). Genera assigned to the family include: Bradymyces, Lithohypha, Neostrelitziana, and Knufa. Knufia is a dematiaceous hyphomycetous genus introduced by Hutchison et al. (1995) with K. cryptophialidica as the type species. Two marine Knufa species are known: Knufia petricola isolated from the green marine alga Flabellia petiolata (Gnavi et al. 2017); K. epidermidis isolated from deep sea sediments (Wei et al. 2018).
2.3.19 Trimorphomycetaceae
Trimorphomycetaceae is a family in the order Tremellales (Basidiomycota) and currently contains four genera: Carlosrosaea, Saitozyma, Sugitazyma and Trimorphomyces, the type genus, and most are only known from their yeast states (Liu et al. 2015). Torula flava was introduced by Saito (1922) and proposed as a new genus Saitozyma in Liu et al. (2015) with five species. Saitozyma podzolica (syn. Cryptococcus podzolicus) is a yeast reported from the marine environment. Hoondee et al. (2019) and Kunthiphun et al. (2019) have isolated S. podzolica from sediment in mangroves of Thailand. The species is of interest as it is an oleaginous yeast yielding single cell oil and gluconic acid.
2.3.20 Zygosporiaceae
Li et al. (2017) introduced the Zygosporiaceae as a new family in the Xylariales (Sordariomycetes) based on phylogenetic analyses of a combined LSU and ITS sequence data generated under maximum likelihood and Bayesian criteria that showed Zygosporium species forming a strongly supported monophyletic lineage basal to the Coniocessiaceae. Zygosporiaceae had been previously introduced by Locquin (1984) but was invalidly published (Nom. inval., Art. 39.1, Melbourne). The type species of Zygosporium is Z. oscheoides (Montagne 1842) with 23 currently accepted species. Zygosporium gibbum has been reported from the marine environment on cellulosic substrata in salt marshes (Fell and Hunter 1979) and mangroves (Sarma and Vittal 2000; Vittal and Sarma 2006). Zygosporium gibbum is characterised by stalked vesicular conidiophores produced directly from the superficial mycelium, and spherical, hyaline, smooth to finely verruculose conidia (Whitton et al. 2003).
2.4 Notes on marine fungal genera
2.4.1 Diaporthe
Dayarathne et al. (2020a) described three marine Diaporthe species: D. krabiensis, D. marina, D. salinicola, from mangrove wood. The scientific name Diaporthe salinicola is preoccupied by an earlier fungus (Spegazzini 1912), and so the new ‘Diaporthe salinicola’ was invalidly described (Nom. illegit., Art. 53.1). Diaporthe krabiensis, a species isolated from submerged wood of Bruguiera sp., was published without nomenclatural repository identifier (Nom. inval., Art. F.5.1 [Shenzhen]). In the present work, we validly publish Diaporthe salinicola and Diaporthe krabiensis.
Diaporthe xylocarpi (Dayar.) M.S. Calabon et E.B.G. Jones, comb. nov.
Mycobank number: MB848445.
Basionym: Diaporthe salinicola Dayar., Mycosphere 11(1): 91 (2020).
Etymology: Name referring to the host genus Xylocarpus.
Sexual morph: Descriptions and illustrations refer to Dayarathne et al. (2020a). Asexual morph: Not observed.
Distribution: THAILAND, Ranong Province, on decaying wood of Xylocarpus sp. (Dayarathne et al. 2020a).
Notes: Holotype MFLU 18–0553. ITS (MN047098, MN047099) and TEF-1α (MN077073, MN077074) sequence data are available.
Diaporthe krabiensis (Dayar.) M.S. Calabon et E.B.G. Jones, comb. nov.
Mycobank number: MB848522.
Basionym: Diaporthe krabiensis Dayar., Mycosphere 11(1): 92 (2020).
Sexual morph: Not observed; Asexual morph: Descriptions and illustrations refer to Dayarathne et al. (2020a).
Distribution: THAILAND, Krabi Province, on decaying submerged wood of Bruguiera sp. (Dayarathne et al. 2020a).
Notes: Holotype MFLU 17–2618. ITS (MN047100, MN047101), TEF-1α (MN433215), β-tubulin tub2 (MN431495), and from Krabi, Thailand.
2.4.2 Parasarocladium
Summerbell et al. (2018) introduced the genus Parasarocladium to accommodate three Acremonium species: P. breve, P. gamsii and P. radiatum that formed a distinct clade based on 18S, ITS, 28S rDNA and actin genes. Currently, Parasarocladium comprises nine species listed in the Index Fungorum (April 2023) and MycoBank databases. The nine species were isolated from soil (five species), one species from decaying leaves and three as endophytes from marine algae. Crous et al. (2018a) erected the family Sarocladiaceae based on multigene phylogenies to accommodate the genera: Parasarocladium and Sarocladium and described Parasarocladium debruynii from a soil sample in the Netherlands. Gonçalves et al. (2019) described three new marine Parasarocladium species P. aestuarinum, P. alavariense and P. fusiforme as endophytes of macroalgae (Enteromorpha, Fucus, Ulva and unidentified Rhodophyta), sea water and sponges that were collected from Ria de Aveiro estuary, Portugal based on ITS and β-tubulin (tub2) and actin (act1) genes. Crous et al. (2020) recorded for the first time the sexual stage of the new species Parasarocladium tasmanniae that was recorded from leaves of Tasmannia insipida collected from Limpinwood Nature Reserve, New South Wales, Australia. The new species was described based on morphology and ITS, LSU, actA, TEF-1α and tub2 genes. Crous et al. (2021) described Parasarocladium wereldwijsianum from a soil sample collected from Utrecht Province, the Netherlands. Parasarocladium wereldwijsianum differs from the phylogenetically related species P. aestuarinum and P. alavariense by having olivaceous buff colonies, irregularly branched conidiophores, and hyaline, smooth-walled, ellipsoidal to obovoid, with truncate base.
The three species were described as endophytes of the marine algae (Fucus, Ulva), sea water and sponges. Parasarocladium aestuarinum is phylogenetically closely related to P. alavariense, however, both species differ in conidial morphology and dimensions with 0.013 p-distance of nucleotide sites among the ITS and β-tubulin genes. Parasarocladium fusiforme formed a basal clade to the other eight species of Parasarocladium (Crous et al. 2021; Gonçalves et al. 2020).
2.4.3 Sclerococcum
Traditionally Sclerococcum comprised lichenized species that produce sporodochial asexual morphs (Diederich et al. 2013; Hawksworth 1975, 1979), while Dactylospora species are sexual fungi that produce black apothecial ascomata with asci covered with an external, amyloid apical gelatinous cap (Hafellner 1979). Molecular phylogenetics of the type species of Dactylospora, D. parasitica, placed it in a monophyletic clade with the type species of Sclerococcum, Sclerococcum sphaerale, and the two genera were merged into the older genus name Sclerococcum under the family Dactylosporaceae and the recently described Sclerococcales (Diederich et al. 2018; Pino-Bodas et al. 2017; Réblová et al. 2017). Diederich et al. (2018) listed sixty-four Sclerococcum species and Joshi (2021) described two new species of Sclerococcum from India, so that 66 species of the genus are known worldwide. Of these, four species were recorded from marine habitats: S. canariense, S. haliotrephum, S. mangrovei, S. vrijmoediae. Marine species are saprobic and recorded from driftwood and decaying, submerged parts of the mangroves (Jones et al. 1999; Kohlmeyer 1967; Kohlmeyer and Kohlmeyer 1965; Pang et al. 2014).
Nine discomycete species have been previously described from the marine environment, of which four belong to the genus Sclerococcum. Apothecia of the four Sclerococcum species are similar, however, they differ in the ascospore shape, ornamentation, and the presence/absence of appendages (Jones et al. 1999; Kohlmeyer and Kohlmeyer 1965; Kohlmeyer and Volkmann-Kohlmeyer 1998; Pang et al. 2014). Ascospores of Sclerococcum haliotrephum are ellipsoidal with uneven cells; while they are ellipsoidal to subglobose in S. vrijmoediae and elongate-ellipsoidal in both Sclerococcum mangrovei and Sclerococcum canariense. Ascospores of Sclerococcum canariense and Sclerococcum vrijmoediae are appendaged, while they are ornamented in S. haliotrephum and S. mangrovei.
Kohlmeyer (1967) recorded Sclerococcum canariense from driftwood collected from Tenerife, Canary Islands and misidentified it as Banhegyia uralensis. Kohlmeyer and Volkmann-Kohlmeyer (1998) described the taxon in Dactylospora as D. canariensis as the second marine member of the genus. Olariaga et al. (2019) placed all non-lichenized Dactylospora species with clavate to cylindrical asci covered with an amyloid gelatinous apical cap in Sclerococcum including D. canariensis. The molecular data of S. canariense are unknown and the sequences of relevant genes are needed in order to confirm its taxonomic placement.
2.4.4 Microascus
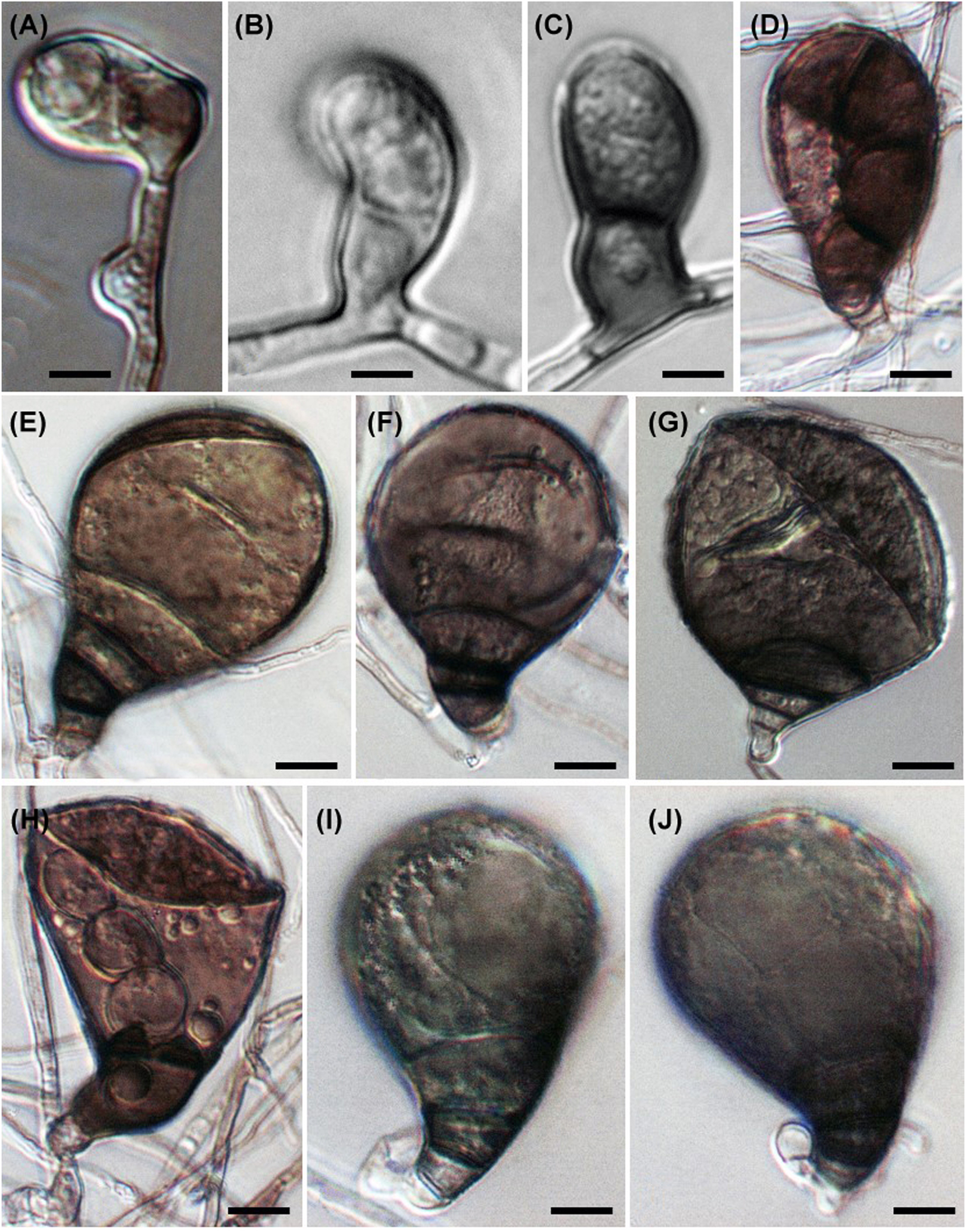
Six Microascus species have been reported from marine habitats, isolated from seawater, sponges, corals and the deep sea (see https://marinefungi.org). We illustrate two species Microascus cinereus and M. trigonosporus (Figures 1 and 2) isolated from intertidal wood collected from the coasts of the Yellow Sea and Bohai Sea, China, that are new records for marine fungi in China (Jin et al. 2004, 2005).
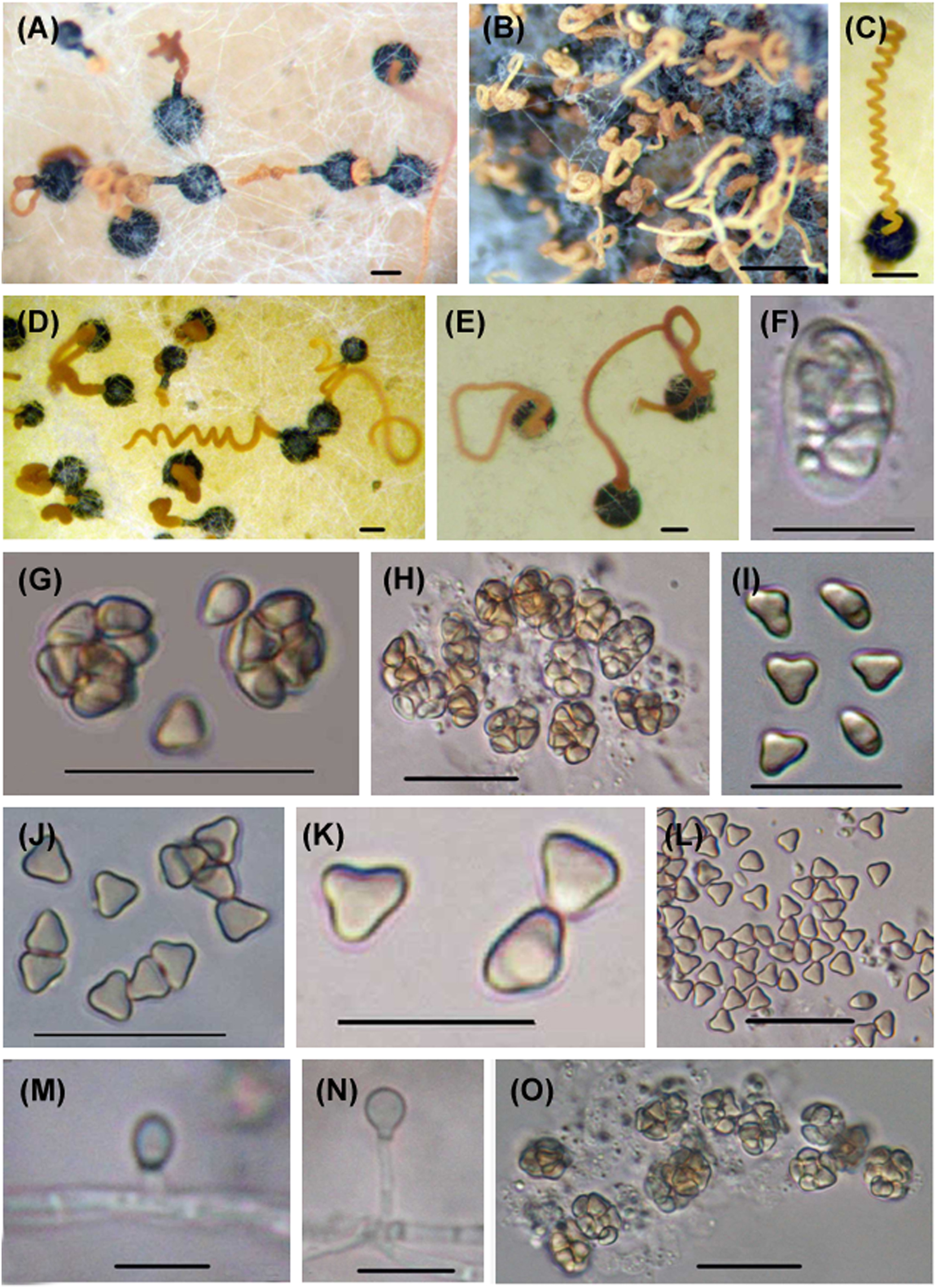
Microascus trigonosporus. (A–E) Perithecia with spore cirrus. (F) Young ascus. (G, H, O) Mature asci. (I–L) Ascospores. (M, N) Anamorphic Scopulariopsis trigonosporus of Microascus trigonosporus. Scale bars: (A–E) = 100 μm, (G, H, O) = 20 μm, (F, I–N) = 10 μm.
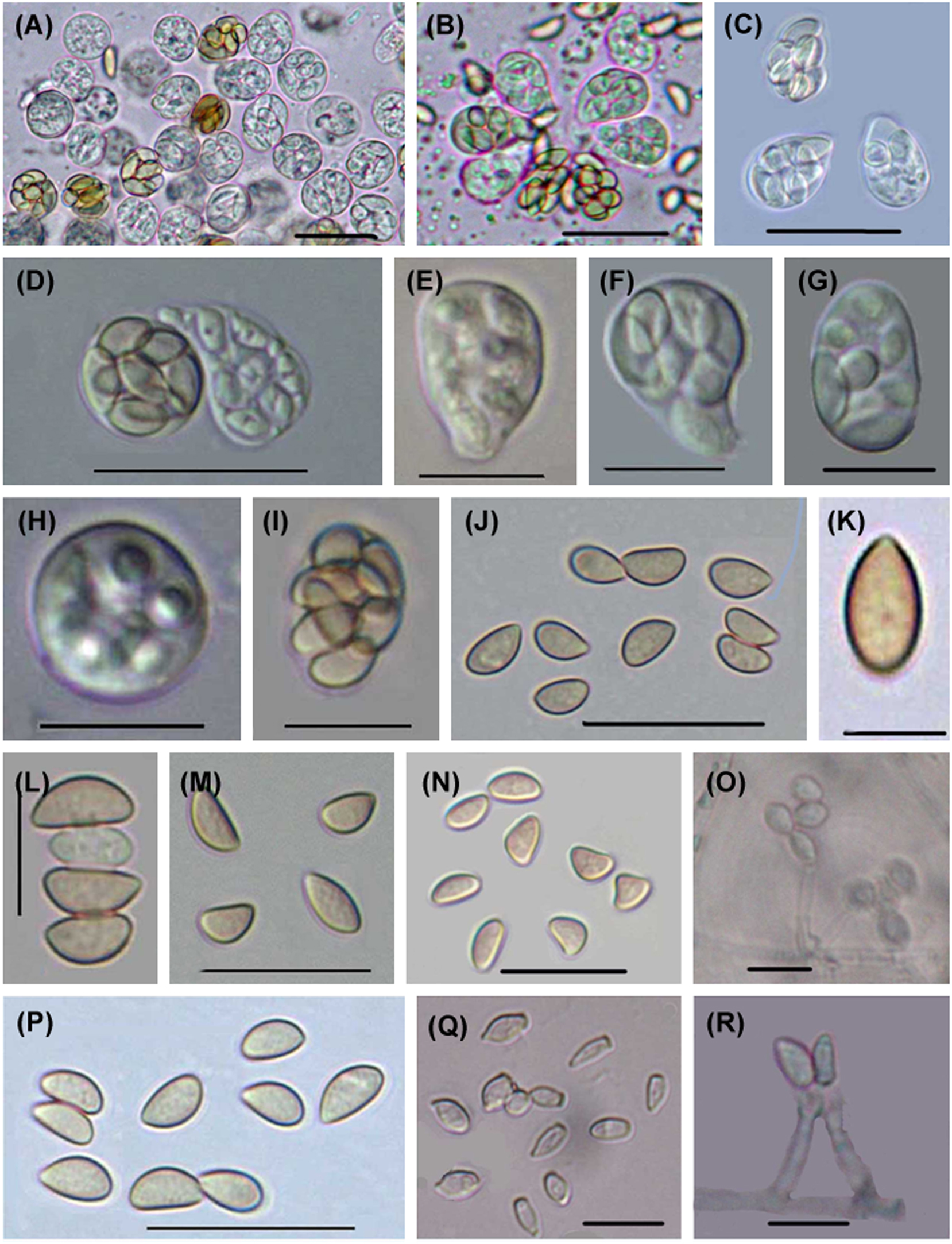
Microascus cinereus. (A–I) Asci. (J–N, P) Ascospores. (O, Q, R) Anamorphic Scopulariopsis cinerea of Microascus cinereus. Scale bars: (A–D, K, M, P–Q) = 20 μm, (E–J, L, N–O, R) = 10 μm.
2.5 Notes on marine fungal species
2.5.1 Asteromyces cruciatus
Asteromyces cruciatus was introduced by Moreau and Moreau (1941) from sand dunes of the Atlantic Coast of France. This species has a worldwide distribution and reported from a wide range of substrata (Jones and Ward 1973). Despite its profuse growth in culture and its wide occurrence in the marine milieu, its taxonomic position has remained unresolved. Asteromyces cruciatus was classified as incertae sedis in Index Fungorum and Species Fungorum, while Mycobank classified it under the old family name Dematiaceae. In a new study, Torres-Garcia et al. (2022) placed A. cruciatus in the Pleosporaceae based on a phylogenetic analysis of five gene markers (ITS, LSU, gapdh, rbp2, and tef1-α). Asteromyces is in a well-supported distinct clade with two marine Paradendryphiella species as its sister taxa, and closely related to both Paradendryphiella and Stemphylium, but they are markedly different in morphology. Brown (1958), Hennebert (1962), Kohlmeyer and Kohlmeyer (1964–1969), and Jones (1968), all report the conidia of A. cruciatus as ovoid or pyriform, smooth, thin-walled, one-celled without germ slit or pore, 10–20 × 4–9 µm, with up to 13 µm conidia developing in basipetal succession, singly on cylindrical denticles 3–6 × 0.5–1 µm. The first formed conidium is apical and retains this position. On germination, conidia often become 1-septate and these have been illustrated by Kohlmeyer and Kohlmeyer (1964–1969), Hennebert (1962), and Nolan (1972), who also reports 2-septate conidia. Jones and Ward (1973) investigated the effect of light on the sporulation of A. cruciatus: continuous illumination and in total darkness when conidia were unicellular. However, when subjected to black light (fluorescent lamp with a continuous spectrum maximum at 360 nm, 18 in. above plates), 1–6-septate conidia were formed depending on the growth media. Asteromyces cruciatus is also of interest because it can tolerate concentrations of up to 2.0 M NaCl in liquid media (Jones and Jennings 1965), with tolerance increased to 2.5 M NaCl with the addition of 0.05 M CaCl2. Asteromyces cruciatus is beautifully illustrated in Kohlmeyer and Kohlmeyer (1964–1969).
2.5.2 Biatriospora borsei
Biatriospora, typified by B. marina, was described as a sexual genus from mangrove roots in Seychelles with fusiform, 1–4 septate, brown to dark brown ascospores with hyaline polar chambers which release mucilage (Hyde and Borse 1986). Sequences of B. marina from a collection made in Singapore placed this species basal to Lophiostomataceae, Sporormiaceae and a group of unclassified taxa in a phylogenetic analysis of 18S, 28S rDNA, RPB2 and TEF-1α (Suetrong et al. 2009).
The genus Nigrograna was established to accommodate N. mackinnonii, a species originally described in Pyrenochaeta (de Gruyter et al. 2013). Hyde et al. (2013) established a new family Biatriosporaceae to accommodate Biatriospora. In a phylogenetic analysis using the same genes, Ahmed et al. (2014) found that N. mackinnonii grouped with B. marina, and thus referred the former species to Biatriospora. Later, four more species of Biatriospora were described from woody plants as endophytes: B. antibiotica, B. carollii, B. yasuniana and B. peruviensis (Kolařík et al. 2017), unaware that a new family Nigrogranaceae was established to accommodate Nigrograna with N. mackinnonii as the type, and the new combination of Melanomma fuscidulum in Nigrograna and the description of the new species N. mycophila, N. norvegica and N. obliqua (Jaklitsch and Voglmayr 2016). Jaklitsch and Voglmayr (2016) voiced doubts on the identification of the Biatriospora marina isolate and the validity of the Biatriosporaceae. Consequently, Kolařík (2018) accepted the genus Nigrograna and the family Nigrogranaceae, and transferred B. antibiotica, B. carollii, B. yasuniana and B. peruviensis to Nigrograna. After these revisions, Biatriospora became a monotypic genus but B. borsei was described as a new species in Hongsanan et al. (2020a). Biatriospora borsei clustered with B. marina (with a good support) within a group of Nigrograna spp. based on 18S, 28S, ITS rDNA, RPB2 and TEF-1α, although B. borsei is a hyphomycete with no known sexual stage. As discussed here, fresh isolates of B. marina should be collected to clarify the taxonomic position of this species, B. borsei, the genus Nigrograna and the families Biatriosporaceae and Nigrogranaceae.
2.5.3 Coniochaeta krabiensis
Coniochaeta krabiensis formed a monophyletic group with C. tetraspora and another new species (C. arenariae) in the same study (Dayarathne et al. 2020a). Morphologically, C. krabiensis fits within the generic description of Coniochaeta with its two-layered peridium, cylindrical asci with an apical apparatus and dark brown, ellipsoidal ascospores with a germ slit which are arranged uniseriately in the asci (García et al. 2006). Coniochaeta krabiensis differs from C. tetraspora in having longer, narrower, 8-spored asci and smaller ascospores (Cain 1961).
2.5.4 Cryptosphaeria avicenniae and C. halophila
Cryptosphaeria avicenniae and C. halophila were described as new species from the marine environment based on phylogenetic analyses of ITS rDNA and from India, based on ITS and β-tubulin genes for C. avicenniae and of 18S and 28S rDNA for C. halophila (Dayarathne et al. 2020b). However, the different trees shown in the study suggested that Cryptosphaeria was not a monophyletic genus. Cryptosphaeria avicenniae is morphologically similar to Cryptosphaeria bathurstensis in terms of both the sexual and asexual stages but differs in the morphology of the asci and ascospore (Dayarathne et al. 2020b; Hyde and Rappaz 1993). Cryptosphaeria halophila is an asexual fungus which is morphologically similar to C. avicenniae. The phylogenetic position of C. halophila was unstable in different analyses based on 18S and 28S rDNA genes (Dayarathne et al. 2020b).
2.5.5 Halosphaeriopsis alopallonella (Meyers et R.T. Moore) Devadatha et E.B.G. Jones, comb. nov.
Basionym: Humicola alopallonella Meyers et R.T. Moore, Am. J. Bot. 47: 346 (1960).
Synonymy: Trichocladium alopallonellum (Meyers et R.T. Moore) Kohlm. et Volkm.-Kohlm., Mycotaxon 53: 352 (1995).
Mycobank number: MB332021.
Type species: CBS 207.60; IFO 7833; MUCL 8014; UPSC 2185.
Saprobic on wood. Sexual morph: Undetermined. Asexual morph: Hyphomycetous. Hyphae hyaline to light brown, septate, branched. Conidiophores micronematous, mononematous, resemble non-specialised short lateral vegetative hyphae or conidiophore indistinct, conidia developing directly on hyphae. Conidia 14–24 μm × 7–11 μm, solitary, thick-walled, smooth, straight, 2–4-septate, strongly constricted at the septa, apical cell large subglobose, ellipsoidal, ovoidal to obpyriform, fuscous, reddish-brown to dark brown, basal cell smaller, obconical, light brown (Description based on Pang et al. 2011).
Material examined: USA, Tilia americana, wood, in seawater; Culture isolated by S.P. Meyers, No. F-123, Sept. 1960 (Holotype); USA Florida, Marine Laboratory, Univ. Miami.
Distribution: Aden, Australia, Bahamas, Bermuda, Cameron, Canada, France, Germany, India, Italy, Ivory Coast, Japan, Liberia, Norway, UK (England Scotland, Wales), USA (Alaska, Hawaii, Washington).
Molecular data: ITS GenBank MH857957 and LSU (MH869509) of CBS 207.60.
Notes: Taxonomically, the seven marine Trichocladium species were difficult to resolve until the advent of molecular phylogeny (Jones et al. 2009). Cultures of T. achrasporum were shown to be the asexual morph of Halosphaeriopsis mediosetigera (Shearer and Crane 1977). Based on sequence data Trichocladium constrictum has been designated the type species of a new genus Cucurbitinus as Cucurbitinus constrictus (Liu et al. 2020). Trichocladium medullare has been transferred to the new genus Kohlmeyeriopsis as K. medullaris (Klaubauf et al. 2014). Jones et al. (2019) listed four marine Trichocladium species T. alopallonellum T. griseum, T. lignicola, and T. melhae. Here we refer T. alopallonellum to Halosphaeriopsis based on sequence data of the type collection (Meyers and Moore 1960). The taxonomic status of the remaining marine Trichocladium species remains unresolved as does that of many other terrestrial and freshwater Trichocladium species (Index Fungorum, viewed April 2023).
In Figure 3, T. alopallonella groups in a sister clade with H. mediosetigera with high statistical support (100 %) in the Halosphaeriaceae, while other Trichocladium species form a sister clade to Humicola species. The phylogenetic tree was based on a combined analysis of the 28S and 18S rRNA sequence data. Morphologically, Trichocladium and Humicola species are difficult to separate resulting in confusing taxonomy, which is not surprising as they are phylogenetically closely related. Goh and Hyde (1999), in their review of the genus Trichocladium, illustrated the great diversity of conidial morphology of its species, with T. alopallonella often confused with Humicola as illustrated in the above synonymy.

Randomized axelerated maximum likelihood (RAxML) phylogenetic tree based on a combined analysis of the 28S and 18S rRNA sequence data. Bootstrap support values for ML (>70 %) are given above each branch. The newly transferred isolate is shown in blue. The tree is rooted to Ceratocystis adiposa CCFC212707.
2.5.6 Hypoxylon aurantium and H. mangrovei
Dayarathne et al. (2020a) introduced these two species based on morphological observations and nucleotide differences of the ITS and tub2 gene loci. Hypoxylon aurantium is a distinctive Hypoxylon species in possessing a bright orange ascostromatal surface which is somewhat similar to H. fendleri (Vasilyeva et al. 2007). Hypoxylon aurantium and H. fendleri formed a sister lineage with high statistical support (97 % ML, 0.97 PP) and the base pair difference of ITS and tub2 gene loci accounts 52 out of 604 (84 %) and 163 bp out of 1028 bp (15 %), respectively. Hypoxylon mangrovei strains formed a well-separated (100 % ML, 1.00 PP) lineage in a sister clade to H. lenormandii, with which it shares some morphological features. They differ in the size of stromata (2–3.5 × 1.5–2.8 × 0.4–1 mm vs. 13–85 × 6–27 × 0.5–1 mm) and base pair difference: ITS, 27 bp out of 470 bp (5.7 %) and tub2 73 out of 594 (12.3 %) and RPB2 data.
2.5.7 Hypoxylon teeravasati
Hypoxylon teeravasati was introduced by Devadatha et al. (in Phookamsak et al. 2019) on decaying wood of Avicennia marina, and decaying branches and twigs of Suaeda monoica from Tamil Nadu, India. Hypoxylon teeravasati groups with H. jaklitschii and H. lenormandii (terrestrial species) as a basal taxon, in a single monophyletic clade with high support. Hypoxylon teeravasati can be easily distinguished from H. lenormandii, H. jaklitschii, and H. croceum in having a burnt sienna surface, with KOH extractable pigments eye brown and larger perithecia (Phookamsak et al. 2019).
2.5.8 Halorosellinia xylocarpi
Halorosellinia is a phylogenetically well circumscribed genus with four of its five species forming a strongly supported clade in a phylogenetic analysis of 28S, ITS rDNA, tub2 and RPB2 genes (Dayarathne et al. 2020a). Halorosellinia xylocarpi is distinguishable from the other Halorosellina spp. by the unique opaque green colouration of the ascospores.
2.5.9 Monosporascus cannonballus (Figure 4)
Pollack and Uecker (1974) introduced this species from the roots of Cucumis melo (Cucurbitaceae) in Arizona. It causes root rot and vine decline (MRVD) of muskmelon in Greece (Markakis et al. 2018), and is reported as ubiquitous as a root endophyte (Robinson et al. 2020). The genomes of Monosporascus species were assembled by Robinson et al. (2020). The species was recorded for the first time from the marine environment by Jin and Huang (2009) from incubated drift and intertidal woods collected from the coasts of Yellow Sea and Bohai Sea (China). It groups in the Diatrypaceae (Xylariales, Xylariomycetidae, Sordariomcyetes) (Maharachchikumbura et al. 2015).

Monosporascus cannonballus. (A) Oil globules realised from squashed ascospore. (B) Ascus with single globose ascospore. (C) Thick-walled ascospore. (D) Ascus with three globose ascospores. (E, F) Released ascospores around the projecting neck of the ascoma. Scale bars: (A–D) = 20 μm, (E–F) = 300 μm.
2.5.10 Neocamarosporium aestuarinum
Gonçalves et al. (2019) described three new Neocamarosporium species from marine habitats, N. aestuarinum from seawater and N. halimiones and N. endophyticum from Halimione portulacoides based on a phylogenetic analysis of ITS rDNA sequences. Neocamarosporium aestuarinum grouped with N. jorjanensis with good support. Morphologically, both species are similar with slight differences in conidial dimension (Papizadeh et al. 2018), and 23 nucleotide positions (substitution or insertion/deletion) in ITS rDNA. It is important to calculate, however, the genetic differences within and between species of Neocamarosporium in order to use differences in nucleotide positions in differentiating species in the genus. Currently nine marine Neocamarosporium species are included in the marine fungi checklist, with new saltmarsh species N. aquaticum, N. chenopodii, N. maritimae N. phragmitis and N. salsolae (Calabon et al. 2021; Dayarathne et al. 2020a; Prematunga et al. 2023; Wanasinghe et al. 2017).
2.5.11 Neocosmospora rhizophorae
Dayarathne et al. (2020a) described this species from Rhizophora sp. collected in Thailand. Morphologically, this species is similar to Kallichroma tethys. Although Dayarathne et al. (2020a) distinguished Neocosmospora rhizophorae from K. tethys in having papillate ascomata, lacking a clypeoid ascomata, and an ascospore sheath, no ascospore sheath was actually reported for either K. tethys or K. glabra (Kohlmeyer and Volkmann-Kohlmeyer 1993). Also, a nucleotide BLAST search of 18S, 28S and ITS rDNA sequences available for N. rhizophorae resulted in predominantly Fusarium spp. The validity of this new species is doubtful.
2.5.12 Okeanomyces marinus
Okeanomyces marinus is an asexual fungus collected in Thailand (in Hyde et al. 2020a). This is the second species in the genus along with the type species O. cucullatus. The asexual stage of O. cucullatus is Periconia prolifica (Pang et al. 2004). Okeanomyces marinus is morphologically similar to P. prolifica and P. abyssa but differs from P. prolifica in lacking hyaline erect conidiophores and from P. abyssa in having smaller conidia (in Hyde et al. 2020a). In the same study, O. marinus formed a monophyletic group with isolates of O. cucullatus and P. prolifica based on a combined analysis of 18S and 28S rDNA.
2.5.13 Parathyridariella dematiacea
Poli et al. (2020b) introduced a monotypic genus Parathyridariella with Parathyridariella dematiacea as the type species isolated from the seagrass Posidonia oceanica in Italy. Multi-locus phylogenetic analysis shows that Parathyridariella dematiacea strains form a distinct clade within Thyridariaceae. Parathyridariella only developed solitary and chained chlamydospores in the mycelium above the wood surface of Pinus pinaster and Quercus ruber (cork).
2.5.14 Periconia salina
Six Periconia species were listed as marine in our last treatise of marine fungi: P. byssoides, P. cookei, P. digitata, P. echinochloae, P. minutissima, and P. salina (Jones et al. 2019). Periconia salina was introduced by Dayarathne et al. (2020a) on buried bark in sand dunes in Wales (UK).
2.5.15 Peroneutypa indica and P. polysporae
Both P. indica and P. polysporae formed a strongly supported group with other Peroneutypa spp. in a phylogenetic analysis of ITS rDNA and tub2 gene (Dayarathne et al. 2020a). Peroneutypa indica grouped with P. kochiana but, morphologically, P. indica has longer asci with a J-apical ring and a marine occurrence. Peroneutypa polysporae grouped with P. mangrovei but it is unique in the genus in having polysporous asci.
2.5.16 Phaeoseptum carolshearerianum
Phaeoseptum (type species P. aquaticum) was originally described from submerged wood in a freshwater swamp, and is characterized by black ascomata under a pseudoclypeus, narrowly cellular pseudoparaphyses, cylindrical-clavate asci with a short pedicel and brown ascospores, with 10–13(–15)-transversely septate and one vertical septum in each cell (Zhang et al. 2013). Using a combined LSU, SSU, TEF-1α, ITS, and RPB2 analysis, two isolates of Phaeoseptum carolshearerianum formed a sister clade with other Phaeoseptum species (P. aquaticum, P. mali, P. manglicola, P. terricola) (Dayarathne et al. 2020a). Morphologically, P. carolshearianum fits in the generic description of Phaeoseptum in having dark pigmented, immersed ascomata, cylindrical to clavate, pedicellate asci and muriform brown ascospores. However, P. carolshearianum lacks a clypeus and has asymmetric oblong to broadly fusiform ascospores. Currently, there are seven species in Phaeoseptum from terrestrial, freshwater, and marine habitats.
2.5.17 Qarounispora grandiappendiculata
Nourel-Din et al. (2022) introduced the monotypic genus Qarounispora to accommodate Q. grandiappendiculata that was identified from decaying submerged wood that was collected from Qaroun Lake, El-Faiyum governorate, Egypt. Qarounispora grandiappendiculata is characterized by having membranous ascomata that are yellow to orange in colour with one-layered peridium, semi-persistent asci and one-septate, thick-walled, distoseptate, hyaline to yellow-orange, broadly ellipsoidal ascospores with one polar appendage that is amorphous in structure, large in size and irregular in shape. Multigene analyses based on LSU, SSU and ITS rDNA placed Q. grandiappendiculata in the Halosphaeriaceae in a branch distinct from morphologically related fungal taxa with one polar appendage to the ascospores: Moana, Okeanomyces, Ophiodeira, and Tirispora. Qarounispora is phylogenetically related to Nimbospora, but Nimbospora has two types of appendages: an enlarged sheath surrounding the ascospores and fibrillar equatorial appendages (Koch 1982; Kohlmeyer and Volkmann-Kohlmeyer 1991).
2.5.18 Raghukumaria keshaphalae
Jones et al. (2020) introduced a monotypic genus Raghukumaria with Raghukumaria keshaphalae as the type species isolated from decayed wood of Aegiceras cornicualtum in India. Multi-locus phylogenetic analysis showed that Raghukumaria forms a distinct clade within the Trematosphaeriaceae and shares a close phylogenetic affinity to Halomassarina species. Raghukumaria differs from the latter in ascomatal (e.g., immersed, without a clypeus and periphyses, covered by hyphae-like setae) and ascospore morphology (e.g., broadly fusiform, with narrowly rounded ends, rough-walled or slightly verruculose, lacking globules, appendages or gelatinous sheaths) (Jones et al. 2020).
2.5.19 Remispora submersa
This new species was described based on the close phylogenetic relationship with Remispora quadri-remis and R. stellata in a phylogenetic tree of 28S and ITS rDNA (Gonçalves et al. 2021). This is an asexual fungus producing white mycelia and abundant cylindrical conidia. The conidiogenesis was unclear although it was described as “conidiophores reduced to conidiogenous cells”. This type of asexual morphology is unusual in the Halosphaeriaceae, as they generally have brown, septate conidia. A wider range of taxa should have been included in the phylogenetic analysis, especially the type species of Remispora, R. maritima. Holotype material was requested, especially slides of the conidiophores and conidia, but these were not available. A culture of R. submersa was provided, but failed to sporulate, despite growth on three different media and wood picks (Figure 5). The culture was re-sequenced, grouping in the Sarocladium clade and the species S. kiliense with high statistical support (Figure 6), a species with hyaline conidia as in those reported for R. submersa. Further studies are required to resolve this anomaly.
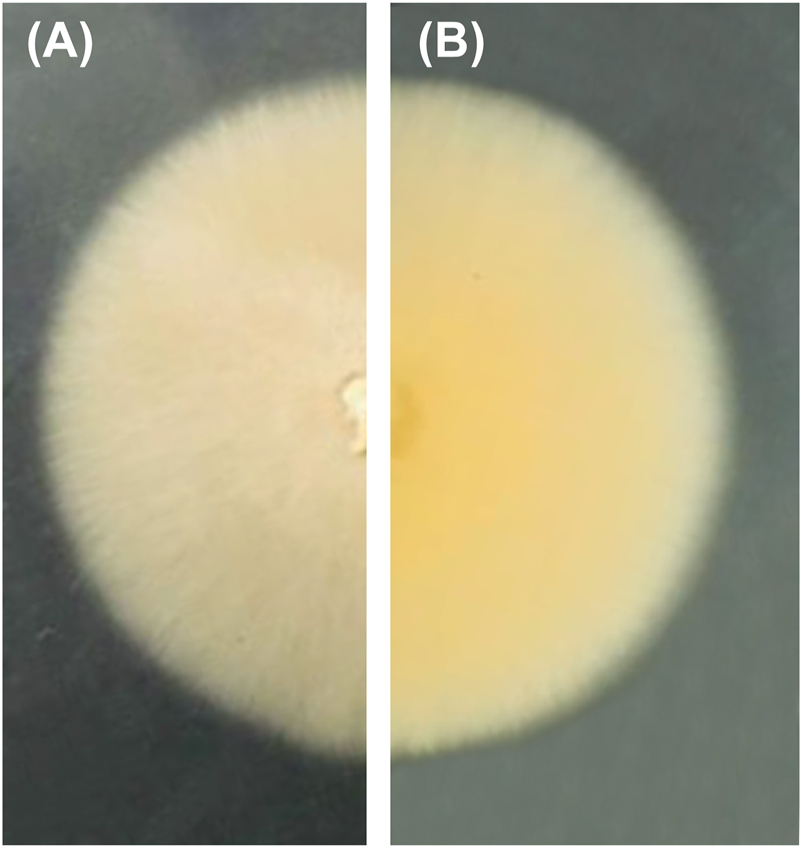
Colony of Remispora submersa (MUM 20.48) in malt extract agar (MEA) for 7 days at 25 °C: (A) obverse, (B) reverse.

Phylogram generated from maximum likelihood (ML) analysis based on combined LSU, ITS, and act1 sequence data representing Sarocladiaceae (Hypocreales). Thirty-seven strains are included in the combined analyses which comprised 1753 characters (504 characters for LSU, 478 characters for ITS, 771 characters for act1) after alignment. Acremonium variecolor strains CBS 130360 and FMR 11141 in Bionectriaceae (Hypocreales) were used as the outgroup taxa. The best scoring RAxML tree with a final likelihood value of −8526.836 is presented. The matrix had 435 distinct alignment patterns, 356 parsimony-informative, 73 singleton sites, and 1332 constant sites. Estimated base frequencies were as follows: A = 0.216, C = 0.298, G = 0.265, T = 0.221; substitution rates: AC = 2.09479, AG = 3.08268, AT = 2.09479, CG = 1.00000, CT = 10.05849, GT = 1.000000; gamma distribution shape parameter α = 0.574. Bootstrap support values for ML equal to or greater than 75 % are given above the nodes (left side). Bayesian posterior probabilities (BYPP) equal to or greater than 0.95 are given above the nodes (right side). Ex-type strains are in bold and newly generated sequences are in blue.
2.5.20 Rhytidhysteron bruguierae (Figure 7)
Rhytidhysteron bruguierae occurring on decaying Bruguiera wood, is phylogenetically related to the species R. mangrovei and R. thailandicum based on 18S, 28S rDNA and TEF-1α genes (Dayarathne et al. 2020a). Morphologically, R. bruguierae differs from the two species in having a red epithecium and thicker exciple (Kumar et al. 2019; Thambugala et al. 2016). Base pair differences of TEF-1α gene between R. bruguierae and R. mangrovei/R. thailandicum were over 3 % and were regarded as sufficient to warrant description of a new taxon (Jeewon and Hyde 2016). The type species of Rhytidhysteron is R. brasiliense which has no available sequences in GenBank (Spegazzini 1881). It is essential to obtain a culture of the type species to validate the taxonomic position of R. bruguierae and other species.
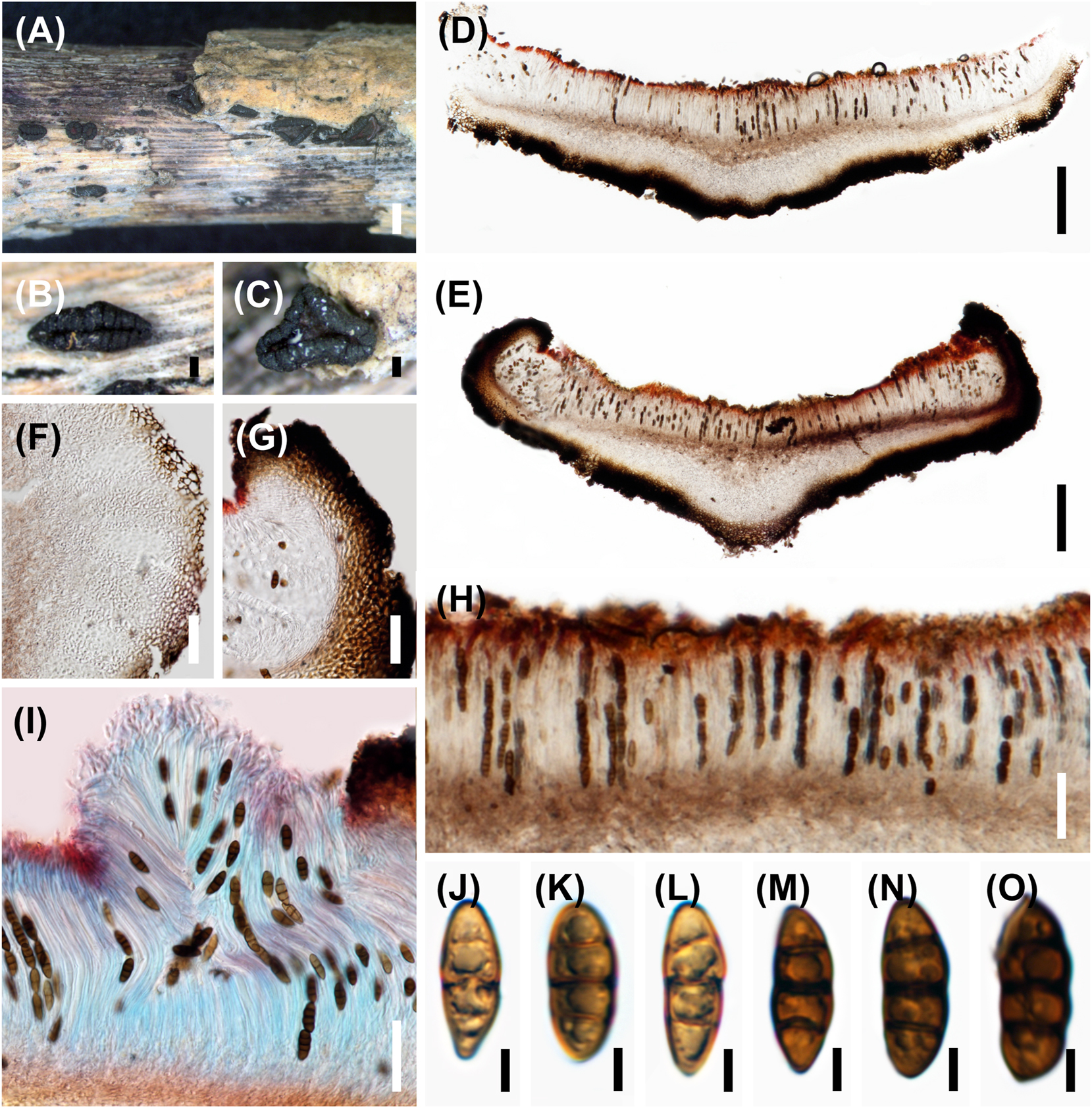
Rhytidhysteron bruguierae. (A–C) Appearance of hysterothecia on host. (D, E) Vertical section through hysteriothecium. (F, G) Cells of peridium. (H, I) Pseudoparaphyses, asci, and ascospores. (I) Stained with lactophenol cotton blue. (J–O) Ascospores. Scale bars: (A) = 1 mm, (B–E) = 200 μm, (F–I) = 50 μm, (J–O) = 5 μm.
2.5.21 Safagamyces marinus
Bakhit and Abdel-Wahab (2022) introduced the monotypic genus Safagamyces for the marine asexual species S. marinus that was recorded from decaying stem of Phragmites australis inside Safaga mangrove, Red Sea, Egypt. The taxon is characterized by straight or slightly curved, branched, smooth, 2–6 septate conidia and the sympodial conidial proliferation. Conidial cells increase in size and pigmentation from hyaline at the base to dark brown at the apex.
Phylogenetic analysis of a combined SSU and LSU rDNA sequences dataset placed S. marinus in the Halosphaeriaceae as a distinct basal branch to a node that contains the following marine genera: Antennospora, Cirrenalia, Cucullosporella, Cucurbitinus, Halosarpheia, Morakotiella, Pileomyces, Pseudolignincola and Remispora (Bakhit and Abdel-Wahab 2022). Safagamyces marinus is phylogenetically distant from the other two asexual genera: Cirrenalia and Cucurbitinus by having straight or slightly curved, branched conidia that are brown in colour with sympodial conidial proliferation. Conidia in Cirrenalia macrocephala are helicoid, reddish fuscous with determinate conidiogenesis (Meyers and Moore 1960; Zhao et al. 2007). Cucurbitinus species have straight or slightly curved conidia with constricted septa with determinate conidiogenesis. Both species of Cucurbitinus have larger conidial dimensions than S. marinus (Hernández-Restrepo et al. 2017; Kohlmeyer and Volkmann-Kohlmeyer 1991; Liu et al. 2020; Schmidt 1985).
2.5.22 Savoryella sarushimana (Figure 8)
Abdel-Wahab (2011) reported 27 marine taxa from driftwood collected at Sarushima Island, Japan, among which seven were suspected to be possibly new to science. Savoryella sarushimana is an asexual fungus forming a monophyletic group with Savoryella nypae in a combined analysis of 18S, 28S, ITS rDNA, RPB2, and TEF-1α (Zhang et al. 2019a). The conidia of S. sarushimana are characteristic in having 3–5-septa, not or slightly constricted at the septa, thick septa with dark-coloured bands, reddish brown to black mature conidia, with a rough surface and sometimes with long tapering projections forming hairy tufts. Savoryella sraushimana can be distinguished from S. nypae by its much larger conidia and wall ornamentations.
Savoryella sarushimana. (A–C) Holoblastic conidiogenous cells. (D–J) Variously shaped conidia. Note the proliferating conidia in (D, H). Scale bar = 10 μm.
2.6 Additional marine fungal taxa
Species not included in Jones et al. (2019) on the classification of marine fungi are listed in Supplementary Table 1. Higher taxonomic ranks are included for each species.
3 Conclusions
Since Jones et al. (2019), a total of 641 marine species have been discovered from various substrata, and recent updates in the classification of marine fungi. Most of these additions are novel taxa or new observations of extant species thriving in various marine substrata. The classification has been continuously updated with new data from molecular studies incorporating robust multi-locus phylogenetic analyses to large-scale whole genome sequencing techniques.
About the authors

Mark S. Calabon obtained a PhD (Biological Sciences) degree from the Center of Excellence in Fungal Research, Mae Fah Luang University (Thailand). He is an assistant professor in the University of the Philippines Visayas. His research focuses on the taxonomy, phylogeny, and classification of aquatic fungi.

Prof. E.B. Gareth Jones is a widely cited author on marine fungi, their ecology, physiology, and systematics and has co-edited several books and journal special issues. He has reported on marine fungi from around the world, in particular Asia, especially those found on mangroves. He supervised over 100 Ph.D. and a highly cited scientist.

Ka-Lai Pang obtained his BSc and PhD degrees from the City University of Hong Kong in 1998 and 2001, respectively. Prof. Pang studies the biology of marine fungi and fungus-like organisms and endophytic fungi associated with mangrove plants and macroalgae.

Mohamed A. Abdel-Wahab is a professor of marine mycology, and his research interest includes ecology and taxonomy of marine fungi, natural products from aquatic and endophytic fungi, and ecology and physiology of thraustochytrids.

Prof. Kevin D. Hyde is the director of the Center of Excellence in Fungal Research, Mae Fah Luang University. As a prolific researcher, internationally renowned mycologist, and most respected educator, he has been recognized as a highly cited scientist and distinguished mycologist by different organizations for his works in various disciplines of mycology. At present, he supervises 100+ graduate students, and is a proud mentor of his previous students, now collaborators, who became leading mycologists as educators and researchers.
-
Author contributions: The study conception was performed by Mark S. Calabon and E.B. Gareth Jones. All authors contributed to material preparation, data collection and analysis. All authors read, approved, and accepted responsibility for the entire content of this submitted manuscript and approved submission.
-
Research funding: This work was funded by the UP System Balik PhD Program (OVPAA-BPhD-2022-02). Ka-Lai Pang acknowledges the financial support of National Science and Technology Council (111-2621-B-019-001-MY3). Gareth Jones acknowledges research funding under the Distinguished Scientist Fellowship Program (DSFP), King Saud University, Kingdom of Saudi Arabia.
-
Conflict of interest statement: The authors declare that they have no conflicts of interest regarding this article.
-
Data availability: The datasets analysed during the current study are available from the corresponding author on reasonable request. All sequences generated here are available from GenBank with accession numbers: OR162608 (ITS).
References
Abdel-Wahab, M.A. (2011). Marine fungi from Sarushima Island, Japan, with a phylogenetic evaluation of the genus Naufragella. Mycotaxon 115: 443–456, https://doi.org/10.5248/115.443.Search in Google Scholar
Abdel-Wahab, M.A. and Bahkali, A.H.A. (2012). Taxonomy of filamentous anamorphic marine fungi: morphology and molecular evidence. In: Jones, E.B.G. and Pang, K.L. (Eds.), Marine fungi and fungal-like organisms. De Gruyter, Berlin, pp. 65–90.10.1515/9783110264067.65Search in Google Scholar
Abdel-Wahab, M.A., Hodhod, M.S., Bahkali, A.H.A., and Jones, E.B.G. (2014). Marine fungi of Saudi Arabia. Bot. Mar. 57: 323–335, https://doi.org/10.1515/bot-2014-0010.Search in Google Scholar
Abdel-Wahab, M.A., Bahkali, A.H., Elgorban, A.M., and Jones, E.B.G. (2021). High-throughput amplicon sequencing of fungi and microbial eukaryotes associated with the seagrass Halophila stipulacea (Forssk.) Asch. from Al-Leith mangroves, Saudi Arabia. Mycol. Prog. 20: 1365–1381, https://doi.org/10.1007/s11557-021-01744-2.Search in Google Scholar
Abdollahzadeh, J., Groenewald, J.Z., Coetzee, M.P.A., Wingfield, M.J., and Crous, P.W. (2020). Evolution of lifestyles in Capnodiales. Stud. Mycol. 95: 381–414, https://doi.org/10.1016/j.simyco.2020.02.004.Search in Google Scholar PubMed PubMed Central
Ahmed, S.A., van de Sande, W.W.J., Stevens, D.A., Fahal, A., van Diepeningen, A.D., Menken, S.B.J., and de Hoog, G.S. (2014). Revision of agents of black-grain eumycetoma in the order Pleosporales. Pers.: Mol. Phylogeny Evol. Fungi 33: 141–154, https://doi.org/10.3767/003158514x684744.Search in Google Scholar PubMed PubMed Central
Arzanlou, M., Groenewald, J.Z., Gams, W., Braun, U., Shin, H.D., and Crous, P.W. (2007). Phylogenetic and morphotaxonomic revision of Ramichloridium and allied genera. Stud. Mycol. 58: 57–93, https://doi.org/10.3114/sim.2007.58.03.Search in Google Scholar PubMed PubMed Central
Bakhit, M.S. and Abdel-Wahab, M.A. (2022). Safagamyces marinus gen. et sp. nov. (Halosphaeriaceae, Sordariomycetes) from Red Sea mangroves, Egypt. Phytotaxa 568: 221–229, https://doi.org/10.11646/phytotaxa.568.2.7.Search in Google Scholar
Barr, M.E. (1979). A classification of Loculoascomycetes. Mycologia 71: 935–957, https://doi.org/10.2307/3759283.Search in Google Scholar
Bojko, J. and Stentiford, G.D. (2022). Microsporidian pathogens of aquatic animals. In: Weiss, L.M. and Reinke, A.W. (Eds.), Microsporidia: Current advances in biology. Springer International Publishing, Cham, pp. 247–283.10.1007/978-3-030-93306-7_10Search in Google Scholar PubMed
Bonorden, H.F. (1864). Abhandlungen aus dem Gebiete der Mykologie. Abhandlungen der Naturforschenden Gesellschaft zu Halle 8: 1–168.Search in Google Scholar
Brown, J.C. (1958). Soil fungi of some British sand dunes in relation to soil type and succession. J. Ecol. 46: 641–664, https://doi.org/10.2307/2257543.Search in Google Scholar
Cain, R.F. (1961). Studies of soil fungi: iii. New species of Coniochaeta, Chaetomidium, and Thielavia. Canad. J. Bot. 39: 1231–1239, https://doi.org/10.1139/b61-107.Search in Google Scholar
Calabon, M.S., Jones, E.B.G., Promputtha, I., and Hyde, K.D. (2021). Fungal biodiversity in salt marsh ecosystems. J. Fungi 7: 648, https://doi.org/10.3390/jof7080648.Search in Google Scholar PubMed PubMed Central
Cheng, X.L., Li, W., and Zhang, T.Y. (2014). A new species of Phaeoisaria from intertidal marine sediment collected in Weihai, China. Mycotaxon 127: 17–24, https://doi.org/10.5248/127.17.Search in Google Scholar
Chomnunti, P., Bhat, D.J., Jones, E.B.G., Chukeatirote, E., Bahkali, A.H., and Hyde, K.D. (2012). Trichomeriaceae, a new sooty mould family of Chaetothyriales. Fungal Divers. 56: 63–76, https://doi.org/10.1007/s13225-012-0197-2.Search in Google Scholar
Chung, D., Kim, H., and Choi, H.S. (2019). Fungi in salterns. J. Microbiol. 57: 717–724, https://doi.org/10.1007/s12275-019-9195-3.Search in Google Scholar PubMed
Ciferri, R. (1958). Manuale di micologia medica. Renzo Cortina, Pavia, p. 370.Search in Google Scholar
Crous, P.W., Braun, U., and Groenewald, J.Z. (2007a). Mycosphaerella is polyphyletic. Stud. Mycol. 58: 1–32, https://doi.org/10.3114/sim.2007.58.01.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Schubert, K., Braun, U., De Hoog, G.S., Hocking, A.D., Shin, H.D., and Groenewald, J.Z. (2007b). Opportunistic, human-pathogenic species in the Herpotrichiellaceae are phenotypically similar to saprobic or phytopathogenic species in the Venturiaceae. Stud. Mycol. 58: 185–217, https://doi.org/10.3114/sim.2007.58.07.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Summerell, B.A., Carnegie, A.J., Wingfield, M.J., and Groenewald, J.Z. (2009a). Novel species of Mycosphaerellaceae and Teratosphaeriaceae. Pers.: Mol. Phylogeny Evol. Fungi 23: 119–146, https://doi.org/10.3767/003158509x479531.Search in Google Scholar
Crous, P.W., Summerell, B.A., Carnegie, A.J., Wingfield, M.J., Hunter, G.C., Burgess, T.I., Andjic, V., Barber, P.A., and Groenewald, J.Z. (2009b). Unravelling Mycosphaerella: Do you believe in genera? Pers.: Mol. Phylogeny Evol. Fungi 23: 99–118, https://doi.org/10.3767/003158509x479487.Search in Google Scholar
Crous, P.W., Summerell, B.A., Shivas, R.G., Romberg, M., Mel’nik, V.A., Verkley, G.J.M., and Groenewald, J.Z. (2011a). Fungal planet description sheets: 92-106. Pers.: Mol. Phylogeny Evol. Fungi 27: 130–162, https://doi.org/10.3767/003158511x617561.Search in Google Scholar
Crous, P.W., Summerell, B.A., Swart, L., Denman, S., Taylor, J.E., Bezuidenhout, C.M., Palm, M.E., Marincowitz, S., and Groenewald, J.Z. (2011b). Fungal pathogens of Proteaceae. Pers.: Mol. Phylogeny Evol. Fungi 27: 20–45, https://doi.org/10.3767/003158511x606239.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Tanaka, K., Summerell, B.A., and Groenewald, J.Z. (2011c). Additions to the Mycosphaerella complex. IMA Fungus 2: 49–64, https://doi.org/10.5598/imafungus.2011.02.01.08.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Wingfield, M.J., Richardson, D.M., Le Roux, J.J., Strasberg, D., Edwards, J., Roets, F., Hubka, V., Taylor, P.W.J., Heykoop, M., et al.. (2016). Fungal planet description sheets: 400–468. Pers.: Mol. Phylogeny Evol. Fungi 36: 316–458, https://doi.org/10.3767/003158516x692185.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Luangsa-Ard, J.J., Wingfield, M.J., Carnegie, A.J., Hernández-Restrepo, M., Lombard, L., Roux, J., Barreto, R.W., Baseia, I.G., Cano-Lira, J.F., et al.. (2018a). Fungal planet description sheets: 785–867. Pers.: Mol. Phylogeny Evol. Fungi 41: 239–251, https://doi.org/10.3767/persoonia.2018.41.12.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Wingfield, M.J., Burgess, T.I., Hardy, G.E.S.J., Gené, J., Guarro, J., Baseia, I.G., García, D., Gusmão, L.F.P., Souza-Motta, C.M., et al.. (2018b). Fungal planet description sheets: 716–784. Pers.: Mol. Phylogeny Evol. Fungi 40: 239–392, https://doi.org/10.3767/persoonia.2018.40.10.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Cowan, D.A., Maggs-Kölling, G., Yilmaz, N., Larsson, E., Angelini, C., Brandrud, T.E., Dearnaley, J.D.W., Dima, B., Dovana, F., et al.. (2020). Fungal planet description sheets: 1112–1181. Pers.: Mol. Phylogeny Evol. Fungi 45: 251–409, https://doi.org/10.3767/persoonia.2020.45.10.Search in Google Scholar PubMed PubMed Central
Crous, P.W., Hernández-Restrepo, M., Schumacher, R.K., Cowan, D.A., Maggs-Kölling, G., Marais, E., Wingfield, M.J., Yilmaz, N., Adan, O.C.G., Akulov, A., et al.. (2021). New and interesting Fungi. 4. Fungal Syst. Evol. 7: 255–343, https://doi.org/10.3114/fuse.2021.07.13.Search in Google Scholar PubMed PubMed Central
Dayarathne, M.C., Jones, E.B.G., Maharachchikumbura, S.S.N., Devadatha, B., Sarma, V.V., Khongphinitbunjong, K., Chomnunti, P., and Hyde, K.D. (2020a). Morpho-molecular characterization of microfungi associated with marine based habitats. Mycosphere 11: 1–188, https://doi.org/10.5943/mycosphere/11/1/1.Search in Google Scholar
Dayarathne, M.C., Wanasinghe, D.N., Devadatha, B., Abeywickrama, P., Gareth Jones, E.B., Chomnunti, P., Sarma, V.V., Hyde, K.D., Lumyong, S., and Mckenzie, E.H.C. (2020b). Modern taxonomic approaches to identifying diatrypaceous fungi from marine habitats, with a novel genus Halocryptovalsa Dayarathne et K.D.Hyde, gen. nov. Cryptogam. Mycol. 41: 21–67, https://doi.org/10.5252/cryptogamie-mycologie2020v41a3.Search in Google Scholar
de Gruyter, H., Woudenberg, J.H.C., Aveskamp, M.M., Verkley, G.J.M., Groenewald, J.Z., and Crous, P.W. (2013). Redisposition of phoma-like anamorphs in Pleosporales. Stud. Mycol. 75: 1–36, https://doi.org/10.3114/sim0004.Search in Google Scholar PubMed PubMed Central
Devadatha, B., Sarma, V.V., Jeewon, R., Wanasinghe, D.N., Hyde, K.D., and Gareth Jones, E.B. (2018). Thyridariella, a novel marine fungal genus from India: morphological characterization and phylogeny inferred from multigene DNA sequence analyses. Mycol. Prog. 17: 791–804, https://doi.org/10.1007/s11557-018-1387-4.Search in Google Scholar
Devadatha, B., Jones, E.B.G., Pang, K.L., Abdel-Wahab, M.A., Hyde, K.D., Sakayaroj, J., Bahkali, A.H., Calabon, M.S., Sarma, V.V., Sutreong, S., et al.. (2021). Occurrence and geographical distribution of mangrove fungi. Fungal Divers. 106: 137–227, https://doi.org/10.1007/s13225-020-00468-0.Search in Google Scholar
Diederich, P., Ertz, D., Lawrey, J.D., Sikaroodi, M., and Untereiner, W.A. (2013). Molecular data place the hyphomycetous lichenicolous genus Sclerococcum close to Dactylospora (Eurotiomycetes) and S. parmeliae in Cladophialophora (Chaetothyriales). Fungal Divers. 58: 61–72, https://doi.org/10.1007/s13225-012-0179-4.Search in Google Scholar
Diederich, P., Lawrey, J.D., and Ertz, D. (2018). The 2018 classification and checklist of lichenicolous fungi, with 2000 non-lichenized, obligately lichenicolous taxa. The Bryologist 121: 340–425, https://doi.org/10.1639/0007-2745-121.3.340.Search in Google Scholar
Ding, B., Yin, Y., Zhang, F., and Li, Z. (2011). Recovery and phylogenetic diversity of culturable fungi associated with marine sponges Clathrina luteoculcitella and Holoxea sp. in the South China Sea. Mar. Biotechnol. 13: 713–721, https://doi.org/10.1007/s10126-010-9333-8.Search in Google Scholar PubMed
Doweld, A. (2001). Prosyllabus tracheophytorum tentamen systematis plantarum vascularium (Tracheophyta). Geos, Moscow, pp. 200.Search in Google Scholar
Egidi, E., De Hoog, G.S., Isola, D., Onofri, S., Quaedvlieg, W., De Vries, M., Verkley, G.J.M., Stielow, J.B., Zucconi, L., and Selbmann, L. (2014). Phylogeny and taxonomy of meristematic rock-inhabiting black fungi in the Dothideomycetes based on multi-locus phylogenies. Fungal Divers. 65: 127–165, https://doi.org/10.1007/s13225-013-0277-y.Search in Google Scholar
Elíades, L.A., Voget, C.E., Arambarri, A.M., and Cabello, M.N. (2007). Fungal communities on decaying saltgrass (Distichlis spicata) in Buenos Aires province (Argentina). Sydowia 59: 227–234.Search in Google Scholar
Ertz, D. and Diederich, P. (2015). Dismantling Melaspileaceae: a first phylogenetic study of Buelliella, Hemigrapha, Karschia, Labrocarpon and Melaspilea. Fungal Divers. 71: 141–164, https://doi.org/10.1007/s13225-015-0321-1.Search in Google Scholar
Fell, J.W. and Hunter, I.L. (1979). Fungi associated with the decomposition of the Black Rush, Juncus roemerianus, in South Florida. Mycologia 71: 322–342, https://doi.org/10.2307/3759156.Search in Google Scholar
Fryar, S.C., Haelewaters, D., and Catcheside, D.E.A. (2019). Annabella australiensis gen. et sp. nov. (Helotiales, Cordieritidaceae) from South Australian mangroves. Mycol. Prog. 18: 973–981, https://doi.org/10.1007/s11557-019-01499-x.Search in Google Scholar
García, D., Stchigel, A.M., Cano, J., Calduch, M., Hawksworth, D.L., and Guarro, J. (2006). Molecular phylogeny of Coniochaetales. Mycol. Res. 110: 1271–1289, https://doi.org/10.1016/j.mycres.2006.07.007.Search in Google Scholar PubMed
Gessner, R.V. and Kohlmeyer, J. (1976). Geographical distribution and taxonomy of fungi from salt marsh Spartina. Canad. J. Bot. 54: 2023–2037, https://doi.org/10.1139/b76-216.Search in Google Scholar
Gnavi, G., Garzoli, L., Poli, A., Prigione, V., Burgaud, G., and Varese, G.C. (2017). The culturable mycobiota of Flabellia petiolata: first survey of marine fungi associated to a Mediterranean green alga. PLoS One 12: e0175941, https://doi.org/10.1371/journal.pone.0175941.Search in Google Scholar PubMed PubMed Central
Goh, T.K. and Hyde, K.D. (1999). A synopsis of Trichocladium species, based on the literature. Fungal Divers. 2: 101–118.Search in Google Scholar
Gonçalves, M.F.M., Aleixo, A., Vicente, T.F.L., Esteves, A.C., and Alves, A. (2019). Three new species of Neocamarosporium isolated from saline environments: N. aestuarinum sp. nov., N. endophyticum sp. nov. and N. halimiones sp. nov. Mycosphere 10: 608–621, https://doi.org/10.5943/mycosphere/10/1/11.Search in Google Scholar
Gonçalves, M.F.M., Vicente, T.F.L., Esteves, A.C., and Alves, A. (2020). Novel halotolerant species of Emericellopsis and Parasarocladium associated with macroalgae in an estuarine environment. Mycologia 112: 154–171, https://doi.org/10.1080/00275514.2019.1677448.Search in Google Scholar PubMed
Gonçalves, M.F.M., Abreu, A.C., Hilário, S., and Alves, A. (2021). Diversity of marine fungi associated with wood baits in the estuary Ria de Aveiro, with descriptions of Paralulworthia halima, comb. nov., Remispora submersa, sp. nov., and Zalerion pseudomaritima, sp. nov. Mycologia 113: 664–683, https://doi.org/10.1080/00275514.2021.1875710.Search in Google Scholar PubMed
Gunde-Cimerman, N., Zalar, P., De Hoog, S., and Plemenitaš, A. (2000). Hypersaline waters in salterns – natural ecological niches for halophilic black yeasts. FEMS Microbiol. Ecol. 32: 235–240, https://doi.org/10.1016/s0168-6496(00)00032-5.Search in Google Scholar
Hafellner, J. (1979). Karschia – revision einer Sammelgattung an der Grenze von lichenisierten und nicht lichenisierten Ascomyceten. Nova Hedwigia 62: 1–248.Search in Google Scholar
Hagestad, O.C., Hou, L., Andersen, J.H., Hansen, E.H., Altermark, B., Li, C., Kuhnert, E., Cox, R.J., Crous, P.W., Spatafora, J.W., et al.. (2021). Genomic characterization of three marine fungi, including Emericellopsis atlantica sp. nov. with signatures of a generalist lifestyle and marine biomass degradation. IMA Fungus 12: 1–23, https://doi.org/10.1186/s43008-021-00072-0.Search in Google Scholar PubMed PubMed Central
Hassett, B.T., Vonnahme, T.R., Peng, X., Jones, E.B.G., and Heuzé, C. (2020). Global diversity and geography of planktonic marine fungi. Bot. Mar. 63: 121–139, https://doi.org/10.1515/bot-2018-0113.Search in Google Scholar
Hawksworth, D.L. (1975). A revision of lichenicolous fungi accepted by Keissler in Coniothecium. Trans. Br. Mycol. Soc. 65: 219–238, https://doi.org/10.1016/s0007-1536(75)80005-2.Search in Google Scholar
Hawksworth, D.L. (1979). The lichenicolous hyphomycetes. Bull. Br. Mus. Nat. Hist. (Bot.) 6: 183–300.Search in Google Scholar
Hawksworth, D.L. (2000). Freshwater and marine lichen-forming fungi. In: Hyde, K.D., Ho, W.H. and Pointing, S.B. (Eds.), Aquatic mycology across the millennium. Fungal Diversity Press, Hong Kong, pp. 1–7.Search in Google Scholar
Hawksworth, D.L. and Boise, J.R. (1985). Some additional species of Astrosphaeriella, with a key to the members of the genus. Sydowia 38: 114–124.Search in Google Scholar
Hennebert, G.L. (1962). Wardomyces and Asteromyces. Canad. J. Bot. 40: 1203–1216, https://doi.org/10.1139/b62-111.Search in Google Scholar
Hernández-Restrepo, M., Gené, J., Castañeda-Ruiz, R.F., Mena-Portales, J., Crous, P.W., and Guarro, J. (2017). Phylogeny of saprobic microfungi from Southern Europe. Stud. Mycol. 86: 53–97, https://doi.org/10.1016/j.simyco.2017.05.002.Search in Google Scholar PubMed PubMed Central
Hibbett, D.S., Binder, M., Bischoff, J.F., Blackwell, M., Cannon, P.F., Eriksson, O.E., Huhndorf, S., James, T., Kirk, P.M., Lücking, R., et al.. (2007). A higher-level phylogenetic classification of the Fungi. Mycol. Res. 111: 509–547, https://doi.org/10.1016/j.mycres.2007.03.004.Search in Google Scholar PubMed
Hirayama, K. and Tanaka, K. (2011). Taxonomic revision of Lophiostoma and Lophiotrema based on reevaluation of morphological characters and molecular analyses. Mycoscience 52: 401–412, https://doi.org/10.1007/s10267-011-0126-3.Search in Google Scholar
Hongsanan, S., Maharachchikumbura, S.S.N., Hyde, K.D., Samarakoon, M.C., Jeewon, R., Zhao, Q., Al-Sadi, A.M., and Bahkali, A.H. (2017). An updated phylogeny of Sordariomycetes based on phylogenetic and molecular clock evidence. Fungal Divers. 84: 25–41, https://doi.org/10.1007/s13225-017-0384-2.Search in Google Scholar
Hongsanan, S., Hyde, K.D., Phookamsak, R., Wanasinghe, D.N., McKenzie, E.H.C., Sarma, V.V., Boonmee, S., Lücking, R., Bhat, D.J., Liu, N.G., et al.. (2020a). Refined families of Dothideomycetes: Dothideomycetidae and Pleosporomycetidae. Mycosphere 11: 1553–2107, https://doi.org/10.5943/mycosphere/11/1/13.Search in Google Scholar
Hongsanan, S., Hyde, K.D., Phookamsak, R., Wanasinghe, D.N., McKenzie, E.H.C., Sarma, V.V., Lücking, R., Boonmee, S., Bhat, J.D., Liu, N.G., et al.. (2020b). Refined families of Dothideomycetes: orders and families incertae sedis in Dothideomycetes. Fungal Divers. 105: 17–318, https://doi.org/10.1007/s13225-020-00462-6.Search in Google Scholar
Hoondee, P., Wattanagonniyom, T., Weeraphan, T., Tanasupawat, S., and Savarajara, A. (2019). Occurrence of oleaginous yeast from mangrove forest in Thailand. World J. Microbiol. Biotechnol. 35: 108, https://doi.org/10.1007/s11274-019-2680-3.Search in Google Scholar PubMed
Hutchison, L.J., Untereiner, W.A., and Hiratsuka, Y. (1995). Knufia cryptophialidica gen. et sp. nov., a dematiaceous hyphomycete isolated from black galls of trembling aspen (Populus tremuloides). Mycologia 87: 902–908, https://doi.org/10.2307/3760868.Search in Google Scholar
Hyde, K.D. (1988a). Studies on the tropical marine fungi of Brunei. Bot. J. Linn. Soc. 98: 135–151, https://doi.org/10.1111/j.1095-8339.1988.tb01700.x.Search in Google Scholar
Hyde, K.D. (1988b). The genus Linocarpon from the mangrove plam Nypa fruticans. Trans. Mycol. Soc. Japan 29: 339–350.Search in Google Scholar
Hyde, K.D. (1989). Ecology of tropical marine fungi. Hydrobiologia 178: 199–208, https://doi.org/10.1007/bf00006027.Search in Google Scholar
Hyde, K.D. (1992). Fungi from decaying intertidal fronds of Nypa fruticans, including three new genera and four new species. Bot. J. Linn. Soc. 110: 95–110, https://doi.org/10.1111/j.1095-8339.1992.tb00284.x.Search in Google Scholar
Hyde, K.D. (2022). The numbers of fungi. Fungal Divers. 114: 1–1.10.1007/s13225-022-00507-ySearch in Google Scholar
Hyde, K.D. and Alias, S.A. (2000). Biodiversity and distribution of fungi associated with decomposing Nypa fruticans. Biodivers. Conserv. 9: 393–402, https://doi.org/10.1023/a:1008911121774.10.1023/A:1008911121774Search in Google Scholar
Hyde, K.D. and Borse, B.D. (1986). Marine fungi from Seychelles V. Biatriospora marina gen. et sp. nov. from mangrove wood. Mycotaxon 26: 263–270.Search in Google Scholar
Hyde, K.D. and Rappaz, F. (1993). Eutypa bathurstensis sp. nov. from intertidal Avicennia. Mycol. Res. 97: 861–864, https://doi.org/10.1016/s0953-7562(09)81163-5.Search in Google Scholar
Hyde, K.D., Jones, E.B.G., Liu, J.K., Ariyawansa, H., Boehm, E., Boonmee, S., Braun, U., Chomnunti, P., Crous, P.W., Dai, D.Q., et al.. (2013). Families of Dothideomycetes. Fungal Divers. 63: 1–313, https://doi.org/10.1007/s13225-013-0263-4.Search in Google Scholar
Hyde, K.D., Maharachchikumbura, S.S.N., Hongsanan, S., Samarakoon, M.C., Lücking, R., Pem, D., Harishchandra, D., Jeewon, R., Zhao, R.L., Xu, J.C., et al.. (2017). The ranking of fungi: a tribute to David L. Hawksworth on his 70th birthday. Fungal Divers. 84: 1–23, https://doi.org/10.1007/s13225-017-0383-3.Search in Google Scholar
Hyde, K.D., Chaiwan, N., Norphanphoun, C., Boonmee, S., Camporesi, E., Chethana, K.W.T., Dayarathne, M.C., de Silva, N.I., Dissanayake, A.J., Ekanayaka, A.H., et al.. (2018). Mycosphere notes 169–224. Mycosphere 9: 271–430, https://doi.org/10.5943/mycosphere/9/2/8.Search in Google Scholar
Hyde, K.D., Jeewon, R., Chen, Y.J., Bhunjun, C.S., Calabon, M.S., Jiang, H.B., Lin, C.G., Norphanphoun, C., Sysouphanthong, P., Pem, D., et al.. (2020a). The numbers of fungi: is the descriptive curve flattening? Fungal Divers. 103: 219–271, https://doi.org/10.1007/s13225-020-00458-2.Search in Google Scholar
Hyde, K.D., Norphanphoun, C., Maharachchikumbura, S.S.N., Bhat, D.J., Jones, E.B.G., Bundhun, D., Chen, Y.J., Bao, D.F., Boonmee, S., Calabon, M.S., et al.. (2020b). Refined families of Sordariomycetes. Mycosphere 11: 305–1059, https://doi.org/10.5943/mycosphere/11/1/7.Search in Google Scholar
Jaklitsch, W.M. and Voglmayr, H. (2016). Hidden diversity in Thyridaria and a new circumscription of the Thyridariaceae. Stud. Mycol. 85: 35–64, https://doi.org/10.1016/j.simyco.2016.09.002.Search in Google Scholar PubMed PubMed Central
Jeewon, R. and Hyde, K.D. (2016). Establishing species boundaries and new taxa among fungi: Recommendations to resolve taxonomic ambiguities. Mycosphere 7: 1669–1677, https://doi.org/10.5943/mycosphere/7/11/4.Search in Google Scholar
Jin, J. and Huang, J.G. (2009). The finding of Monosporascus cannonballus in marine environment (in Chinese). Mycosystema 28: 865–869.Search in Google Scholar
Jin, J., Li, G.F., Li, B.H., Zhang, T.Y., Liang, C., and Li, B.D. (2004). Wood-inhabiting marine fungi from the coast of Shandong I. Mycosystema 23: 211–215.Search in Google Scholar
Jin, J., Li, G.F., Zhang, T.Y., and Li, B.D. (2005). New report of marine higher fungi from the Yellow Sea, China I. Mycosystema 24: 336–339.Search in Google Scholar
Jones, E.B.G. (1963). Marine fungi: II. Ascomycetes and deuteromycetes from submerged wood and drift Spartina. Trans. Br. Mycol. Soc. 46: 135–144, https://doi.org/10.1016/s0007-1536(63)80015-7.Search in Google Scholar
Jones, E.B.G. (1968). The distribution of marine fungi on wood submerged in the sea. In: Walters, H. and Elphick, J. (Eds.), Biodeterioration of materials. Elsevier, London, pp. 460–485.Search in Google Scholar
Jones, E.B.G. and Jennings, D.H. (1965). The effect of cations on the growth of fungi. New Phytol. 64: 86–100, https://doi.org/10.1111/j.1469-8137.1965.tb05378.x.Search in Google Scholar
Jones, E.B.G. and Ward, A.W. (1973). Septate conidia in Asteromyces cruciatus. Trans. Br. Mycol. Soc. 61: 181–186, https://doi.org/10.1016/s0007-1536(73)80100-7.Search in Google Scholar
Jones, E.B.G., Abdel-Wahab, M.A., Alias, S.A., and Hsieh, S.Y. (1999). Dactylospora mangrovei sp. nov. (Discomycetes, Ascomycota) from mangrove wood. Mycoscience 40: 317–320, https://doi.org/10.1007/bf02463875.Search in Google Scholar
Jones, E.B.G., Sakayaroj, J., Suetrong, S., Somrithipol, S., and Pang, K.L. (2009). Classification of marine Ascomycota, anamorphic taxa and Basidiomycota. Fungal Divers. 35: 1–187.Search in Google Scholar
Jones, E.B.G., Suetrong, S., Sakayaroj, J., Bahkali, A.H., Abdel-Wahab, M.A., Boekhout, T., and Pang, K.L. (2015). Classification of marine Ascomycota, Basidiomycota, Blastocladiomycota and Chytridiomycota. Fungal Divers. 73: 1–72, https://doi.org/10.1007/s13225-015-0339-4.Search in Google Scholar
Jones, E.B.G., Pang, K.L., Abdel-Wahab, M.A., Scholz, B., Hyde, K.D., Boekhout, T., Ebel, R., Rateb, M.E., Henderson, L., Sakayaroj, J., et al.. (2019). An online resource for marine fungi. Fungal Divers. 96: 347–433, https://doi.org/10.1007/s13225-019-00426-5.Search in Google Scholar
Joshi, Y. (2021). Two new species of lichenicolous fungus Sclerococcum (Dactylosporaceae, Sclerococcales) from India. Acta Bot. Hung. 63: 67–75.10.1556/034.63.2021.1-2.5Search in Google Scholar
Jones, E.B.G., Devadatha, B., Abdel-Wahab, M.A., Dayarathne, M.C., Zhang, S.N., Hyde, K.D., Liu, J.K., Bahkali, A.H., Sarma, V.V., Tibell, S., et al.. (2020). Phylogeny of new marine Dothideomycetes and Sordariomycetes from mangroves and deep-sea sediments. Bot. Mar. 63: 155–181, https://doi.org/10.1515/bot-2019-0014.Search in Google Scholar
Jülich, W. (1982). Higher taxa of basidiomycetes. Bibl. Mycol. 85: 1–485.Search in Google Scholar
Kirk, P., Cannon, P., Minter, D., and Stalpers, J. (2008). Dictionary of the fungi. CABI, Trowbridge, p. 784.Search in Google Scholar
Klaubauf, S., Tharreau, D., Fournier, E., Groenewald, J.Z., Crous, P.W., De Vries, R.P., and Lebrun, M.H. (2014). Resolving the polyphyletic nature of Pyricularia (Pyriculariaceae). Stud. Mycol. 79: 85–120, https://doi.org/10.1016/j.simyco.2014.09.004.Search in Google Scholar PubMed PubMed Central
Koch, J. (1982). Some lignicolous marine fungi from Sri Lanka. Nord. J. Bot. 2: 163–169, https://doi.org/10.1111/j.1756-1051.1982.tb01177.x.Search in Google Scholar
Kohlmeyer, J. (1967). Intertidal and phycophilous fungi from Tenerife (Canary Islands). Trans. Br. Mycol. Soc. 50: 137–147, https://doi.org/10.1016/s0007-1536(67)80070-6.Search in Google Scholar
Kohlmeyer, J. and Kohlmeyer, E. (1964–1969). Icones fungorum maris. Fasc. 1–9. J. Cramer, Lehre, pp. 1–80.Search in Google Scholar
Kohlmeyer, J. and Kohlmeyer, E. (1965). New marine fungi from mangroves and trees along eroding shorelines. Nova Hedwigia 9: 89–104.Search in Google Scholar
Kohlmeyer, J. and Volkmann-Kohlmeyer, B. (1991). Illustrated key to the filamentous higher marine fungi. Bot. Mar. 34: 1–61, https://doi.org/10.1515/botm.1991.34.1.1.Search in Google Scholar
Kohlmeyer, J. and Volkmann-Kohlmeyer, B. (1993). Observations on Hydronectria and Kallichroma gen. nov. Mycol. Res. 97: 753–761, https://doi.org/10.1016/s0953-7562(09)80157-3.Search in Google Scholar
Kohlmeyer, J. and Volkmann-Kohlmeyer, B. (1998). Dactylospora canariensis sp. nov. and notes on D. haliotrepha. Mycotaxon 67: 247–250.Search in Google Scholar
Kohlmeyer, J., Volkmann-Kohlmeyer, B., and Eriksson, O.E. (1996). Fungi on Juncus roemerianus. 8. New bitunicate ascomycetes. Canad. J. Bot. 74: 1830–1840, https://doi.org/10.1139/b96-220.Search in Google Scholar
Kolařík, M. (2018). New taxonomic combinations in endophytic representatives of the genus Nigrograna. Czech Mycol. 70: 123–126, https://doi.org/10.33585/cmy.70202.Search in Google Scholar
Kolařík, M., Spakowicz, D.J., Gazis, R., Shaw, J., Kubátová, A., Nováková, A., Chudíčková, M., Forcina, G.C., Kang, K.W., Kelnarová, I., et al.. (2017). Biatriospora (Ascomycota: Pleosporales) is an ecologically diverse genus including facultative marine fungi and endophytes with biotechnological potential. Plant Syst. Evol. 303: 35–50, https://doi.org/10.1007/s00606-016-1350-2.Search in Google Scholar
Konta, S., Hongsanan, S., Liu, J.K., Eungwanichayapant, P.D., Jeewon, R., Hyde, K.D., Maharachchikumbura, S.S.N., and Boonmee, S. (2017). Leptosporella (Leptosporellaceae fam. nov.) and Linocarpon and Neolinocarpon (Linocarpaceae fam. nov.) are accommodated in Chaetosphaeriales. Mycosphere 8: 1943–1974, https://doi.org/10.5943/mycosphere/8/10/16.Search in Google Scholar
Kumar, V., Cheewangkoon, R., Thambugala, K.M., Jones, E.B.G., Brahmanage, R.S., Doilom, M., Jeewon, R., and Hyde, K.D. (2019). Rhytidhysteron mangrovei (Hysteriaceae), a new species from mangroves in Phetchaburi Province, Thailand. Phytotaxa 401: 166–178, https://doi.org/10.11646/phytotaxa.401.3.2.Search in Google Scholar
Kunthiphun, S., Wattanagonniyom, T., Endoh, R., Takashima, M., Ohkuma, M., Tanasupawat, S., and Savarajara, A. (2019). Heterocephalacria mucosa sp. nov., a new basidiomycetous yeast species isolated from a mangrove forest in Thailand. Int. J. Syst. Evol. Microbiol. 69: 2823–2827, https://doi.org/10.1099/ijsem.0.003562.Search in Google Scholar PubMed
Le Calvez, T., Burgaud, G., Mahé, S., Barbier, G., and Vandenkoornhuyse, P. (2009). Fungal diversity in deep-sea hydrothermal ecosystems. Appl. Environ. Microbiol. 75: 6415–6421, https://doi.org/10.1128/aem.00653-09.Search in Google Scholar PubMed PubMed Central
Li, J.F., Phookamsak, R., Jeewon, R., Tibpromma, S., Maharachchikumbura, S.S.N., Bhat, D.J., Chukeatirote, E., Lumyong, S., Hyde, K.D., and McKenzie, E.H.C. (2017). Establishment of Zygosporiaceae fam. nov. (Xylariales, Sordariomycetes) based on rDNA sequence data to accommodate Zygosporium. Mycosphere 8: 1855–1868, https://doi.org/10.5943/mycosphere/8/10/10.Search in Google Scholar
Liu, L.L., Liu, N., Yang, J., Chen, Y., and Liu, Z. (2020). Cucurbitinus gen. nov. (Halosphaeriaceae, Microascales), a new genus to accommodate Cucurbitinus constrictus comb. nov. and Cucurbitinus ibericus comb. nov. Phytotaxa 455: 119–136, https://doi.org/10.11646/phytotaxa.455.2.4.Search in Google Scholar
Liu, X.Z., Wang, Q.M., Göker, M., Groenewald, M., Kachalkin, A.V., Lumbsch, H.T., Millanes, A.M., Wedin, M., Yurkov, A.M., Boekhout, T., et al.. (2015). Towards an integrated phylogenetic classification of the Tremellomycetes. Stud. Mycol. 81: 85–147, https://doi.org/10.1016/j.simyco.2015.12.001.Search in Google Scholar PubMed PubMed Central
Locquin, M. (1984). Mycologie générale et structurale. Masson, Paris, pp. 551.Search in Google Scholar
Lücking, R., Lumbsch, H.T., Di Stéfano, J.F., Lizano, D., Carranza, J., Bernecker, A., Chaves, J.L., and Umaña, L. (2008). Eremithallus costaricensis (Ascomycota: Lichinomycetes: Eremothallales), a new fungal lineage with a novel lichen symbiotic lifestyle discovered in an urban relict forest in Costa Rica. Symbiosis 46: 161–170.10.3158/0015-0746(2008)46[1:AFAOTT]2.0.CO;2Search in Google Scholar
Luttrell, E.S. (1951). Taxonomy of the Pyrenomycetes. Univ. Mo. Stud., Sci. Ser. 24: 1–120.Search in Google Scholar
Maharachchikumbura, S.S.N., Hyde, K.D., Jones, E.B.G., McKenzie, E.H.C., Huang, S.K., Abdel-Wahab, M.A., Daranagama, D.A., Dayarathne, M., D’souza, M.J., Goonasekara, I.D., et al.. (2015). Towards a natural classification and backbone tree for Sordariomycetes. Fungal Divers. 72: 199–301, https://doi.org/10.1007/s13225-015-0331-z.Search in Google Scholar
Mantle, P.G., Hawksworth, D.L., Pazoutova, S., Collinson, L.M., and Rassing, B.R. (2006). Amorosia littoralis gen. sp. nov., a new genus and species name for the scorpinone and caffeine-producing hyphomycete from the littoral zone in The Bahamas. Mycol. Res. 110: 1371–1378, https://doi.org/10.1016/j.mycres.2006.09.013.Search in Google Scholar
Mapook, A., Hyde, K.D., Dai, D.Q., Li, J., Jones, E.B.G., Bahkali, A.H., and Boonmee, S. (2016). Muyocopronales, ord. nov., (Dothideomycetes, Ascomycota) and a reappraisal of Muyocopron species from northern Thailand. Phytotaxa 265: 225–237, https://doi.org/10.11646/phytotaxa.265.3.3.Search in Google Scholar
Mapook, A., Hyde, K.D., McKenzie, E.H.C., Jones, E.B.G., Bhat, D.J., Jeewon, R., Stadler, M., Samarakoon, M.C., Malaithong, M., Tanunchai, B., et al.. (2020). Taxonomic and phylogenetic contributions to fungi associated with the invasive weed Chromolaena odorata (Siam weed). Fungal Divers. 101: 1–175, https://doi.org/10.1007/s13225-020-00444-8.Search in Google Scholar
Maria, G.L. and Sridhar, K.R. (2003). Diversity of filamentous fungi on woody litter of five mangrove plant species from the southwest coast of India. Fungal Divers. 14: 109–126.Search in Google Scholar
Markakis, E.A., Trantas, E.A., Lagogianni, C.S., Mpalantinaki, E., Pagoulatou, M., Ververidis, F., and Goumas, D.E. (2018). First report of root rot and vine decline of melon caused by Monosporascus cannonballus in Greece. Plant Dis. 102: 1036, https://doi.org/10.1094/pdis-10-17-1568-pdn.Search in Google Scholar
Meyers, S.P. and Moore, R.T. (1960). Thalassiomycetes II new genera and species of deuteromycetes. Am. J. Bot. 47: 345–349, https://doi.org/10.1002/j.1537-2197.1960.tb07134.x.Search in Google Scholar
Montagne, J.P.F.C. (1842). Troisième Centurie de plantes cellulaires exotiques nouvelles, Décades I, II, III et IV. Fungi cubenses. Ann. Sci. Nat., Bot. 17: 119–128.Search in Google Scholar
Moreau, F. and Moreau, V. (1941). Première contribution à l’étude de la microflore des dunes. Revue de Mycologie (Paris) 6: 49–94.Search in Google Scholar
Nolan, R.A. (1972). Asteromyces cruciatus from North America. Mycologia 64: 430–433, https://doi.org/10.2307/3757849.Search in Google Scholar
Nourel-Din, A.A.H., Abdel-Aziz, F.A., and Abdel-Wahab, M.A. (2022). Qarounispora grandiappendiculata gen. et sp. nov. (Halosphaeriaceae, Microascales) from Qaroun Lake, Egypt. Phytotaxa 530: 86–94, https://doi.org/10.11646/phytotaxa.530.1.7.Search in Google Scholar
Olariaga, I., Teres, J., Martín, J., Prieto, M., and Baral, H.O. (2019). Pseudosclerococcum golindoi gen. et sp. nov., a new taxon with apothecial ascomata and a chalara-like anamorph within the Sclerococcales (Eurotiomycetes). Mycol. Prog. 18: 895–905, https://doi.org/10.1007/s11557-019-01500-7.Search in Google Scholar
Pang, K.L., Jheng, J.S., and Jones, E.B.G. (2011). Marine mangrove fungi of Taiwan. National Taiwan Ocean University, Taiwan.Search in Google Scholar
Pang, K.L., Jones, E.B.G., Vrijmoed, L.L.P., and Vikineswary, S. (2004). Okeanomyces, a new genus to accommodate Halosphaeria cucullata (Halosphaeriales, Ascomycota). Bot. J. Linn. Soc. 146: 223–229, https://doi.org/10.1111/j.1095-8339.2004.00314.x.Search in Google Scholar
Pang, K.L., Guo, S.Y., Alias, S.A., Hafellner, J., and Jones, E.B.G. (2014). A new species of marine Dactylospora and its phylogenetic affinities within the Eurotiomycetes, Ascomycota. Bot. Mar. 57: 315–321, https://doi.org/10.1515/bot-2014-0025.Search in Google Scholar
Pang, K.L., Hassett, B.T., Shaumi, A., Guo, S.Y., Sakayaroj, J., Chiang, M.W.L., Yang, C.H., and Jones, E.B.G. (2021). Pathogenic fungi of marine animals: a taxonomic perspective. Fungal Biol. Rev. 38: 92–106, https://doi.org/10.1016/j.fbr.2021.03.008.Search in Google Scholar
Panno, L., Bruno, M., Voyron, S., Anastasi, A., Gnavi, G., Miserere, L., and Varese, G.C. (2013). Diversity, ecological role and potential biotechnological applications of marine fungi associated to the seagrass Posidonia oceanica. New Biotechnol. 30: 685–694, https://doi.org/10.1016/j.nbt.2013.01.010.Search in Google Scholar PubMed
Papizadeh, M., Wijayawardene, N.N., Amoozegar, M.A., Saba, F., Fazeli, S.A.S., and Hyde, K.D. (2018). Neocamarosporium jorjanensis, N. persepolisi, and N. solicola spp. nov. (Neocamarosporiaceae, Pleosporales) isolated from saline lakes of Iran indicate the possible halotolerant nature for the genus. Mycol. Prog. 17: 661–679, https://doi.org/10.1007/s11557-017-1341-x.Search in Google Scholar
Phookamsak, R., Norphanphoun, C., Tanaka, K., Dai, D.Q., Luo, Z.L., Liu, J.K., Su, H.Y., Bhat, D.J., Bahkali, A.H., Mortimer, P.E., et al.. (2015). Towards a natural classification of Astrosphaeriella-like species; introducing Astrosphaeriellaceae and Pseudoastrosphaeriellaceae fam. nov. and Astrosphaeriellopsis, gen. nov. Fungal Divers. 74: 143–197, https://doi.org/10.1007/s13225-015-0352-7.Search in Google Scholar
Phookamsak, R., Hyde, K.D., Jeewon, R., Bhat, D.J., Jones, E.B.G., Maharachchikumbura, S.S.N., Raspé, O., Karunarathna, S.C., Wanasinghe, D.N., Hongsanan, S., et al.. (2019). Fungal diversity notes 929–1035: taxonomic and phylogenetic contributions on genera and species of fungi. Fungal Divers. 95: 1–273, https://doi.org/10.1007/s13225-019-00421-w.Search in Google Scholar
Phukhamsakda, C., Nilsson, R.H., Bhunjun, C.S., de Farias, A.R.G., Sun, Y.R., Wijesinghe, S.N., Raza, M., Bao, D.F., Lu, L., Tibpromma, S., et al.. (2022). The numbers of fungi: contributions from traditional taxonomic studies and challenges of metabarcoding. Fungal Divers 5: 1–60.10.1007/s13225-022-00502-3Search in Google Scholar
Pino-Bodas, R., Zhurbenko, M.P., and Stenroos, S. (2017). Phylogenetic placement within Lecanoromycetes of lichenicolous fungi associated with Cladonia and some other genera. Pers.: Mol. Phylogeny Evol. Fungi 39: 91–117, https://doi.org/10.3767/persoonia.2017.39.05.Search in Google Scholar PubMed PubMed Central
Poli, A., Bovio, E., Ranieri, L., Varese, G.C., and Prigione, V. (2020a). Fungal diversity in the Neptune Forest: comparison of the mycobiota of Posidonia oceanica, Flabellia petiolata, and Padina pavonica. Front. Microbiol. 11: 933, https://doi.org/10.3389/fmicb.2020.00933.Search in Google Scholar PubMed PubMed Central
Poli, A., Bovio, E., Ranieri, L., Varese, G.C., and Prigione, V. (2020b). News from the sea: a new genus and seven new species in the pleosporalean families Roussoellaceae and Thyridariaceae. Diversity 12: 144, https://doi.org/10.3390/d12040144.Search in Google Scholar
Pollack, F.G. and Uecker, F.A. (1974). Monosporascus cannonballus an unusual ascomycete in cantaloupe roots. Mycologia 66: 346–349, https://doi.org/10.2307/3758370.Search in Google Scholar
Poncet, R., Voglmayr, H., and Kervran, L. (2022). A new species of Stigmatodiscus (Ascomycota, Dothideomycetes, Stigmatodiscaceae) from Juan de Nova (Mozambique Channel, Scattered Islands, French Southern and Antarctic Lands). Plant Fungal Syst. 67: 55–62, https://doi.org/10.35535/pfsyst-2022-0007.Search in Google Scholar
Prematunga, C., Boonmee, S., Jones, E.B.G., and Calabon, M.S. (2023). Neocamarosporium aquaticum (Neocamarosporiaceae, Dothideomycetes), a novel fungus from salt marsh habitats. Bot. Mar. 66: 271–279. https://doi.org/10.1515/bot-2022-0071.Search in Google Scholar
Quaedvlieg, W., Binder, M., Groenewald, J.Z., Summerell, B.A., Carnegie, A.J., Burgess, T.I., and Crous, P.W. (2014). Introducing the consolidated species concept to resolve species in the Teratosphaeriaceae. Pers.: Mol. Phylogeny Evol. Fungi 33: 1–40, https://doi.org/10.3767/003158514x681981.Search in Google Scholar PubMed PubMed Central
Raitviir, A. (2004). Revised synopsis of the Hyaloscyphaceae. Scripta Mycol. Tartu 20: 1–133.Search in Google Scholar
Réblová, M., Untereiner, W.A., and Réblová, K. (2013). Novel evolutionary lineages revealed in the Chaetothyriales (Fungi) based on multigene phylogenetic analyses and comparison of its secondary structure. PLoS One 8: e63547, https://doi.org/10.1371/journal.pone.0063547.Search in Google Scholar PubMed PubMed Central
Réblová, M., Seifert, K.A., Fournier, J., and Štepánek, V. (2016). Newly recognised lineages of perithecial ascomycetes: the new orders Conioscyphales and Pleurotheciales. Pers.: Mol. Phylogeny Evol. Fungi 37: 57–81, https://doi.org/10.3767/003158516x689819.Search in Google Scholar
Réblová, M., Untereiner, W.A., Štěpánek, V., and Gams, W. (2017). Disentangling Phialophora section Catenulatae: disposition of taxa with pigmented conidiophores and recognition of a new subclass, Sclerococcomycetidae (Eurotiomycetes). Mycol. Prog. 16: 27–46, https://doi.org/10.1007/s11557-016-1248-y.Search in Google Scholar
Ren, X., Chen, C., Ye, Y., Xu, Z., Zhao, Q., Luo, X., Liu, Y., and Guo, P. (2022). Anti-inflammatory compounds from the mangrove endophytic fungus Amorosia sp. SCSIO 41026. Front. Microbiol. 13: 976399, https://doi.org/10.3389/fmicb.2022.976399.Search in Google Scholar PubMed PubMed Central
Robinson, A.J., Natvig, D.O., and Chain, P.S.G. (2020). Genomic analysis of diverse members of the fungal genus monosporascus reveals novel lineages, unique genome content and a potential bacterial associate. G3: Genes, Genomes, Genetics 10: 2573–2583, https://doi.org/10.1534/g3.120.401489.Search in Google Scholar PubMed PubMed Central
Saito, K. (1922). Untersuchungen über die atmosphärischen Pilzkeime. III Mitteilung. J. Jap. Bot. 1: 1–54.Search in Google Scholar
Sapir, A., Dillman, A.R., Connon, S.A., Grupe, B.M., Ingels, J., Mundo-Ocampo, M., Levin, L.A., Baldwin, J.G., Orphan, V.J., and Sternberg, P.W. (2014). Microsporidia-nematode associations in methane seeps reveal basal fungal parasitism in the deep sea. Front. Microbiol. 5: 43, https://doi.org/10.3389/fmicb.2014.00043.Search in Google Scholar PubMed PubMed Central
Sarma, V.V. and Vittal, B.P.R. (2000). Biodiversity of mangrove fungi on different substrata of Rhizophora apiculata and Avicennia spp. from Godavari and Krishna deltas, east coast of India. Fungal Divers. 5: 23–41.Search in Google Scholar
Schmidt, I. (1985). Types and type collections of new higher marine and fresh-water fungi from the Baltic coast. Mycotaxon 24: 419–421.Search in Google Scholar
Senanayake, I.C., Pem, D., Rathnayaka, A.R., Wijesinghe, S.N., Tibpromma, S., Wanasinghe, D.N., Phookamsak, R., Kularathnage, N.D., Gomdola, D., Harishchandra, D., et al.. (2022). Predicting global numbers of teleomorphic ascomycetes. Fungal Divers 114: 237–278.10.1007/s13225-022-00498-wSearch in Google Scholar
Shearer, C.A. and Crane, J.L. (1977). Fungi of the Chesapeake Bay and its tributaries VI. Trichocladium achrasporum, the imperfect state of Halosphaeria mediosetigera, a marine ascomycete. Mycologia 69: 1218–1223.10.1080/00275514.1977.12020186Search in Google Scholar
Shen, M., Zhang, J.Q., Zhao, L.L., Groenewald, J.Z., Crous, P.W., and Zhang, Y. (2020). Venturiales. Stud. Mycol. 96: 185–308, https://doi.org/10.1016/j.simyco.2020.03.001.Search in Google Scholar PubMed PubMed Central
Shinn, A. (2016). Asian shrimp production and the economic costs of disease [Abstract]. In: FAO Technical Cooperation Programme (Ed.), Second international technical seminar/workshop on AHPND: There is a way forward. FAO, Rome, p. 56.Search in Google Scholar
Simmons, D.R., James, T.Y., Meyer, A.F., and Longcore, J.E. (2009). Lobulomycetales, a new order in the Chytridiomycota. Mycol. Res. 113: 450–460, https://doi.org/10.1016/j.mycres.2008.11.019.Search in Google Scholar PubMed
Spegazzini, C. (1881). Fungi argentini additis nonnullis brasiliensibus montevideensibusque. Pugillus quartus (Continuacion). An. Soc. Cient. Argent 12: 174–189.Search in Google Scholar
Spegazzini, C. (1912). Mycetes argentinenses (Series VI). Anales Mus. Nac. Hist. Nat. Buenos Aires 23: 1–146.Search in Google Scholar
Sridhar, K.R. and Maria, G.L. (2003). Diversity of filamentous fungi on woody litter of five mangrove plant species from the southwest coast of India. Fungal Divers. 14: 109–126.Search in Google Scholar
Sridhar, K.R. and Maria, G.L. (2006). Fungal diversity on woody litter of Rhizophora mucronata in a southwest Indian mangrove. Indian J. Mar. Sci. 35: 318–325.Search in Google Scholar
Suetrong, S., Schoch, C.L., Spatafora, J.W., Kohlmeyer, J., Volkmann-Kohlmeyer, B., Sakayaroj, J., Phongpaichit, S., Tanaka, K., Hirayama, K., and Jones, E.B.G. (2009). Molecular systematics of the marine Dothideomycetes. Stud. Mycol. 64: 155–173S6, https://doi.org/10.3114/sim.2009.64.09.Search in Google Scholar PubMed PubMed Central
Summerbell, R.C., Gueidan, C., Guarro, J., Eskalen, A., Crous, P.W., Gupta, A.K., Gené, J., Cano-Lira, J.F., van Iperen, A., Starink, M., et al.. (2018). The protean Acremonium. A. sclerotigenum/egyptiacum: revision, food contaminant, and human disease. Microorganisms 6: 88, https://doi.org/10.3390/microorganisms6030088.Search in Google Scholar PubMed PubMed Central
Swann, E.C., Frieders, E.M., and McLaughlin, D.J. (2001). Urediniomycetes. In: McLaughlin, D.J., McLaughlin, E.G. and Lemke, P.A. (Eds.), Systematics and evolution. The mycota, Vol. 7B. Springer Berlin Heidelberg, Berlin, Heidelberg, pp. 37–56.10.1007/978-3-662-10189-6_2Search in Google Scholar
Tedersoo, L., Sánchez-Ramírez, S., Kõljalg, U., Bahram, M., Döring, M., Schigel, D., May, T., Ryberg, M., and Abarenkov, K. (2018). High-level classification of the Fungi and a tool for evolutionary ecological analyses. Fungal Divers. 90: 135–159, https://doi.org/10.1007/s13225-018-0401-0.Search in Google Scholar
Thambugala, K.M., Ariyawansa, H.A., Li, Y.M., Boonmee, S., Hongsanan, S., Tian, Q., Singtripop, C., Bhat, D.J., Camporesi, E., Jayawardena, R., et al.. (2014). Dothideales. Fungal Divers. 68: 105–158, https://doi.org/10.1007/s13225-014-0303-8.Search in Google Scholar
Thambugala, K.M., Hyde, K.D., Tanaka, K., Tian, Q., Wanasinghe, D.N., Ariyawansa, H.A., Jayasiri, S.C., Boonmee, S., Camporesi, E., Hashimoto, A., et al.. (2015). Towards a natural classification and backbone tree for Lophiostomataceae, Floricolaceae, and Amorosiaceae fam. nov. Fungal Divers. 74: 199–266, https://doi.org/10.1007/s13225-015-0348-3.Search in Google Scholar
Thambugala, K.M., Hyde, K.D., Eungwanichayapant, P.D., Romero, A.I., and Liu, Z.Y. (2016). Additions to the genus Rhytidhysteron in Hysteriaceae. Cryptogam. Mycol. 37: 99–116, https://doi.org/10.7872/crym/v37.iss1.2016.99.Search in Google Scholar
Torres-Garcia, D., García, D., Cano-Lira, J.F., and Gené, J. (2022). Two novel genera, Neostemphylium and Scleromyces (Pleosporaceae) from freshwater sediments and their global biogeography. J. Fungi 8: 868, https://doi.org/10.3390/jof8080868.Search in Google Scholar PubMed PubMed Central
Vargas-Gastélum, L. and Riquelme, M. (2020). The mycobiota of the deep sea: what omics can offer. Life 11: 1–18, https://doi.org/10.3390/life10110292.Search in Google Scholar PubMed PubMed Central
Varrella, S., Barone, G., Tangherlini, M., Rastelli, E., Dell’Anno, A., and Corinaldesi, C. (2021). Diversity, ecological eole and biotechnological potential of antarctic marine fungi. J. Fungi 7: 391, https://doi.org/10.3390/jof7050391.Search in Google Scholar PubMed PubMed Central
Vasilyeva, L.N., Rogers, J.D., and Miller, A.N. (2007). Pyrenomycetes of the Great Smoky Mountains National Park. V. Annulohypoxylon and Hypoxylon (Xylariaceae). Fungal Divers. 27: 231–245.Search in Google Scholar
Vittal, B.P.R. and Sarma, V.V. (2006). Diversity and ecology of fungi on mangroves of Bay of Bengal region – an overview. Indian J. Mar. Sci. 35: 308–317.Search in Google Scholar
Voglmayr, H. and Amengual, A.P. (2018). Three new species of Stigmatodiscus from Mallorca (Spain). Mycol. Prog. 17: 1189–1201, https://doi.org/10.1007/s11557-018-1435-0.Search in Google Scholar
Voglmayr, H., Gardiennet, A., and Jaklitsch, W.M. (2016). Asterodiscus and Stigmatodiscus, two new apothecial dothideomycete genera and the new order Stigmatodiscales. Fungal Divers. 80: 271–284, https://doi.org/10.1007/s13225-016-0356-y.Search in Google Scholar PubMed PubMed Central
Vrijmoed, L.L.P., Hyde, K.D., and Jones, E.B.G. (1996). Melaspilea mangrovei sp. nov., from Australian and Hong Kong mangroves. Mycol. Res. 100: 291–294, https://doi.org/10.1016/s0953-7562(96)80156-0.Search in Google Scholar
Walker, A.K. and Robicheau, B.M. (2021). Fungal diversity and community structure from coastal and barrier island beaches in the United States Gulf of Mexico. Sci. Rep. 11: 3889, https://doi.org/10.1038/s41598-021-81688-5.Search in Google Scholar PubMed PubMed Central
Wanasinghe, D.N., Hyde, K.D., Jeewon, R., Crous, P.W., Wijayawardene, N.N., Jones, E.B.G., Bhat, D.J., Phillips, A.J.L., Groenewald, J.Z., Dayarathne, M.C., et al.. (2017). Phylogenetic revision of Camarosporium (Pleosporineae, Dothideomycetes) and allied genera. Stud. Mycol. 87: 207–256, https://doi.org/10.1016/j.simyco.2017.08.001.Search in Google Scholar PubMed PubMed Central
Wang, X., Singh, P., Gao, Z., Zhang, X., Johnson, Z.I., and Wang, G. (2014). Distribution and diversity of planktonic fungi in the west pacific warm pool. PLoS One 9: e101523, https://doi.org/10.1371/journal.pone.0101523.Search in Google Scholar PubMed PubMed Central
Wang, Q.M., Begerow, D., Groenewald, M., Liu, X.Z., Theelen, B., Bai, F.Y., and Boekhout, T. (2015). Multigene phylogeny and taxonomic revision of yeasts and related fungi in the Ustilaginomycotina. Stud. Mycol. 81: 55–83, https://doi.org/10.1016/j.simyco.2015.10.004.Search in Google Scholar PubMed PubMed Central
Wang, Z.P., Liu, Z.Z., Wang, Y.L., Bi, W.H., Liu, L., Wang, H.Y., Zheng, Y., Zhang, L.L., Hu, S.G., Xu, S.S., et al.. (2019). Fungal community analysis in seawater of the Mariana Trench as estimated by Illumina HiSeq. RSC Adv. 9: 6956–6964, https://doi.org/10.1039/c8ra10142f.Search in Google Scholar PubMed PubMed Central
Wei, X., Guo, S., Gong, L.F., He, G., Pang, K.L., and Luo, Z.H. (2018). Cultivable fungal diversity in deep-sea sediment of the East Pacific Ocean. Geomicrobiol. J. 35: 790–797, https://doi.org/10.1080/01490451.2018.1473531.Search in Google Scholar
Whitton, S.R., McKenzie, E.H.C., and Hyde, K.D. (2003). Microfungi on the Pandanaceae: Zygosporium, a review of the genus and two new species. Fungal Divers. 12: 207–222.Search in Google Scholar
Wijayawardene, N.N., Hyde, K.D., Dai, D.Q., Sánchez-García, M., Goto, B.T., Saxena, R.K., Erdoğdu, M., Selçuk, F., Rajeshkumar, K.C., Aptroot, A., et al.. (2022). Outline of Fungi and fungus-like taxa – 2021. Mycosphere 13: 53–453.10.5943/mycosphere/13/1/2Search in Google Scholar
Xu, W., Guo, S., Pang, K.L., and Luo, Z.H. (2017). Fungi associated with chimney and sulfide samples from a South Mid-Atlantic Ridge hydrothermal site: distribution, diversity and abundance. Deep-Sea Res. I: Oceanogr. Res. Pap. 123: 48–55, https://doi.org/10.1016/j.dsr.2017.03.004.Search in Google Scholar
Yan, L., Hui, N., Simpanen, S., Tudeer, L., and Romantschuk, M. (2020). Simulation of microbial response to accidental diesel spills in basins containing brackish sea water and sediment. Front. Microbiol. 11: 593232, https://doi.org/10.3389/fmicb.2020.593232.Search in Google Scholar PubMed PubMed Central
Zhang, S.N., Abdel-Wahab, M.A., Jones, E.B.G., Hyde, K.D., and Liu, J.K.J. (2019a). Additions to the genus Savoryella (Savoryellaceae), with the asexual morphs Savoryella nypae comb. nov. and S. sarushimana sp. nov. Phytotaxa 408: 195–207, https://doi.org/10.11646/phytotaxa.408.3.4.Search in Google Scholar
Zhang, S.N., Hyde, K.D., Jones, E.B.G., Jeewon, R., Cheewangkoon, R., and Liu, J.K. (2019b). Striatiguttulaceae, a new pleosporalean family to accommodate Longicorpus and Striatiguttula gen. nov. from palms. MycoKeys 49: 99–129, https://doi.org/10.3897/mycokeys.49.30886.Search in Google Scholar PubMed PubMed Central
Zhang, Y., Fournier, J., Phookamsak, R., Bahkali, A.H., and Hyde, K.D. (2013). Halotthiaceae fam. nov. (Pleosporales) accommodates the new genus Phaeoseptum and several other aquatic genera. Mycologia 105: 603–609, https://doi.org/10.3852/11-286.Search in Google Scholar PubMed
Zhao, G., Liu, X., and Wu, W. (2007). Helicosporous hyphomycetes from China. Fungal Divers. 26: 313–524.Search in Google Scholar
Zhao, R.L., Li, G.J., Sánchez-Ramírez, S., Stata, M., Yang, Z.L., Wu, G., Dai, Y.C., He, S.H., Cui, B.K., Zhou, J.L., et al.. (2017). A six-gene phylogenetic overview of Basidiomycota and allied phyla with estimated divergence times of higher taxa and a phyloproteomics perspective. Fungal Divers. 84: 43–74, https://doi.org/10.1007/s13225-017-0381-5.Search in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2023-0032).
© 2023 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Whither marine mycology: the way forward
- Taxonomy and Phylogeny
- Updates on the classification and numbers of marine fungi
- Recent progress in marine mycological research in different countries, and prospects for future developments worldwide
- Neocamarosporium aquaticum (Neocamarosporiaceae, Dothideomycetes), a novel fungus from salt marsh habitats
- Characterization of novel estuarine Ascomycota based on taxonomic and phylogenetic evaluation
- Ecology
- Diversity of fungi isolated from carapace and gut of the marine crab Portunus sanguinolentus in northern waters of Taiwan
- Genetic diversity of culturable fungi associated with scleractinian corals in the Gulf of Thailand
- Diversity of fungi from marine inundated wood from the Bay of Fundy, Nova Scotia, Canada
- Physiology/Biochemistry
- A review of polyunsaturated fatty acids (PUFA) of marine zoosporic microorganisms and their commercialization
- Salinity and temperature affect growth rate of Alphamyces chaetifer and Gorgonomyces haynaldii (Chytridiomycota) isolated from coastal habitats of Taiwan
Articles in the same Issue
- Frontmatter
- In this issue
- Editorial
- Whither marine mycology: the way forward
- Taxonomy and Phylogeny
- Updates on the classification and numbers of marine fungi
- Recent progress in marine mycological research in different countries, and prospects for future developments worldwide
- Neocamarosporium aquaticum (Neocamarosporiaceae, Dothideomycetes), a novel fungus from salt marsh habitats
- Characterization of novel estuarine Ascomycota based on taxonomic and phylogenetic evaluation
- Ecology
- Diversity of fungi isolated from carapace and gut of the marine crab Portunus sanguinolentus in northern waters of Taiwan
- Genetic diversity of culturable fungi associated with scleractinian corals in the Gulf of Thailand
- Diversity of fungi from marine inundated wood from the Bay of Fundy, Nova Scotia, Canada
- Physiology/Biochemistry
- A review of polyunsaturated fatty acids (PUFA) of marine zoosporic microorganisms and their commercialization
- Salinity and temperature affect growth rate of Alphamyces chaetifer and Gorgonomyces haynaldii (Chytridiomycota) isolated from coastal habitats of Taiwan