Mesencephalic GABA neuronal development: no more on the other side of oblivion
-
Suyan Li
Suyan Li is a Research Fellow in the Department of Psychiatry at Harvard Medical School and McLean Hospital. Dr. Li received her Master’s degree in Biology from the Academy of Agriculture Science, China and PhD in Molecular Biology from the Academy of Military Medical Sciences, China. In 2011, Dr. Li joined the Angiogenesis & Brain Development Laboratory for postdoctoral research and is currently exploring multiple aspects of neurovascular interactions and signaling that shape brain development with lasting consequences for the mature brain.
Sampada Joshee is a Research Assistant in the Angiogenesis & Brain Development Laboratory at McLean Hospital since April, 2014. Ms. Joshee received her Bachelor’s degree in Biochemistry from the University of Colorado at Boulder and Master’s degree in Biomedical Sciences with a specialization in Neurobiology from Colorado State University in Fort Collins, Colorado. Ms. Joshee is contributing to several projects in the laboratory with emphasis on studying cellular and molecular mechanisms underlying GABA neuronal development.
Anju Vasudevan is an Assistant Professor of Psychiatry at Harvard Medical School, Director of the Angiogenesis & Brain Development Laboratory and Associate Neuroscientist at McLean Hospital. Dr. Vasudevan received her Master’s degree in Biotechnology from the University of Mysore, India and PhD in Neurobiology from the University of Cologne, Germany. Dr. Vasudevan completed postdoctoral studies in the Department of Neurology at Massachusetts General Hospital/Harvard Medical School. Dr. Vasudevan has received the NARSAD Young and Independent Investigator Awards and NIH awards to pursue her research. Dr. Vasudevan’s research focuses primarily on elucidating fundamental mechanisms regulating forebrain and midbrain development and the underlying causes for dysfunction that leads to neurological and psychiatric conditions.


Abstract
Midbrain GABA neurons, endowed with multiple morphological, physiological and molecular characteristics as well as projection patterns are key players interacting with diverse regions of the brain and capable of modulating several aspects of behavior. The diversity of these GABA neuronal populations based on their location and function in the dorsal, medial or ventral midbrain has challenged efforts to rapidly uncover their developmental regulation. Here we review recent developments that are beginning to illuminate transcriptional control of GABA neurons in the embryonic midbrain (mesencephalon) and discuss its implications for understanding and treatment of neurological and psychiatric illnesses.
Introduction
The midbrain – an important relay center for sensory inputs and motor outputs networking with the forebrain, hindbrain and spinal cord – is a hub of constant activity (1–10). Gamma-aminobutyric acid (GABA) neurons are key players in midbrain neuronal circuitry, robustly networking with glutamatergic and dopaminergic (DA) neurons to shape multiple aspects of behavior. Midbrain GABA neurons function not only as local inhibitory interneurons but also as projection neurons with targets in various brain regions. Along their developmental path, midbrain GABA neurons acquire molecular and functional diversity and can be divided into three categories based on anatomy and function: (1) dorsally located GABA neurons in the superior colliculus (SC) and inferior colliculus (IC); (2) medially located GABA neurons in the midbrain reticular formation (MRF) and periaqueductal gray (PAG); (3) ventrally located GABA neurons in substantia nigra (SN), ventral tegmental area (VTA) and retrorubral field (RRF), (Figure 1A).

(A) Schema of location of mature midbrain GABA neuronal populations in the superior colliculus (SC), inferior colliculus (IC), midbrain reticular formation (MRF), periaqueductal gray (PAG), substantia nigra (SN) and ventral tegmental area (VTA). Midbrain GABA neuronal populations regulate several brain functions, for instance, they are involved in the SC and IC for sensory integration, in the MRF for sleep, PAG for nociception and defensive behavior, SN for motor activity and in the VTA for motivated behavior. (B) Coronal schematic section of the embryonic mesencephalon depicting its patterning along the dorso-ventral axis into roof plate (RP), alar plate (AP), basal plate (BP) and floor plate (FP) on the left side along with dorso-ventral subdivisions on the right (m1–m7). Aq, aqueduct.
The functional diversity of midbrain GABA neurons based on their location is fascinating. Dorsal midbrain GABA neurons together with glutamatergic neurons are involved in processing and incorporation of sensorimotor, visual, auditory, and defensive behavior (11–16). Although GABA neurons are present in all of the seven layered SC, an abundance of GABA neurons is found in the superficial layers when compared to deep and intermediate layers (17–20). The SC is noteworthy for its high GABA content next only to the SN, globus pallidus and hypothalamus in the central nervous system (CNS) (21, 22). Medial midbrain GABA neurons of PAG regulate vocalization, endogenous control of pain sensation, fear, anxiety and aggression (23–25) while those in the MRF have been implicated with sleep-wake state control (26, 27). Ventral midbrain GABA neurons regulate DA neuron activity in the SN and VTA (6, 7, 28–31) and have projection targets similar to DA neurons, to the prefrontal cortex, basal ganglia and other limbic areas (6–10). Additionally, a considerable number of ascending projections from midbrain dopaminergic nuclei are GABAergic in nature (7). Ventral midbrain GABA neurons are thus critical for the function of dopaminergic pathways; have important functional roles in control of voluntary movements, emotion, mood, motivation, processing of appetite and aversive stimuli, addiction and reward behaviors (6, 29, 32–35) and may be involved in the etiology of several neurological and psychiatric disorders including schizophrenia, depression, mood disorders, addiction and Parkinson’s disease. Midbrain GABA neurons can therefore serve as important targets for treatment of neuropsychiatric disorders and for drugs of abuse. Recent evidence reveals that midbrain SN and VTA DA neurons co-release GABA although they do not synthesize it, by using GABA reuptake transporters (mGATs) to recycle extracellular GABA for release (36). Another study examining mechanisms of nicotine addiction shows that activation of midbrain GABA neurons in the VTA controls nicotine elicited burst firing of DA neurons and points to a concerted role for GABA neurons and DA neurons in mediating nicotine reinforcement (35). Thus, mechanisms adopted by DA neurons to locally regulate GABAergic transmission (36) or GABAergic regulation of DA neuron activity (35) illustrates the complexity of midbrain GABA-DA neuron interactions.
Despite the functional significance of GABA neurons in the adult midbrain, progress made in identifying the mechanisms underlying GABA neuron development has been relatively slow. Compared with the embryonic forebrain (37–40) and developing dorsal spinal cord (41–46) in which tremendous advances have been made in understanding GABA neuron development, in the embryonic midbrain, we are a decade behind. In the mesencephalon, DA neurons have been in the spotlight due to discovery of consequences of midbrain DA neurons degeneration in Parkinson’s disease (PD) and dopamine related neuropsychiatric disorders (47–54). Therefore the molecular mechanisms that define DA neuron development – generation, specification and differentiation of DA neurons have been extensively studied to generate cell replacement and pharmacological intervention strategies to alleviate some of the disease symptoms. However, proper migration of GABA neurons to their final location in the ventral mesencephalon during embryonic development seems to be dependent on the complete DA neuron architecture in the ventral mesencephalon, signifying important interactions between them for final location and connectivity (55). Thus the embryonic developmental period offers a favorable milieu when mesencephalic GABA-DA neuron interactions form and establish. Clearly it is a very exciting time to bring mesencephalic GABA neuron development for study onto a common platform with DA neuron development. In this mini-review we discuss current knowledge about origin of mesencephalic GABA neurons and molecular mechanisms involved in their specification, proliferation, differentiation and migration and emphasize missing links where more work is needed. Generation of detailed maps of key regulators of mesencephalic GABA neuron fate and function will initiate new studies to screen for their failure in mouse and human pathology and help develop new therapeutic strategies that focus on co-ordinate rescue of GABA-DA neuron interactions and novel pharmacological intervention paradigms.
Origin of mesencephalic GABA neurons
The embryonic mesencephalon is patterned along the dorso-ventral axis into roof plate (RP), alar plate (AP), basal plate (BP) and floor plate (FP) by BMP/Wnt signals from the RP and Shh signals from the FP/BP (56–59) and further divided into seven dorso-ventral subdivisions (m1–m7) with specific gene expression codes (60–62) to ensure cellular diversity (Figure 1B). Domains m1–m3 are structured into layers and domains m4–m7 are organized into distinct nuclei. For spatial patterning at dorso-ventral levels of the mesencephalon, several GABAergic progenitor domains have been identified, defined by expression of transcription factors such as Nkx6-1, Nkx2-2 and Pax3/7 (63). GAD expression, starting at E10.5 has been reported in BP and AP (64, 65). Both ventro-lateral and dorsal regions of the mesencephalon produce GABA neurons at E10.5–E14.5 (61, 66) and laminar organization develops during E13–E17 (67). The ventral mesencephalon at E13 is still completely devoid of GABA neurons, but by E17, GABA neurons are found robustly intermingled with DA neurons (55, 68).
GABA neurons originate from five of the seven mesencephalic progenitor domains (m1, m2, m3, dorsal half of m4 and m5). GABA neurons of the dorsal midbrain arise from the m1 and m2 domains. Domains m3–m5 may give rise to GABA neurons of the medial midbrain, MRF and PAG. GABA neurons in the ventral midbrain seem to have two developmental origins: 1) outside the midbrain and 2) from BP region. An early study has reported that all SN neurons in the rat brain originate from the midbrain-hindbrain border known as the isthmus (rostral most rhombomere 1) (69). More recent fate mapping studies have shown that midbrain VTA and SN pars reticulata (SNpr) GABA neurons originate from rhombomere 1 (r1), whereas the most anterior part of SNpr has a distinct origin outside the midbrain, possibly in the diencephalon (68). We have observed many GFP-positive GABA neurons in GAD65-GFP mesencephalon oriented ventrally in BP region at E13 and a stream of these neurons coursing from BP region to the ventral mesencephalon by E17. BrdU birthdating experiments additionally revealed that many E11-labeled neuronal progenitors migrated to contribute to GABA neurons of the ventral mesencephalon by E17 (55). The origin of these migrating cells remains to be better clarified.
Specification of GABAergic neuronal fate in the mesencephalon
Combinations of multiple transcription factors act as first selectors of neuronal fate (Figures 2 and 3) and contribute to neuronal diversity. In the mesencephalon, GABA neuronal fate determination is primarily associated with two basic helix-loop-helix (bHLH) genes: Helt (also known as Megane or Heslike) and Ascl1 (also known as Mash1) (61, 65, 70–72).

Overview of the molecular codes essential for generation of mesencephalic GABA neurons in relation to their specific position in the ventricular zone (VZ), intermediate zone (IZ) and mantle zone (MZ).
Beginning with inductive signals, a specific cascade of molecular instruction follows in the VZ, IZ and MZ to ensure GABAergic fate specification, neurogenesis, fate maintenance and differentiation into mature GABA neuronal populations.
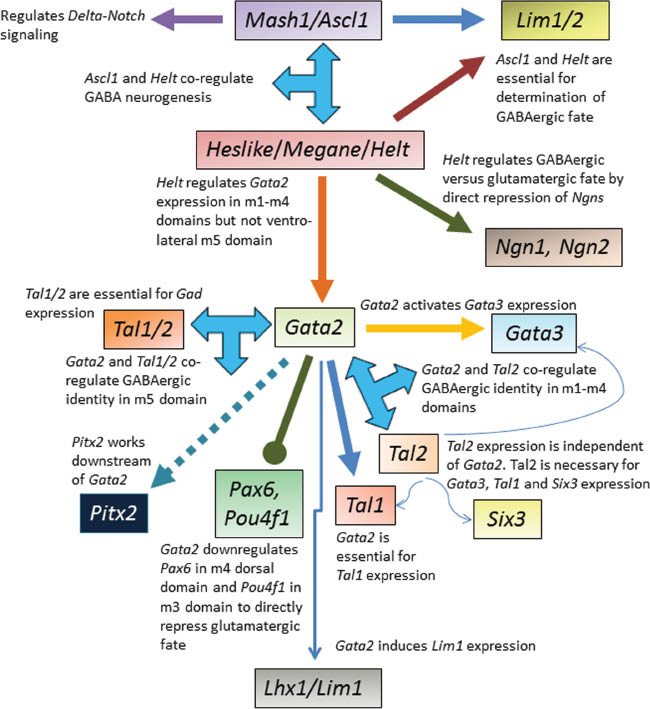
Summary of the signaling pathway of key molecular players identified up until now and their interplay that is essential for efficient generation of mesencephalic GABA neurons.
Helt is co-expressed with Ascl1 in the ventricular zone of the alar-basal mesencephalic boundary from E9.5 onwards. Subsequently the Helt- Ascl1 co-expression domain expands dorsally as well. Gain of function studies in which transgenic mice were generated mis-expressing Helt from the nestin promoter-enhancer resulted in ectopic GABA neurons in the mantle layer outside the Helt/Ascl1+ zone (70). As the effect of mis-expression of Helt was specific to the mesencephalon and no ectopic cells were observed in other brain regions (dorsal telencephalon and thalamus) which do not express Ascl1, Helt was initially believed to specify the GABA neuronal fate only when Ascl1 was co-expressed (70). Interestingly, although ectopic Helt expression induced GABA neurons, it also suppressed the generation of glutamatergic neurons (61). Loss of function studies revealed that in homozygous Helt-null mice GABAergic progenitors were generated (61, 71) but they failed to become GABA neurons specifically in the dorsal mesencephalon. Ventral GABA neurons formed only from the m5 domain in Helt-null mesencephalon whereas in the dorsal domains there was a complete loss of GABA neurons and instead, glutamatergic neurons, induced by neurogenin (Ngn) genes emerged (61). Helt is now recognized as a key determinant of the GABAergic fate in dorsal mesencephalon by direct repression of Ngn genes (Ngn1 and Ngn2) in GABAergic progenitors, and induction of Lim1/2 and Gata2 (60, 61). Helt is required for Gata2 expression in the embryonic mesencephalon except in the ventro-lateral m5 domain (60). Loss of function of Helt did not affect dopaminergic and cholinergic neurons. Thus the essential role of Helt for regulating GABA neuron fate specification in the dorsal midbrain and for development of the SC was confirmed.
Ascl1, expressed by neural progenitors in all mesencephalic progenitor domains is another GABAergic fate determinant that is not regulated by Helt or other proneural genes such as Ngns. Ascl1 was unaffected in the Helt-null mice and may have also compensated for loss of Helt in ventral but not dorsal mesencephalon (70). In Ascl1-null mice, virtually no GABA neurons formed in the mesencephalon up until E11.5 while Helt continued to be expressed and other neurons were generated. From E13.5 onwards, GABA neurons were produced in the ventromedial mesencephalon (m3–m5), but not dorsally (m1–m2). By E18.5, GABA neurons were completely lost in Ascl1-null dorsal mesencephalon including SC, IC and dorsal PAG. Although GABA neurons were reduced in medial mesencephalon including ventral PAG and MRF, GABA neurons of the ventral mesencephalon in SN and VTA were not affected and do not seem to require Ascl1 (72).
Thus ventral mesencephalic GABA neurons represent an intriguing population of GABA neurons that develop independently of regulation by Helt and Ascl1.
Mesencephalic GABA neurogenesis
GABAergic neurogenesis is completed first in the MRF by E12.5, next in the SNpr by E13.5, followed by SC at E14.5 (68) and has been associated mostly with transcription factor combinations Helt and Ascl1 (Figures 2 and 3). The Helt-Ascl1 co-expression domain in the mesencephalon is co-ordinate with GABA neurogenesis, decreasing rapidly as neuronal differentiation proceeds (70). However, Ascl1’s requirement for neurogenesis seems to be highly region-specific. In Ascl1-null mesencephalon, a loss of neurogenic and neural stem cell specific expression with markers Delta1 (Notch ligand) and Hes5 (Notch target) was observed at early embryonic stages. This delay at the start of neurogenesis is believed to cause reduced GABA neuron numbers in the ventromedial midbrain of Ascl1-null mice (72). Ascl1 has been reported to promote GABAergic neurogenesis in in vitro cultured mesencephalic neural progenitors (73). Co-expression of both Ascl1 and Helt seems to significantly promote GABA neurogenesis in neural precursor cell cultures (70).
Mesencephalic GABA neuron differentiation and fate maintenance
After the cell cycle exit, the GABA neuron precursors activate genes typical for functional GABA neuron precursors. This role is assigned to C4 zinc-finger transcription factor Gata2, bHLH transcription factors Tal1/2, paired-like homeodomain transcription factor Pitx2 and homeobox transcription factor Lhx1 (Lim1) (Figures 2 and 3). Gata2 is expressed in m1–m5 domains in the embryonic mesencephalon and expression is activated in GABAergic progenitors as they exit the cell cycle, become postmitotic precursor cells and begin to differentiate (60). Selective loss of Gata2 in the mouse midbrain and rhombomere 1 (Gata2cko mutant) resulted in a specific loss of GABA neuron precursors at early embryonic stages and transformation from GABAergic to glutamatergic phenotype in all mesencephalic GABA neuron subpopulations in the Gata2cko mutant during embryonic development except for the GABA neurons associated with DA neurons in the SNpr and VTA that were unaffected (60, 63). Tal1/2 are expressed in m1–m4 dorsal half and m5 mesencephalic domains and co-operatively activate genes necessary for GABA neuronal differentiation (74). Tal2 is co-expressed with Gata2 in the ventricular zone (VZ) and intermediate zone (IZ) and regulates selection of GABAergic over glutamatergic neuronal fate. An ectopic upregulation of glutamatergic gene expression was observed in Tal2cko mutants in m3 and m4 dorsal domains (74). Similar to Gata2cko mutant, in Tal2cko mutant, GABAergic markers were completely lost in the dorsal mesencephalon including SC. Again, both Gata2 and Tal2 are dispensable for ventral mesencephalic GABA neurons (74).
Pitx2, is expressed in m1–m4 and m6 domains with highest expression in the intermediate layer of the SC (75) and works downstream of Gata2 in the transcription factor cascade (60, 75). Loss of Pitx2 disrupts GABAergic neuronal differentiation and axonal outgrowth in the dorsal mesencephalon, specifically SC (76). In the ventral mesencephalon Pitx2 lineage neurons are mostly glutamatergic (75). Lhx1 (or Lim1) is expressed in postmitotic precursors of all mesencephalic GABA neurons in domains m1–m5 (60) and is an important marker of GABAergic differentiation.
Migration of mesencephalic GABA neurons
Neuronal migration is a key event during brain development as neurons and/or neuronal progenitors originating in VZs navigate along diverse courses to eventually find their destination and integrate into specific brain circuits. In the pallial telencephalon, pyramidal neurons follow radial migratory routes using radial glial guides to the cortical plate (40, 77). GABA neurons of the subpallial telencephalon, conversely, take long tangential paths into the cortex (78, 79) along vascular guides (80) to become intermingled with excitatory neurons. Unlike in the embryonic telencephalon where neuronal migration has been well elucidated, this vital event has been little explored in the embryonic mesencephalon. In the dorsal mesencephalon, majority of GABA neurons have been reported to migrate radially from the dorsal VZ to their final location in superficial layers through deep and intermediate layers in an inside-out manner (81) followed by slight tangential dispersion within the superficial layer (Figure 4A). Few cells here (81) show direct tangential migration from VZ to the superficial layer of the SC (Figure 4A). Three types of inhibitory neurons – stellate cells, pyriform cells and horizontal cells with tangentially oriented dendrites – thus come to reside in the multilayered SC (20).

(A) Coronal view of embryonic mesencephalon showing different patterns of neuronal migration. Black arrows depict radial migration of GABA neurons from the VZ to their final location. Green arrows indicate slight tangential dispersion within the superficial layer of the SC. Red arrow depicts that few cells show direct tangential migration from VZ to the superficial layer of the SC. Blue arrows highlight perpendicular migration of GABA neurons from basal plate region to ventral mesencephalon. (B) Sagittal view of embryonic mesencephalon depicting migration of postmitotic GABA neuronal precursors that cross the specific mesencephalon-rhombomere1 (r1) boundary to enter from r1 into the ventral mesencephalon (vm). The mode of migration of these neurons with origins outside the mesencephalon is currently unknown. tel, Telencephalon; mes, Mesencephalon; SC, superior colliculus.
In the ventral mesencephalon, GABA neuronal migration seems to be more complex with contributions from both outside and within. Recent genetic fate mapping studies have uncovered a separate origin of VTA and SNpr GABA neurons outside the mesencephalon (68). The findings unveil a new migratory pathway of GABA neurons originating in r1 compartment and crossing the r1-mesencephlon boundary to migrate to the VTA and SNpr as postmitotic neuronal precursors at E14.5–E15.5 (Figure 4B). Guidance mechanisms for this form of migration remain to be elucidated. The anterior SNpr GABA neurons in the diencephalon however do not arrive from the r1 region; they are speculated to come from the diencephalon although contribution from the mesencephalon is not excluded (68). These results were further supported by analysis of Tal1 mutant mice in which GABAergic precursors are normal in the mesencephalon, but failure in GABA neuron production in r1 correlated with loss of mature VTA and SNpr GABA neurons (68). Additionally, as Gata2 regulates GABA neuronal differentiation in the mesencephalon but not r1, in Gata2 mutants, although mesencephalic precursors failed to activate GABAergic neuron-specific gene expression, the VTA and SNpr GABA neurons appeared largely unaffected (60), supporting their outside origin and border migration across r1-mesencephalon compartments.
Our studies have shown that both DA and GABA neurons occupy ventral mesencephalon in a temporally sequential manner during embryonic development and depicted GABA neuronal silhouette oriented from BP region to ventral mesencephalon in GAD65-GFP embryos (55). It emphasized the importance of perpendicular migration, a novel mode of neuronal migration that seems to be unique to the mesencephalon (Figure 4A) and essential for proper set up of the anatomical architecture of ventral mesencephalic structures (55). BrdU birthdating experiments revealed that E10 and E11 labeled neuronal progenitors migrated ventrally (from FP) as well as perpendicular (from BP) to the aqueduct to form the distinct anatomical architecture of the boat shaped SN and VTA by E17 (55). Pitx3 represents a critical regulator of mesencephalic DA neuron development (49, 51, 82–85) and there is selective and early loss of A9 DA neurons in the SN of ak/ak mice (49, 84–86). The cells affected in the ak/ak mice are known to be very sensitive to neurotoxins such as 6-OHDA and MPTP (54, 87) and are the same cells that degenerate first in Parkinson’s disease (PD) patients (83–85). Our findings have revealed that loss of SN DA neurons in ak/ak mesencephalon is a result of defective perpendicular migration that resulted in cells which were stuck or trailing in the middle of their migratory trajectory and distributed abnormally in the red nucleus area. Interestingly, coordinate with this loss of DA neurons there was also a significant loss of ventral mesencephalic GABA neurons that persisted in the adult ak/ak midbrain (55). Pre-existing DA neurons seem to modulate the migration of BP GABA neurons to ventral mesencephalon along perpendicular migration routes suggestive of important interactions between both neuronal populations for final location and connectivity and the earliest signs of inter-dependence that arises during embryonic development (55). Identification of the anatomical/cellular substrate for perpendicular neuronal migration will help model future investigations to induce this intriguing population of GABA neurons to migrate successfully in cell transplantation experiments coordinate with DA neurons.
At a mechanistic level, neuronal migration overall in the mesencephalon suffers greatly from lack of insights. Mesencephalic axon guidance and/or ventral migration of DA neurons has been linked only to a small number of molecules [netrin 1 (88) and its receptor DCC (89), reelin (90), proteoglycan phosphacan 6B4 (91), L1CAM (92), neural cell adhesion molecule (93) and polysialic acid (93)]. Transcription factors, primary mechanisms controlling neuronal migration, remain unidentified in the embryonic mesencephalon. For perpendicular migration to occur from the BP to the ventral mesencephalon, neurons need to perfectly synchronize multiple actions and respond appropriately to guidance cues encountered during their trajectory. Transcriptional regulation is inevitably a key determinant of this process. In the vertebrate spinal cord, for instance, expression of specific combinations of transcription factors in postmitotic motorneurons encodes axon trajectories (94–96) and similar transcription factors carry on different functions depending on the cellular context (97–99). Another example is that of postmitotic Nkx2.1 expression, which has to be down-regulated for interneurons to migrate to the cortex, whereas Nkx2.1 expression is required for interneuron migration to the striatum (100). Similar possibilities lie in the embryonic mesencephalon and much work lies ahead with respect to elucidating mechanisms of neuronal migration.
Conclusion and perspectives
The generation of several mouse models with abnormal development and function of cortical GABA interneurons, which recapitulated defective behavior similar to those seen in conditions like schizophrenia, autism, epilepsy, mood and anxiety disorders, was a major step that associated forebrain GABA neurons with the pathobiology of several neuropsychiatric illnesses (101–107). Thus, the identification of the molecular components involved in forebrain GABA neuron development in both mouse and human triggered efficient generation of GABA neuron populations based on ES cell engineering. ES-derived GABA interneurons today have remarkable potential – they are functional, survive well, migrate and integrate into desired regions in both embryonic and adult brains post-transplantation and are attractive candidates for cell-based therapies (108–112). In the midbrain, ES cell technology today focuses selectively to generate DA neurons (113–116). This is possible because the molecular cascade of DA neuron development has been well studied. A missing link in the midbrain field is our current incapability for efficient generation of GABA neurons from ES cells, as the molecular mechanisms underlying mesencephalic GABA neuron fate and function are just beginning to be elucidated and furthermore the transcriptional machinery identified here, except for Ascl1 is different from that of the forebrain.
The quest to unravel the transcriptional cascade in the embryonic midbrain is however no easy task. It is particularly daunting given the intricate diversity of GABA neurons found in the mesencephalon based on location and function and transcriptional factor networks, with overlapping function for GABAergic vs. glutamatergic fate, or GABAergic vs. serotonergic/dopaminergic fate depending on their domain. Table 1 summarizes expression stages, domains of expression and function of key molecular players regulating mesencephalic GABA neuron development. As most of these candidates are broadly expressed in several brain regions as well as other organs, systemic knockouts die prenatally or at birth (Table 1). There is a great need for gain and loss of function assays, including generation of more mouse mutants with deletion of GABA neuron-specific genes selectively in the embryonic mesencephalon and subsequent analysis of brain development and postnatal behavior to be able to fully understand the developmental significance of mesencephalic GABA neurons in the CNS. Helt, Gata2 and Tal1/2 are transcription factors that have been best studied in this regard (Table 1). Given Helt’s unique expression pattern in the mesencephalon and prosomere1-2 in the diencephalon, isolation of Helt served as a vital key to unlock the mysteries of mesencephalic GABA neuronal development. Helt deletion resulted in loss of weight and postnatal lethality in mice around 3–5 weeks of age with behavioral defects such as fore/hindlimb clenching and seizure-like phenotype from P14 onward (71). Elegant work demonstrating conditional inactivation of Gata2 and Tal1/2 in mouse midbrain and r1 have provided several new insights into the specific role of these transcription factors for GABA neuron development in the embryonic midbrain, although postnatal phenotypes have not yet been reported (60, 74).
Key molecular players regulating mesencephalic GABA neuronal development.
| Gene | Earliest stage of expression and expression domains in the embryonic mesencephalon | Specific function in the mesencephalon | All brain regions where genes are expressed | Phenotypes of homozygous null mutants/transgenic mice |
|---|---|---|---|---|
| Heslike/Megane/Helt | E9.5: alar-basal boundary E9.5–E12.5 onward: further dorsal expansion towards roof plate. Mostly absent from ventral mesencephalon. Expression in VZ, not detected in mantle zone. E13.5–E17.5: expression is down-regulated and disappears by 17.5 | 1) Specifies GABA neuron fate in dorsal mesencephalon 2) Possible role in GABA neurogenesis 3) Essential for GABA neuronal development in SC | 1) Diencephalon (p1–2) 2) Mesencephalon | Homozygous Helt-null mice show behavioral defects, seizure-like activity and postnatal lethality at 3–5 weeks after birth Transgenic mice mis-expressing Helt in the Ascl1 region die at E12.5 |
| Mash1/Ascl1 | E9.5: alar-basal boundary and dorsal mesencephalon, E10.5: both domains connect E11.5–E12.5 onward: further dorsal (m1–m2) and ventral (m3–m5) expansion of the domain Expression in VZ; overlaps with Helt, also expressed in the MZ | 1) Specification of GABA neuronal fate in dorsal mesencephalon 2) GABA neurogenesis specifically in the dorsal mesencephalon and partially in the ventro-medial mesencephalon | 1) Telencephalon 2) Diencephalon 3) Mesencephalon 4) R1 5) Developing spinal cord | As Ascl1 shows broad expression in the CNS, PNS and in neuroendocrine cells in lung and kidney, homozygous Ascl1-null mice die soon after birth with severe disruptions in several aspects of neuronal development |
| Gata2 | E10.5–12.5: expression in basal plate region in IZ and MZ E12.5 onward: expression appears in dorsal mesencephalon. Expression coincides with midbrain regions producing Gad1 positive neurons throughout development | Functions as an essential postmitotic selector selector gene for GABAergic over glutamatergic identity | 1) Diencephalon (p1–2) 2) Mesencephalon 3) Ventral spinal cord | Gata2 homozygous null mutant embryos die at E10.5 during embryogenesis. Conditional inactivation of Gata2 in midbrain and r1 depicts its specific role in GABA neuronal development in midbrain and not r1. Postnatal phenotype not reported |
| Tal1/2 | E10.5–E12.5 onward: Tal1 expression confined to IZ and MZ; co-expressed with Gata2 in the IZ; co-expressed with Gata3 in the MZ. Tal2 expressed in VZ and IZ and co-expressed with Gata2. Expressed in m1-m4 dorsal half and m5 domains | 1) Tal1/2 co-operatively guides GABA neuron differentiation 2) Tal2 is important for sub-type specification in postmitotic neurons; selection of GABAergic over glutamatergic neuron fate | 1) Diencephalon 2) Mesencephalon 3) Ventral R1 4) Ventral spinal cord | Tal1 gene knockout results in mid-gestational lethality due to lack of yolk sac erythropoiesis. Conditional inactivation of Tal1/2 in the embryonic midbrain highlights its role in postmitotic GABA neuronal differentiation. Postnatal phenotype not reported |
| Pitx2 | E10.5 onward: expression limited to postmitotic neurons of the mantle zone, in domains m1–m4 and m6. Highest expression in intermediate layer of SC | Essential for GABA neuronal differentiation and axonal outgrowth in the dorsal midbrain, specially SC | 1) Mesencephalon 2) Ventral R1 | Pitx2 is essential for normal development of several organs including pituitary and eyes. Complete loss of Pitx2 in mice results in embryonic lethality by E15 due to cardiac defects |
The VTA and SNpr GABA neurons are particularly distinct and interesting because they develop independently of the known regulators of midbrain GABAergic fate specification and maintenance: Helt, Ascl1, Gata2 and Tal2 (60, 61, 65, 71, 72, 74). Given the functional importance of GABA-DA neuron associations in the adult midbrain, it has become critical to decode transcriptional regulation of ventral mesencephalic GABA neurons as well as delineate development of GABA-DA neuron interactions and connectivity. Additionally, as GABA neurons were also affected in addition to DA neurons as a result of loss of Pitx3 (55), a crucial transcriptional regulator of DA neuron development, there lies the possibility of overlapping molecular pathways and shared mechanisms mediating GABA-DA neuron development in the ventral mesencephalon. Many vital goals remain to be achieved before we can explore and combine the distinctive features of both GABA neurons and DA neurons so as to understand complex neurological and psychiatric illnesses and to test novel therapies that can ultimately become a clinical reality. Clearly, the field of mesencephalic GABA neuron development is poised for several exciting discoveries in the future.
About the authors

Suyan Li is a Research Fellow in the Department of Psychiatry at Harvard Medical School and McLean Hospital. Dr. Li received her Master’s degree in Biology from the Academy of Agriculture Science, China and PhD in Molecular Biology from the Academy of Military Medical Sciences, China. In 2011, Dr. Li joined the Angiogenesis & Brain Development Laboratory for postdoctoral research and is currently exploring multiple aspects of neurovascular interactions and signaling that shape brain development with lasting consequences for the mature brain.

Sampada Joshee is a Research Assistant in the Angiogenesis & Brain Development Laboratory at McLean Hospital since April, 2014. Ms. Joshee received her Bachelor’s degree in Biochemistry from the University of Colorado at Boulder and Master’s degree in Biomedical Sciences with a specialization in Neurobiology from Colorado State University in Fort Collins, Colorado. Ms. Joshee is contributing to several projects in the laboratory with emphasis on studying cellular and molecular mechanisms underlying GABA neuronal development.

Anju Vasudevan is an Assistant Professor of Psychiatry at Harvard Medical School, Director of the Angiogenesis & Brain Development Laboratory and Associate Neuroscientist at McLean Hospital. Dr. Vasudevan received her Master’s degree in Biotechnology from the University of Mysore, India and PhD in Neurobiology from the University of Cologne, Germany. Dr. Vasudevan completed postdoctoral studies in the Department of Neurology at Massachusetts General Hospital/Harvard Medical School. Dr. Vasudevan has received the NARSAD Young and Independent Investigator Awards and NIH awards to pursue her research. Dr. Vasudevan’s research focuses primarily on elucidating fundamental mechanisms regulating forebrain and midbrain development and the underlying causes for dysfunction that leads to neurological and psychiatric conditions.
Acknowledgments
Supported by a National Alliance for Research on Schizophrenia and Depression (NARSAD) Independent Investigator Award and National Institutes of Health grant R01NS073635 to AV.
References
1. Mehler WR, Feferman ME, Nauta WJ. Ascending axon degeneration following anterolateral cordotomy. An experimental study in the monkey. Brain 1960; 83: 718–50.10.1093/brain/83.4.718Suche in Google Scholar
2. Garey LJ, Jones EG, Powell TP. Interrelationships of striate and extrastriate cortex with the primary relay sites of the visual pathway. J Neurol Neurosurg Psychiatry 1968; 31: 135–57.10.1136/jnnp.31.2.135Suche in Google Scholar
3. Valverde F. The neuropil in superficial layers of the superior colliculus of the mouse. A correlated Golgi and electron microscopic study. Z Anat Entwicklungsgesch 1973; 142: 117–47.10.1007/BF00519719Suche in Google Scholar
4. Meredith MA, Stein BE. Visual, auditory, and somatosensory convergence on cells in superior colliculus results in multisensory integration. J Neurophysiol 1986; 56: 640–62.10.1152/jn.1986.56.3.640Suche in Google Scholar
5. Wickelgren BG. Superior colliculus: some receptive field properties of bimodally responsive cells. Science 1971; 173: 69–72.10.1126/science.173.3991.69Suche in Google Scholar
6. Laviolette SR, van der Kooy D. The neurobiology of nicotine addiction: bridging the gap from molecules to behaviour. Nat Rev Neurosci 2004; 5: 55–65.10.1038/nrn1298Suche in Google Scholar
7. Fields HL, Hjelmstad GO, Margolis EB, Nicola SM. Ventral tegmental area neurons in learned appetitive behavior and positive reinforcement. Annu Rev Neurosci 2007; 30: 289–316.10.1146/annurev.neuro.30.051606.094341Suche in Google Scholar
8. Swanson LW. The projections of the ventral tegmental area and adjacent regions: a combined fluorescent retrograde tracer and immunofluorescence study in the rat. Brain Res Bull 1982; 9: 321–53.10.1016/0361-9230(82)90145-9Suche in Google Scholar
9. Oades RD, Halliday GM. Ventral tegmental (A10) system: neurobiology. 1. Anatomy and connectivity. Brain Res 1987; 434: 117–65.10.1016/0165-0173(87)90011-7Suche in Google Scholar
10. Albin RL, Young AB, Penney JB. The functional anatomy of basal ganglia disorders. Trends Neurosci 1989; 12: 366–75.10.1016/0166-2236(89)90074-XSuche in Google Scholar
11. Dean P, Redgrave P, Westby GW. Event or emergency? Two response systems in the mammalian superior colliculus. Trends Neurosci 1989; 12: 137–47.10.1016/0166-2236(89)90052-0Suche in Google Scholar
12. Westby GW, Keay KA, Redgrave P, Dean P, Bannister M. Output pathways from the rat superior colliculus mediating approach and avoidance have different sensory properties. Exp Brain Res 1990; 81: 626–38.10.1007/BF02423513Suche in Google Scholar
13. Oliver DL, Winer JA, Beckius GE, Saint Marie RL. Morphology of GABAergic neurons in the inferior colliculus of the cat. J Comp Neurol 1994; 340: 27–42.10.1002/cne.903400104Suche in Google Scholar
14. Merchan M, Aguilar LA, Lopez-Poveda EA, Malmierca MS. The inferior colliculus of the rat: quantitative immunocytochemical study of GABA and glycine. Neuroscience 2005; 136: 907–25.10.1016/j.neuroscience.2004.12.030Suche in Google Scholar
15. Kaneda K, Phongphanphanee P, Katoh T, Isa K, Yanagawa Y, Obata K, Isa T. Regulation of burst activity through presynaptic and postsynaptic GABAB receptors in mouse superior colliculus. J Neurosci 2008; 28: 816–27.10.1523/JNEUROSCI.4666-07.2008Suche in Google Scholar
16. Ito T, Oliver DL. The basic circuit of the IC: tectothalamic neurons with different patterns of synaptic organization send different messages to the thalamus. Front Neural Circuits 2012; 6: 48.10.3389/fncir.2012.00048Suche in Google Scholar
17. Harvey AR, Heavens RP, Yellachich LA, Sirinathsinghji DJ. Expression of messenger RNAs for glutamic acid decarboxylase, preprotachykinin, cholecystokinin, somatostatin, proenkephalin and neuropeptide Y in the adult rat superior colliculus. Neuroscience 2001; 103: 443–55.10.1016/S0306-4522(00)00581-9Suche in Google Scholar
18. Binns KE. The synaptic pharmacology underlying sensory processing in the superior colliculus. Prog Neurobiol 1999; 59: 129–59.10.1016/S0301-0082(98)00099-9Suche in Google Scholar
19. Jeon CJ, Gurski MR, Mize RR. Glutamate containing neurons in the cat superior colliculus revealed by immunocytochemistry. Vis Neurosci 1997; 14: 387–93.10.1017/S0952523800011500Suche in Google Scholar
20. Mize RR. The organization of GABAergic neurons in the mammalian superior colliculus. Progr Brain Res 1992; 90: 219–48.10.1016/S0079-6123(08)63616-XSuche in Google Scholar
21. Okada Y, Nitsch-Hassler C, Kim JS, Bak IJ, Hassler R. Role of aminobutyric acid (GABA) in the extrapyramidal motor system. 1. Regional distribution of GABA in rabbit, rat, guinea pig and baboon CNS. Exp Brain Res 1971; 13: 514–8.10.1007/BF00234282Suche in Google Scholar
22. Fahn S, Cote LJ. Regional distribution of gamma-aminobutyric acid (GABA) in brain of the rhesus monkey. J Neurochem 1968; 15: 209–13.10.1111/j.1471-4159.1968.tb06198.xSuche in Google Scholar
23. Bandler R, Keay KA, Floyd N, Price J. Central circuits mediating patterned autonomic activity during active vs. passive emotional coping. Brain Res Bull 2000; 53: 95–104.10.1016/S0361-9230(00)00313-0Suche in Google Scholar
24. Behbehani MM. Functional characteristics of the midbrain periaqueductal gray. Prog Neurobiol 1995; 46: 575–605.10.1016/0301-0082(95)00009-KSuche in Google Scholar
25. Barbaresi P. Postnatal development of GABA-immunoreactive neurons and terminals in rat periaqueductal gray matter: a light and electron microscopic study. J Comp Neurol 2010; 518: 2240–60.10.1002/cne.22329Suche in Google Scholar
26. Luppi PH, Clement O, Sapin E, Gervasoni D, Peyron C, Leger L, Salvert D, Fort P. The neuronal network responsible for paradoxical sleep and its dysfunctions causing narcolepsy and rapid eye movement (REM) behavior disorder. Sleep Med Rev 2011; 15: 153–63.10.1016/j.smrv.2010.08.002Suche in Google Scholar
27. Brischoux F, Mainville L, Jones BE. Muscarinic-2 and orexin-2 receptors on GABAergic and other neurons in the rat mesopontine tegmentum and their potential role in sleep-wake state control. J Comp Neurol 2008; 510: 607–30.10.1002/cne.21803Suche in Google Scholar
28. Omelchenko N, Sesack SR. Ultrastructural analysis of local collaterals of rat ventral tegmental area neurons: GABA phenotype and synapses onto dopamine and GABA cells. Synapse 2009; 63: 895–906.10.1002/syn.20668Suche in Google Scholar
29. Tepper JM, Lee CR. GABAergic control of substantia nigra dopaminergic neurons. Progr Bain Res 2007; 160: 189–208.10.1016/S0079-6123(06)60011-3Suche in Google Scholar
30. Margolis EB, Toy B, Himmels P, Morales M, Fields HL. Identification of rat ventral tegmental area GABAergic neurons. PLoS One 2012; 7: e42365.10.1371/journal.pone.0042365Suche in Google Scholar PubMed PubMed Central
31. Nair-Roberts RG, Chatelain-Badie SD, Benson E, White-Cooper H, Bolam JP, Ungless MA. Stereological estimates of dopaminergic, GABAergic and glutamatergic neurons in the ventral tegmental area, substantia nigra and retrorubral field in the rat. Neuroscience 2008; 152: 1024–31.10.1016/j.neuroscience.2008.01.046Suche in Google Scholar
32. Cohen JY, Haesler S, Vong L, Lowell BB, Uchida N. Neuron-type-specific signals for reward and punishment in the ventral tegmental area. Nature 2012; 482: 85–8.10.1038/nature10754Suche in Google Scholar
33. Jhou TC, Fields HL, Baxter MG, Saper CB, Holland PC. The rostromedial tegmental nucleus (RMTg), a GABAergic afferent to midbrain dopamine neurons, encodes aversive stimuli and inhibits motor responses. Neuron 2009; 61: 786–800.10.1016/j.neuron.2009.02.001Suche in Google Scholar
34. Vargas-Perez H, Ting AKR, Walton CH, Hansen DM, Razavi R, Clarke L, Bufalino MR, Allison DW, Steffensen SC, van der Kooy D. Ventral tegmental area BDNF induces an opiate-dependent-like reward state in naive rats. Science 2009; 324: 1732–4.10.1126/science.1168501Suche in Google Scholar
35. Tolu S, Eddine R, Marti F, David V, Graupner M, Pons S, Baudonnat M, Husson M, Besson M, Reperant C, Zemdegs J, Pages C, Hay YA, Lambolez B, Caboche J, Gutkin B, Gardier AM, Changeux JP, Faure P, Maskos U. Co-activation of VTA DA and GABA neurons mediates nicotine reinforcement. Mol Psychiatry 2013; 18: 382–93.10.1038/mp.2012.83Suche in Google Scholar
36. Tritsch NX, Oh WJ, Gu C, Sabatini BL. Midbrain dopamine neurons sustain inhibitory transmission using plasma membrane uptake of GABA, not synthesis. Elife 2014; 3: e01936.10.7554/eLife.01936Suche in Google Scholar
37. DeFelipe J. Cortical interneurons: from Cajal to 2001. Progr Brain Res 2002; 136: 215–38.10.1016/S0079-6123(02)36019-9Suche in Google Scholar
38. Wonders CP, Anderson SA. The origin and specification of cortical interneurons. Nat Rev Neurosci 2006; 7: 687–96.10.1038/nrn1954Suche in Google Scholar
39. Hensch TK. Critical period plasticity in local cortical circuits. Nat Rev Neurosci 2005; 6: 877–88.10.1038/nrn1787Suche in Google Scholar
40. Parnavelas JG. The origin and migration of cortical neurones: new vistas. Trends Neurosci 2000; 23: 126–31.10.1016/S0166-2236(00)01553-8Suche in Google Scholar
41. Cheng L, Arata A, Mizuguchi R, Qian Y, Karunaratne A, Gray PA, Arata S, Shirasawa S, Bouchard M, Luo P, Chen CL, Busslinger M, Goulding M, Onimaru H, Ma Q. Tlx3 and Tlx1 are post-mitotic selector genes determining glutamatergic over GABAergic cell fates. Nat Neurosci 2004; 7: 510–7.10.1038/nn1221Suche in Google Scholar
42. Cheng L, Samad OA, Xu Y, Mizuguchi R, Luo P, Shirasawa S, Goulding M, Ma Q. Lbx1 and Tlx3 are opposing switches in determining GABAergic versus glutamatergic transmitter phenotypes. Nat Neurosci 2005; 8: 1510–5.10.1038/nn1569Suche in Google Scholar
43. Glasgow SM, Henke RM, Macdonald RJ, Wright CV, Johnson JE. Ptf1a determines GABAergic over glutamatergic neuronal cell fate in the spinal cord dorsal horn. Development 2005; 132: 5461–9.10.1242/dev.02167Suche in Google Scholar
44. Gross MK, Dottori M, Goulding M. Lbx1 specifies somatosensory association interneurons in the dorsal spinal cord. Neuron 2002; 34: 535–49.10.1016/S0896-6273(02)00690-6Suche in Google Scholar
45. Mizuguchi R, Kriks S, Cordes R, Gossler A, Ma Q, Goulding M. Ascl1 and Gsh1/2 control inhibitory and excitatory cell fate in spinal sensory interneurons. Nat Neurosci 2006; 9: 770–8.10.1038/nn1706Suche in Google Scholar
46. Muller T, Brohmann H, Pierani A, Heppenstall PA, Lewin GR, Jessell TM, Birchmeier C. The homeodomain factor lbx1 distinguishes two major programs of neuronal differentiation in the dorsal spinal cord. Neuron 2002; 34: 551–62.10.1016/S0896-6273(02)00689-XSuche in Google Scholar
47. Bjorklund A, Dunnett SB. Dopamine neuron systems in the brain: an update. Trends Neurosci 2007; 30: 194–202.10.1016/j.tins.2007.03.006Suche in Google Scholar PubMed
48. Lennington JB, Pope S, Goodheart AE, Drozdowicz L, Daniels SB, Salamone JD, Conover JC. Midbrain dopamine neurons associated with reward processing innervate the neurogenic subventricular zone. J Neurosci 2011; 31: 13078–87.10.1523/JNEUROSCI.1197-11.2011Suche in Google Scholar PubMed PubMed Central
49. Smidt MP, Burbach JP. How to make a mesodiencephalic dopaminergic neuron. Nat Rev Neurosci 2007; 8: 21–32.10.1038/nrn2039Suche in Google Scholar PubMed
50. Damier P, Hirsch EC, Agid Y, Graybiel AM. The substantia nigra of the human brain. II. Patterns of loss of dopamine-containing neurons in Parkinson’s disease. Brain 1999; 122 (Pt 8): 1437–48.10.1093/brain/122.8.1437Suche in Google Scholar PubMed
51. Ding Y, Won L, Britt JP, Lim SA, McGehee DS, Kang UJ. Enhanced striatal cholinergic neuronal activity mediates L-DOPA-induced dyskinesia in parkinsonian mice. Proc Natl Acad Sci USA 2011; 108: 840–5.10.1073/pnas.1006511108Suche in Google Scholar
52. Schultz W. Reward signaling by dopamine neurons. Neuroscientist 2001; 7: 293–302.10.1177/107385840100700406Suche in Google Scholar
53. Seeman P, Guan HC, Van Tol HH. Dopamine D4 receptors elevated in schizophrenia. Nature 1993; 365: 441–5.10.1038/365441a0Suche in Google Scholar
54. Ungerstedt U, Ljungberg T, Steg G. Behavioral, physiological, and neurochemical changes after 6-hydroxydopamine-induced degeneration of the nigro-striatal dopamine neurons. Adv Neurol 1974; 5: 421–6.Suche in Google Scholar
55. Vasudevan A, Won C, Li S, Erdelyi F, Szabo G, Kim KS. Dopaminergic neurons modulate GABA neuron migration in the embryonic midbrain. Development 2012; 139: 3136–41.10.1242/dev.078394Suche in Google Scholar
56. Dessaud E, McMahon AP, Briscoe J. Pattern formation in the vertebrate neural tube: a sonic hedgehog morphogen-regulated transcriptional network. Development 2008; 135: 2489–503.10.1242/dev.009324Suche in Google Scholar
57. Puelles E. Genetic control of basal midbrain development. J Neurosci Res 2007; 85: 3530–4.10.1002/jnr.21363Suche in Google Scholar
58. Zervas M, Blaess S, Joyner AL. Classical embryological studies and modern genetic analysis of midbrain and cerebellum development. Curr Top Dev Biol 2005; 69: 101–38.10.1016/S0070-2153(05)69005-9Suche in Google Scholar
59. Le Dreau G, Marti E. Dorsal-ventral patterning of the neural tube: a tale of three signals. Dev Neurobiol 2012; 72: 1471–81.10.1002/dneu.22015Suche in Google Scholar PubMed
60. Kala K, Haugas M, Lillevali K, Guimera J, Wurst W, Salminen M, Partanen J. Gata2 is a tissue-specific post-mitotic selector gene for midbrain GABAergic neurons. Development 2009; 136: 253–62.10.1242/dev.029900Suche in Google Scholar PubMed
61. Nakatani T, Minaki Y, Kumai M, Ono Y. Helt determines GABAergic over glutamatergic neuronal fate by repressing Ngn genes in the developing mesencephalon. Development 2007; 134: 2783–93.10.1242/dev.02870Suche in Google Scholar PubMed
62. Lahti L, Achim K, Partanen J. Molecular regulation of GABAergic neuron differentiation and diversity in the developing midbrain. Acta Physiol (Oxf) 2013; 207: 616–27.10.1111/apha.12062Suche in Google Scholar
63. Achim K, Salminen M, Partanen J. Mechanisms regulating GABAergic neuron development. Cell Mol Life Sci 2014; 71: 1395–415.10.1007/s00018-013-1501-3Suche in Google Scholar
64. Katarova Z, Sekerkova G, Prodan S, Mugnaini E, Szabo G. Domain-restricted expression of two glutamic acid decarboxylase genes in midgestation mouse embryos. J Comp Neurol 2000; 424: 607–27.10.1002/1096-9861(20000904)424:4<607::AID-CNE4>3.0.CO;2-CSuche in Google Scholar
65. Guimera J, Vogt Weisenhorn D, Echevarria D, Martinez S, Wurst W. Molecular characterization, structure and developmental expression of Megane bHLH factor. Gene 2006; 377: 65–76.10.1016/j.gene.2006.02.026Suche in Google Scholar
66. Tsunekawa N, Yanagawa Y, Obata K. Development of GABAergic neurons from the ventricular zone in the superior colliculus of the mouse. Neurosci Res 2005; 51: 243–51.10.1016/j.neures.2004.11.011Suche in Google Scholar
67. Edwards MA, Caviness VS Jr., Schneider GE. Development of cell and fiber lamination in the mouse superior colliculus. J Comp Neurol 1986; 248: 395–409.10.1002/cne.902480308Suche in Google Scholar
68. Achim K, Peltopuro P, Lahti L, Li J, Salminen M, Partanen J. Distinct developmental origins and regulatory mechanisms for GABAergic neurons associated with dopaminergic nuclei in the ventral mesodiencephalic region. Development 2012; 139: 2360–70.10.1242/dev.076380Suche in Google Scholar
69. Marchand R, Poirier LJ. Isthmic origin of neurons of the rat substantia nigra. Neuroscience 1983; 9: 373–81.10.1016/0306-4522(83)90300-7Suche in Google Scholar
70. Miyoshi G, Bessho Y, Yamada S, Kageyama R. Identification of a novel basic helix-loop-helix gene, Heslike, and its role in GABAergic neurogenesis. J Neurosci 2004; 24: 3672–82.10.1523/JNEUROSCI.5327-03.2004Suche in Google Scholar
71. Guimera J, Weisenhorn DV, Wurst W. Megane/Heslike is required for normal GABAergic differentiation in the mouse superior colliculus. Development 2006; 133: 3847–57.10.1242/dev.02557Suche in Google Scholar
72. Peltopuro P, Kala K, Partanen J. Distinct requirements for Ascl1 in subpopulations of midbrain GABAergic neurons. Dev Biol 2010; 343: 63–70.10.1016/j.ydbio.2010.04.015Suche in Google Scholar PubMed
73. Jo AY, Park CH, Aizawa S, Lee SH. Contrasting and brain region-specific roles of neurogenin2 and mash1 in GABAergic neuron differentiation in vitro. Exp Cell Res 2007; 313: 4066–81.10.1016/j.yexcr.2007.08.026Suche in Google Scholar PubMed
74. Achim K, Peltopuro P, Lahti L, Tsai HH, Zachariah A, Astrand M, Salminen M, Rowitch D, Partanen J. The role of Tal2 and Tal1 in the differentiation of midbrain GABAergic neuron precursors. Biol Open 2013; 2: 990–7.10.1242/bio.20135041Suche in Google Scholar PubMed PubMed Central
75. Waite MR, Skidmore JM, Billi AC, Martin JF, Martin DM. GABAergic and glutamatergic identities of developing midbrain Pitx2 neurons. Dev Dyn 2011; 240: 333–46.10.1002/dvdy.22532Suche in Google Scholar PubMed PubMed Central
76. Martin DM, Skidmore JM, Philips ST, Vieira C, Gage PJ, Condie BG, Raphael Y, Martinez S, Camper SA. PITX2 is required for normal development of neurons in the mouse subthalamic nucleus and midbrain. Dev Biol 2004; 267: 93–108.10.1016/j.ydbio.2003.10.035Suche in Google Scholar PubMed
77. Hatten ME. New directions in neuronal migration. Science 2002; 297: 1660–3.10.1126/science.1074572Suche in Google Scholar PubMed
78. Corbin JG, Nery S, Fishell G. Telencephalic cells take a tangent: non-radial migration in the mammalian forebrain. Nat Neurosci 2001; 4 Suppl: 1177–82.10.1038/nn749Suche in Google Scholar PubMed
79. Anderson SA, Marin O, Horn C, Jennings K, Rubenstein JL. Distinct cortical migrations from the medial and lateral ganglionic eminences. Development 2001; 128: 353–63.10.1242/dev.128.3.353Suche in Google Scholar PubMed
80. Won C, Lin Z, Kumar TP, Li S, Ding L, Elkhal A, Szabo G, Vasudevan A. Autonomous vascular networks synchronize GABA neuron migration in the embryonic forebrain. Nat Commun 2013; 4: 2149–63.10.1038/ncomms3149Suche in Google Scholar PubMed PubMed Central
81. Tan SS, Valcanis H, Kalloniatis M, Harvey A. Cellular dispersion patterns and phenotypes in the developing mouse superior colliculus. Dev Biol 2002; 241: 117–31.10.1006/dbio.2001.0505Suche in Google Scholar PubMed
82. Kim J, Inoue K, Ishii J, Vanti WB, Voronov SV, Murchison E, Hannon G, Abeliovich A. A MicroRNA feedback circuit in midbrain dopamine neurons. Science 2007; 317: 1220–4.10.1126/science.1140481Suche in Google Scholar
83. Nunes I, Tovmasian LT, Silva RM, Burke RE, Goff SP. Pitx3 is required for development of substantia nigra dopaminergic neurons. Proc Natl Acad Sci USA 2003; 100: 4245–50.10.1073/pnas.0230529100Suche in Google Scholar
84. van den Munckhof P, Luk KC, Ste-Marie L, Montgomery J, Blanchet PJ, Sadikot AF, Drouin J. Pitx3 is required for motor activity and for survival of a subset of midbrain dopaminergic neurons. Development 2003; 130: 2535–42.10.1242/dev.00464Suche in Google Scholar
85. Smidt MP, Smits SM, Bouwmeester H, Hamers FP, van der Linden AJ, Hellemons AJ, Graw J, Burbach JP. Early developmental failure of substantia nigra dopamine neurons in mice lacking the homeodomain gene Pitx3. Development 2004; 131: 1145–55.10.1242/dev.01022Suche in Google Scholar
86. Chung S, Hedlund E, Hwang M, Kim DW, Shin BS, Hwang DY, Kang UJ, Isacson O, Kim KS. The homeodomain transcription factor Pitx3 facilitates differentiation of mouse embryonic stem cells into AHD2-expressing dopaminergic neurons. Mol Cell Neurosci 2005; 28: 241–52.10.1016/j.mcn.2004.09.008Suche in Google Scholar
87. Langston JW, Langston EB, Irwin I. MPTP-induced parkinsonism in human and non-human primates – clinical and experimental aspects. Acta neurologica Scandinavica Supplementum 1984; 100: 49–54.Suche in Google Scholar
88. Flores C, Manitt C, Rodaros D, Thompson KM, Rajabi H, Luk KC, Tritsch NX, Sadikot AF, Stewart J, Kennedy TE. Netrin receptor deficient mice exhibit functional reorganization of dopaminergic systems and do not sensitize to amphetamine. Mol Psychiatry 2005; 10: 606–12.10.1038/sj.mp.4001607Suche in Google Scholar
89. Fazeli A, Dickinson SL, Hermiston ML, Tighe RV, Steen RG, Small CG, Stoeckli ET, Keino-Masu K, Masu M, Rayburn H, Simons J, Bronson RT, Gordon JI, Tessier-Lavigne M, Weinberg RA. Phenotype of mice lacking functional deleted in colorectal cancer (Dcc) gene. Nature 1997; 386: 796–804.10.1038/386796a0Suche in Google Scholar
90. Nishikawa S, Goto S, Yamada K, Hamasaki T, Ushio Y. Lack of Reelin causes malpositioning of nigral dopaminergic neurons: evidence from comparison of normal and Reln(rl) mutant mice. J Comp Neurol 2003; 461: 166–73.10.1002/cne.10610Suche in Google Scholar
91. Ohyama K, Kawano H, Asou H, Fukuda T, Oohira A, Uyemura K, Kawamura K. Coordinate expression of L1 and 6B4 proteoglycan/phosphacan is correlated with the migration of mesencephalic dopaminergic neurons in mice. Brain Res Dev Brain Res 1998; 107: 219–26.10.1016/S0165-3806(97)00220-4Suche in Google Scholar
92. Demyanenko GP, Shibata Y, Maness PF. Altered distribution of dopaminergic neurons in the brain of L1 null mice. Brain Res Dev Brain Res 2001; 126: 21–30.10.1016/S0165-3806(00)00129-2Suche in Google Scholar
93. Shults CW, Kimber TA. Mesencephalic dopaminergic cells exhibit increased density of neural cell adhesion molecule and polysialic acid during development. Brain Res Dev Brain Res 1992; 65: 161–72.10.1016/0165-3806(92)90175-VSuche in Google Scholar
94. Kania A, Johnson RL, Jessell TM. Coordinate roles for LIM homeobox genes in directing the dorsoventral trajectory of motor axons in the vertebrate limb. Cell 2000; 102: 161–73.10.1016/S0092-8674(00)00022-2Suche in Google Scholar
95. Sharma K, Sheng HZ, Lettieri K, Li H, Karavanov A, Potter S, Westphal H, Pfaff SL. LIM homeodomain factors Lhx3 and Lhx4 assign subtype identities for motor neurons. Cell 1998; 95: 817–28.10.1016/S0092-8674(00)81704-3Suche in Google Scholar
96. Sharma K, Leonard AE, Lettieri K, Pfaff SL. Genetic and epigenetic mechanisms contribute to motor neuron pathfinding. Nature 2000; 406: 515–9.10.1038/35020078Suche in Google Scholar PubMed
97. De Marco Garcia NV, Jessell TM. Early motor neuron pool identity and muscle nerve trajectory defined by postmitotic restrictions in Nkx6.1 activity. Neuron 2008; 57: 217–31.Suche in Google Scholar
98. Muller M, Jabs N, Lorke DE, Fritzsch B, Sander M. Nkx6.1 controls migration and axon pathfinding of cranial branchio-motoneurons. Development 2003; 130: 5815–26.10.1242/dev.00815Suche in Google Scholar PubMed
99. Shirasaki R, Pfaff SL. Transcriptional codes and the control of neuronal identity. Annu Rev Neurosci 2002; 25: 251–81.10.1146/annurev.neuro.25.112701.142916Suche in Google Scholar PubMed
100. Nobrega-Pereira S, Kessaris N, Du T, Kimura S, Anderson SA, Marin O. Postmitotic Nkx2–1 controls the migration of telencephalic interneurons by direct repression of guidance receptors. Neuron 2008; 59: 733–45.10.1016/j.neuron.2008.07.024Suche in Google Scholar PubMed PubMed Central
101. Hussman JP. Suppressed GABAergic inhibition as a common factor in suspected etiologies of autism. J Autism Dev Disord 2001; 31: 247–8.10.1023/A:1010715619091Suche in Google Scholar
102. Levitt P, Eagleson KL, Powell EM. Regulation of neocortical interneuron development and the implications for neurodevelopmental disorders. Trends Neurosci 2004; 27: 400–6.10.1016/j.tins.2004.05.008Suche in Google Scholar PubMed
103. Rubenstein JL, Merzenich MM. Model of autism: increased ratio of excitation/inhibition in key neural systems. Genes Brain Behav 2003; 2: 255–67.10.1034/j.1601-183X.2003.00037.xSuche in Google Scholar PubMed PubMed Central
104. Lewis DA. GABAergic local circuit neurons and prefrontal cortical dysfunction in schizophrenia. Brain Res Brain Res Rev 2000; 31: 270–6.10.1016/S0165-0173(99)00042-9Suche in Google Scholar
105. Benes FM, Berretta S. GABAergic interneurons: implications for understanding schizophrenia and bipolar disorder. Neuropsychopharmacology 2001; 25: 1–27.10.1016/S0893-133X(01)00225-1Suche in Google Scholar
106. Jetty PV, Charney DS, Goddard AW. Neurobiology of generalized anxiety disorder. Psychiatr Clin North Am 2001; 24: 75–97.10.1016/S0193-953X(05)70207-0Suche in Google Scholar
107. Stefansson H, Sigurdsson E, Steinthorsdottir V, Bjornsdottir S, Sigmundsson T, Ghosh S, Brynjolfsson J, Gunnarsdottir S, Ivarsson O, Chou TT, Hjaltason O, Birgisdottir B, Jonsson H, Gudnadottir VG, Gudmundsdottir E, Bjornsson A, Ingvarsson B, Ingason A, Sigfusson S, Hardardottir H, Harvey RP, Lai D, Zhou M, Brunner D, Mutel V, Gonzalo A, Lemke G, Sainz J, Johannesson G, Andresson T, Gudbjartsson D, Manolescu A, Frigge ML, Gurney ME, Kong A, Gulcher JR, Petursson H, Stefansson K. Neuregulin 1 and susceptibility to schizophrenia. Am J Hum Genet 2002; 71: 877–92.10.1086/342734Suche in Google Scholar PubMed PubMed Central
108. Kim TG, Yao R, Monnell T, Cho JH, Vasudevan A, Koh A, Peeyush KT, Moon M, Datta D, Bolshakov VY, Kim KS, Chung S. Efficient specification of interneurons from human pluripotent stem cells by dorsoventral and rostrocaudal modulation. Stem Cells 2014; 32: 1789–804.10.1002/stem.1704Suche in Google Scholar PubMed PubMed Central
109. Maroof AM, Keros S, Tyson JA, Ying SW, Ganat YM, Merkle FT, Liu B, Goulburn A, Stanley EG, Elefanty AG, Widmer HR, Eggan K, Goldstein PA, Anderson SA, Studer L. Directed differentiation and functional maturation of cortical interneurons from human embryonic stem cells. Cell Stem Cell 2013; 12: 559–72.10.1016/j.stem.2013.04.008Suche in Google Scholar PubMed PubMed Central
110. Maroof AM, Brown K, Shi SH, Studer L, Anderson SA. Prospective isolation of cortical interneuron precursors from mouse embryonic stem cells. J Neurosci 2010; 30: 4667–75.10.1523/JNEUROSCI.4255-09.2010Suche in Google Scholar PubMed PubMed Central
111. Maisano X, Litvina E, Tagliatela S, Aaron GB, Grabel LB, Naegele JR. Differentiation and functional incorporation of embryonic stem cell-derived GABAergic interneurons in the dentate gyrus of mice with temporal lobe epilepsy. J Neurosci 2012; 32: 46–61.10.1523/JNEUROSCI.2683-11.2012Suche in Google Scholar PubMed PubMed Central
112. Liu Y, Liu H, Sauvey C, Yao L, Zarnowska ED, Zhang SC. Directed differentiation of forebrain GABA interneurons from human pluripotent stem cells. Nat Protoc 2013; 8: 1670–9.10.1038/nprot.2013.106Suche in Google Scholar PubMed PubMed Central
113. Kim JH, Auerbach JM, Rodriguez-Gomez JA, Velasco I, Gavin D, Lumelsky N, Lee SH, Nguyen J, Sanchez-Pernaute R, Bankiewicz K, McKay R. Dopamine neurons derived from embryonic stem cells function in an animal model of Parkinson’s disease. Nature 2002; 418: 50–6.10.1038/nature00900Suche in Google Scholar PubMed
114. Kawasaki H, Mizuseki K, Nishikawa S, Kaneko S, Kuwana Y, Nakanishi S, Nishikawa SI, Sasai Y. Induction of midbrain dopaminergic neurons from ES cells by stromal cell-derived inducing activity. Neuron 2000; 28: 31–40.10.1016/S0896-6273(00)00083-0Suche in Google Scholar
115. Bjorklund LM, Sanchez-Pernaute R, Chung S, Andersson T, Chen IY, McNaught KS, Brownell AL, Jenkins BG, Wahlestedt C, Kim KS, Isacson O. Embryonic stem cells develop into functional dopaminergic neurons after transplantation in a Parkinson rat model. Proc Natl Acad Sci USA 2002; 99: 2344–9.10.1073/pnas.022438099Suche in Google Scholar PubMed PubMed Central
116. Yang D, Zhang ZJ, Oldenburg M, Ayala M, Zhang SC. Human embryonic stem cell-derived dopaminergic neurons reverse functional deficit in parkinsonian rats. Stem Cells 2008; 26: 55–63.10.1634/stemcells.2007-0494Suche in Google Scholar PubMed PubMed Central
©2014 by De Gruyter
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Artikel in diesem Heft
- Frontmatter
- Reviews
- Telocytes revisited
- Mesencephalic GABA neuronal development: no more on the other side of oblivion
- All motors have to decide is what to do with the DNA that is given them
- DNA-bound metal ions: recent developments
- Regulation of LINE-1 in mammals
- Glycolipozyme membrane protein integrase (MPIase): recent data
Artikel in diesem Heft
- Frontmatter
- Reviews
- Telocytes revisited
- Mesencephalic GABA neuronal development: no more on the other side of oblivion
- All motors have to decide is what to do with the DNA that is given them
- DNA-bound metal ions: recent developments
- Regulation of LINE-1 in mammals
- Glycolipozyme membrane protein integrase (MPIase): recent data