
Tachykinin: recent developments and novel roles in health and disease
-
Takenori Onaga
Takenori Onaga received his Bachelor’s degree of Doctor of Veterinary Medicine from the Rakuno Gakuen University (Ebetsu, Japan) in 1990. He started his research on bioactive peptides in the laboratory of veterinary physiology in the same university and, in this way, received research training with Dr. Satoru Naruse and Dr. Atsukazu Kuwahara in the Institute of Physiological Science (Okazaki, Japan). He earned his PhD in Veterinary Medicine from the Hokkaido University (Sapporo, Japan) under the supervision of Dr. Yoshiaki Habara in 2001 and continued his research in the aforementioned laboratory. He is a Professor in the present Laboratory since 2009.


Abstract
Over 80 years has passed since the discovery of substance P (SP), and a variety of peptides of the tachykinin (TK) family have been found and investigated. SP, neurokinin A (NKA), and neurokinin B (NKB) are representative peptides in mammalian species. SP and NKA are major excitatory neurotransmitters in the peripheral nervous system, while NKB is primarily involved in the central nervous system (CNS). Moreover, TK peptides play roles not only as neurotransmitters but also as local factors and are involved in almost all aspects of the regulation of physiological functions and pathophysiological processes. The role of SP as a mediator of pain processing and inflammation in peripheral tissues in coordination with transient receptor potential channels is well established, while novel aspects of TKs in relation to hematopoiesis, venous thromboembolism, tendinopathy, and taste perception have been clarified. In the CNS, the NKB signaling system in the hypothalamus has been shown to play a crucial role in the regulation of gonadotropin hormone secretion and the onset of puberty, and molecular biological studies have elucidated novel prophylaxic activities of TKs against neurogenic movement disorders based on their molecular structure. This review provides an overview of the novel aspects of TKs reported around the world in the last 5 years, with particular focus on nociception, inflammation, hemopoiesis, gonadotropin secretion, and CNS diseases.
List of abbreviations: ARC, arculate nucleus; CNS, central nervous system; DRG, dorsal root ganglion; Dyn, dynorphin; EK, endokinin; GCPR, G protein-coupled receptors; GRP, gastrin-releasing peptide; HH, hypogonadtropic hypogonadism; HK-1, hemokinin-1; IL, interleukin; ICV, intracerebroventricular; IV, intravenous; KOR, κ opioid receptor; NK, neurokinin; NKA, neurokinin A; NKB, neurokinin B; NKR, neurokinin receptor; NP, neuropeptide; NMDA, N-methyl-d-asparate; NST, nucleus of solitary tract; PNS, peripheral nervous system; POA, preoptic area; PPT, preprotachykinin; PANs, primary afferent nerves; TK, tachykinin; TRP, transient receptor potential; TNFα, tumor necrosis factor α; VEGF, vascular endothelial growth factor.
Introduction
Tachykinins (TKs) are a family of neuropeptides widely distributed throughout the animal body. The three major TKs in mammalian species are substance P (SP), neurokinin A (NKA), and neurokinin B (1–3) (NKB), while other types of TK peptides have also been identified, i.e., neuropeptide K (NPK) (4), neuropeptide γ (NPγ) (5), hemokinin-1 (HK-1), and endokinin (EKs) (6) (Table 1). SP, NKA, NPK, and NPγ are encoded in the Tac1 (pre-pro-tachykinin A, Ppt-a) gene. NPK and NPγ are forms of NKA with N-terminal extensions of 26 and 11 amino acids residues, respectively (3, 5). NKB and neuropeptide F (NKF) are encoded in the Tac3 (pre-pro-tachykinin B, Ppt-b, or Tac2 in rodents) gene (7, 8), while HK-1 and EKs are encoded by the Tac4 (pre-pro-tachykinin C, Ppt-c) gene (6, 9, 10). Tac4 encodes HK-1 in the mouse and rat, whereas Tac4 is spliced into four variants (α to δ) in humans to produce EK-A to EK-D, respectively (10). Although human EK-A and EK-B are amino (N)-terminal elongated forms of hHK-1 (6), the amino acid sequence of hHK-1, except for the N-terminal six residues, differs from that of mouse and rat HK-1.
Tachykinin family peptides and gene, high-affinity receptor, and amino acid sequence.
| Tachykinin | Gene | High-affinity receptor | Amino acid sequence | References |
|---|---|---|---|---|
| Substance P (SP) | Tac1 (Ppt-a)-α, β, γ, δ | NK1R (TAC1R) | RPKPQQFFGLM-NH2 | (1) |
| Neurokinin A (NKA) | Tac1 (Ppt-a)-β, γ | NK2R (TAC2R) | HKTDSFVGLM-NH2 | (2) |
| Neuropeptide K (NPK) | Tac1 (Ppt-a)-β | NK2R (TAC2R) | DADSSIEKQVALLKALYGHGQISHKRHKTDSFVGLM-NH2 | (3) |
| Neuropeptide γ (NPγ) | Tac1 (Ppt-a)-γ | NK2R (TAC2R) | DA - - - - - - - - - - - - - - - - GHGQISHKRHKTDSFVGLM-NH2 | (2) |
| Neurokinin B (NKB) | Tac3 (Ppt-b) | NK3R (TAC3R) | DMHDFFVGLM-NH2 | (3) |
| Neurokinin F (NKF) | Tac3 (Ppt-b) | NK3R (TAC3R) | YNDIDYDSFVGLM-NH2 | (7) |
| YDDIDYDSFVGLM-NH2 | (7) | |||
| Hemokinin-1 (HK-1) | Tac4 (Ppt-c) | NK1R (TAC1R) | (mouse/rat) SRTRQFYGLM-NH2 | (5) |
| (human) TGKASQFFGLM-NH2 | (8) | |||
| Endokinin (EK)-A | Tac4 (Ppt-c) | DGGEEQTLSTEAETWVIVALEEGAGPSIQLQLQEVKTGKASQFFGLM-NH2 | (5) | |
| EK-B | Tac4 (Ppt-c) | DGGEEQTLSTEAETW - - - - - - EGA - - - - QLQLQEVKTGKASQFFGLM-NH2 | (5) |
Broken lines incidate lacking amino acid residues. Ppt-a, preprotachykinin A; Ppt-b, preprotachykinin B; Ppt-c, preprotachykinin C.
A characteristic structure common to TKs is a five-amino acid sequence at the amidated carboxyl(C)-terminus (4), i.e., FxGLM-NH2, whereas the fourth amino acid (x; a hydrophobic residue) varies in different peptides (11) (Table 1). The three major TKs can bind to neurokinin receptor (NKR) type 1 (NK1R, TACR1), NKR type 2 (NK2R, TACR2), and NKR type 3 (NK3R, TACR3), which are classified in the rhodopsin family of G protein-coupled receptors (GPRs) (12). SP, NKA, and NKB have higher affinity to NK1R, NK2R, and NK3R, respectively (13). HK-1 is an NK1R agonist with equipotent activity to SP in terms of in vivo cardiovascular effects (14) and the immune system (15). As NPK and NPγ have C-terminal structure identical to that of NKA, they also bind to NK2R, and NPγ selectively binds to NK2R more potently than does NKA (16). The C-terminus of NPK, NKγ, and NKA forms an α-helix structure and binds to the transmembrane region of the receptor, while the N-terminus of NPK appears to interact with an extracellular loop of the receptor for stabilization of the C-terminus (17); that is, the C-terminus is the ‘message’ domain concerned with receptor activation, while the N-terminus plays an ‘address’ function in determining receptor subtype specificity (18).
In the past three decades, TKs have been actively investigated as neurotransmitters in the central (CNS) and peripheral nervous system (PNS), and considerable knowledge on neural TKs has been accumulated, with a detailed review on a variety of aspects published (19). Therefore, this review addresses recent developments in TK and NKR biology over the last 5 years, with particular focus on nociception, inflammation, hemopoiesis, gonadotropin secretion, and neurogenic diseases associated with the CNS.
TKs and peripheral afferent nerves
Roles of TKs in nociception
Neural TKs are released from nonmyelinated C-fibers that consist of primary afferent nerves (PANs) of the sensory nervous route of the systemic and autonomic nervous systems (20) (Figure 1). PANs are very sensitive to capsaicin, a ligand of the transient receptor potential (TRP) vanilloid-1 (TRPV1) (21), previously called the vanilloid receptor-1. Among the TRPs, TRPV1, TRP vanilloid-4, and TRP ankyrin-1 (TRPA1) play crucial roles as polymodal nociceptors-sensing chemical, mechanical, and thermal stimuli (21, 22). The neurons involving TKs are related to nociception and axon reflex, and they primarily express TRPV1. Low-dose capsaicin treatment in rats was shown to increase mRNA expression of TRPV1, SP, and NK1R in the sensory nerves in the spinal cord (23). In a rat model of gout, monosodium urate caused nociception and edema, and the responses were abolished by TRPV1 and NK1R antagonists (24). In studies of local inflammation in mice, TRPV1 knockout was shown to increase cytokine release and apoptosis of mononuclear cells in response to endotoxin lipopolysaccharide (LPS), resulting in impaired macrophage-associated defense mechanisms, with a particular decrease in NK1R-dependent phagocytosis by macrophages (25). These observations indicate that the TRPV1 and SP-NK1R axis acts as a defense against damage from sepsis. SP- and NK1R-containing PANs were shown to be involved to some extent in hypersensitivity to surgical pain in rats (26). PANs in the rat spinal cord co-express μ- and δ-opioid receptors, and agonists of both opioid receptors potently inhibited capsaicin-induced SP release (27). These μ- and δ-opioid receptors in the spinal cord serve as crucial action points for pain control by analgesics.
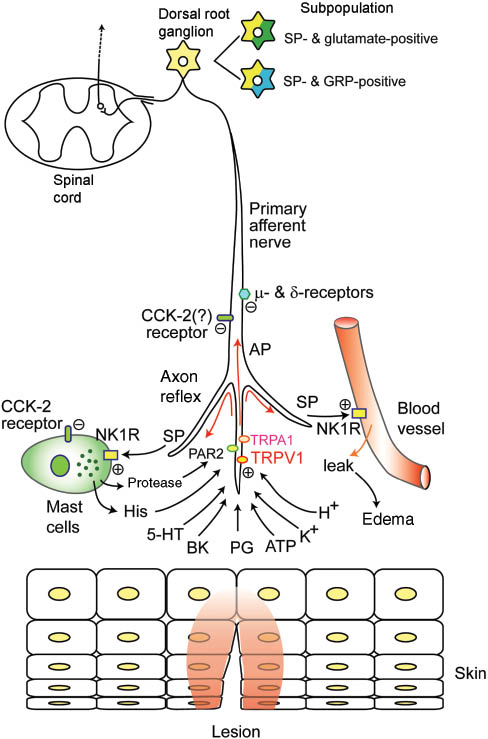
Substance P (SP)-containing primary afferent nerves and nociception.
The illustration shows peripheral tissue lesions and nociception. Irritant stimulus activates TRPA1 and TRPV1, which generates action potential (AP) in primary afferent nerves (PANs) that contain SP and carcitonin gene-related peptides (CGRP). AP causes an axon reflex in PANs releasing SP and CGRP. SP triggers degranulation of mast cells releasing histamine and protease and increasing vascular permeability through NK1R. Activation of both μ- and δ-receptors attenuate peripheral pain, and activation of CCK receptors attenuate itch and degranulation of mast cells. TRP, transient receptor potential; TRPA1, TRP ankiin-2; TRPV1, TRP vanilloid-1; NK1R, neurokinin receptor type-1; PAR2, protease-activated receptor type-2.
In the CNS, the rostral ventromedial medulla (RVM), a central relay in the bulbospinal pathways that modulates nociception, contains high concentrations of SP and NK1R. Microinjection of SP at picomole levels in the RVM produced a concentration-dependent increase in the number of discharges in distal dendrites of RVM neurons that were immunoreactive for NK1R (28). This effect was attenuated by an NK1R antagonist, suggesting the involvement of the SP-NK1R axis in the afferent pathway in the CNS.
TKs and itch
Intradermal injection of SP induces itch and scratching behavior through stimulation of the C-fibers via chemical mediators such as leukotriene B4 (29). Recently, SP/glutamate or SP/gastrin-releasing peptide (GRP) co-expressing neurons have been shown to be the focus of the spinal neurotransmission of histamine-dependent and -independent itch (30) (Figure 1). A majority of GRP immunopositive fibers in dorsal root ganglion (DRG) cells express SP, CGRP, and TRPV1 (31, 32). A recent in vivo study demonstrated that SP, glutamate, and GRP each partially contributes to histamine-independent itch (33). Patch clamp recordings using slices of spinal cord from rats and mice revealed that a portion of dorsal horn neurons receiving input from PANs showed an increased firing frequency of action potential in response to GRP application (34). However, the population of GRP-sensitive neurons was as high as 10% DRG cells, and neurotransmission between the neurons was shown to be primarily mediated by glutamate, not by GRP (34). Subpopulations of chloroquine- and/or histamine-sensitive DRG neurons were immunopositive for SP and/or GRP, and more than 80% of them were immunopositive for vesicular glutamate transporter type 2 (35). Co-administration of NK1R and α-amino-3-hydroxy-5-methyl-4-isoxazole proprionate acid (AMPA)-glutamate receptor antagonists was more effective in abolishing scratching behavior in response to histamine, indicating that histamine-evoked itch is mediated primarily by SP and glutamate, with GRP playing a lesser role. Accordingly, the co-application of NK1R and AMPA receptor antagonists may prove more beneficial than the application of GRP receptor antagonists in treating chronic itch.
Interestingly, topical application of a duodenal peptide cholecystokinin (CCK) to the back skin reduced scratching frequency of an SP-induced response in mice in vivo and degranulation of skin-derived mast cells via CCK receptors type 2 in an in vitro study (36) (Figure 1). Visceral C-fibers, particularly vagal afferent fibers, are sensitive not only to TRPV1 agonist capsaicin but also to CCK, inducing satiety and inhibiting gastric emptying (37), whereas it remains unknown whether epidermal C-fibers are sensitive to CCK and attenuate SP-induced itch behavior. The aforementioned report suggests that epidermal C-fibers also express CCK receptors. This novel effect of CCK may help in the suppression of the itch and the development of a novel antipruritic drug.
TKs and inflammation
Neurogenic inflammation, characterized by vasodilatation and increased capillary permeability, is induced both in the mucosa and skin through the axon reflex in PANs expressing TKs and TRPV1 (38). Released SP activates NF-κB in leukocytes and the release of proinflammatory cytokines (39). Desensitization to SP-evoked current was exhibited in almost all SP-responsive DRG neurons in control rats, while desensitization diminished in a larger proportion (two thirds) of DRG neurons in rats with inflammation (40). In addition, pretreatment with SP potentiated the N-methyl-d-asparate (NMDA)-evoked cytosolic calcium transient response in DRG PANs, and this response was blocked by an NK1R-antagonist (41). The reports suggest that the SP-NK1R axis not only activates postsynaptic dorsal horn neurons but also contributes to DRG sensitization in DRG PANs; that is, they develop and maintain inflammatory pain.
A novel gas-transmitter, hydrogen sulfide (H2 S) was shown to exert cardiovascular effects in septic shock (42). Recently, it was also demonstrated that H2 S regulates TRPV1-mediated neurogenic inflammation in polymicrobial sepsis triggered through the enhancement of SP production and activation of the ERK-NF-κB pathway (43), indicating that the TRPV1 and SP-NK1R axes are involved in systemic inflammatory response and multiple organ dysfunction in cases of sepsis. H2 S donor treatment to the isolated mouse pancreas caused inflammation and increased expression of the Tac1 and NK1R genes in the tissue (44). This observation supports the notion that the SP-NK1R axis has a crucial role as a primary mediator of H2 S-induced deleterious responses.
TRPA1, a major mediator of inflammatory pain and mechanical hyperalgesia (22), is activated by exogenous alimentary and environmental irritants, which cause the release of SP and CGRP from the PANs (45). Further, proteases released from the surrounding tissue, particularly from mast cells stimulated by the axon reflex, are potent mediators of inflammation and pain and activate protease-activated receptor 2 (PAR2), which in turn sensitizes TRPV1 and stimulates SP secretion in the PANs (46, 47) (Figure 1). This feedback loop further amplifies inflammatory pain.
A recent report described an opposite effect, with TRPV1 knockout causing impaired macrophage-associated immunoresponses in a mouse sepsis model, induced by cecal ligation and puncture, in which NK1-dependent phagocytosis and bacterial clearance decreased and plasma cytokines increased (25). This report indicates that TRPV1 has a protective effect against local inflammation in sepsis, implying the existence of a dual role for TRPV1 in inflammation.
Role of TKs in inflammation in the skin
The chemical sensors TRPV1 and TRPA1 are abundantly distributed in PANs containing TKs in the skin, and the SP-NK1R axis is involved in chronic dermal inflammatory diseases (48). Indeed, SP and HK-1, but not NKA and NKB, were shown to switch CD4+ memory T cells into Th17 cells that may act locally on memory T cells in cooperation with proinflammatory cytokines such as interleukin(IL)-1β secreted from monocytes, resulting in amplification of inflammatory responses (49). Further, TKs stimulate secretion of proinflammatory cytokines from keratinocytes through NK2R activation (50), which further enhances inflammation.
Inflammation itself increases the peptide content of DRG PANs (51). Interestingly, exposure of mice to sonic stress for 24 h increased SP+ and CGRP+ neurons in the DRG innervating the skin, indicating neural plasticity in response to stress (52). In mice with atopic eczema, the number of SP nerves decreased in those chronically stressed with atopic-like eczema, whereas the mRNA expression of NK1R tended to increase, together with the number and degranulation of mast cell, suggesting that chronic stress changes sensitivity and SP signaling system outcomes in the skin (53).
Roles of TKs in chemically induced inflammatory colitis
Colonic cancer results in high mortality in Western countries, and many studies have revealed that chronic inflammation of the colon, such as irritable bowel syndrome, is associated with colonic cancer. However, the detailed mechanisms underlying inflammation-derived colonic cancer remains obscure. Although bacterial products and toll-like receptors (TLR) are probably involved in colonic inflammation, several lines of study have indicated that intramural primary afferent neurons (IPANs) expressing TKs are involved in the neuroimmuno-interaction of inflammation between nociceptive receptors and cytokines.
Trinitrobenzene-sulfonic-acid (TNBS), dextran-sulfate-sodium-salt (DSS), and oxazolone are representative chemical tools for the induction of inflammatory colitis. In TNBS-induced inflammatory colitis in rats, the myenteric expression of pan-neural marker protein gene product 9.5 significantly increased in association with elevation in SP, NPK, and galanin immunoreactivity (54). In TNBS-induced colitis in mice, a similar TNBS-induced, TRPA1-dependent and sustained depolarization and release of SP and CGRP in colonic sensory neurons was observed (55). Both TNBS- and DSS-induced colitis was inhibited or reduced in TRPA1 knockout mice as well as by the application of a pharmacological inhibitor of TRPA1. Further, TNBS-induced colitis increased the mechanical hypersensitivity of the mice, whereas the knockout of the Trpa1 gene abolished the response (22). Taken together, TRPA1 can be seen to trigger TK secretion in the visceral sensory neurons, which subsequently exaggerates inflammatory colitis and hyperalgesia. Thus, TRPA1 agonists are a good candidate for the therapeutic treatment of colitis.
In mice, capsaicin- and acid-induced SP and CGRP release from the inflamed distal colon in DSS-induced colitis was significantly higher than that from the noninflamed colon (56). During DSS-induced colitis, the proportions of TRPV1 expressed in DRGs neurons and the relative content of TRPV1 mRNA were significantly increased in DRGs projecting to the inflamed distal colon compared to those receiving vehicle treatment. Systemic pretreatment with resiniferatoxin (an ultrapotent TRPV1 agonist) desensitizes the entire capsaicin-sensitive population of sensory neurons. DSS-induced colitis was ameliorated by resiniferatoxin treatment and in SP-/- knockout mice but not in wild-type or CGRP-/- knockout mice, suggesting that the responsiveness of TRPV1- and SP-expressing neurons to inflammatory factors determines the severity of DSS-induced colitis in mice. Although SP and CGRP were released from identical IPANs in the gastrointestinal tract, they exerted opposite actions in mice with oxazolone-induced colitis; that is, SP-/- knockout protected against colitis, whereas CGRP-/- knockout increased susceptibility to colitis compared to that in wild-type mice (57). SP-/- knockout simultaneously decreased inflammatory cytokine production, including IL-4, IL-5, and IL-11, by monocytes in the colonic lamina propria, indicating neuro-immuno interactions via SP and the involvement of these cytokines. Thus, the augmented release of SP increases cytokine production and leads to further inflammation. TRPV1-triggered SP release was shown to be more abundant in the distal colon and rectum than in the proximal colon in mice and rats (57, 58). The density gradient coincided with severity of inflammation in the animals with DSS-induced colitis. This observation strongly suggests that a neural mediator gradient underlies the distribution of sites susceptible to inflammation and, presumably, to intestinal cancers.
Chronic inflammatory colitis is known to be a factor in the induction of colonic cancer. An immunohistochemical study on human chronic ulcerative colitis revealed that the total density of NK1R increased by 40% and 80% in cases of high-grade dysplasia and carcinoma, respectively, whereas mRNA expression of the truncated form of NK1R differed more than tenfold between the high-grade dysplasia and carcinoma groups (59). These data suggest that not only overexpression of NK1R but also structural abnormalities in NK1R may contribute to the deterioration of chronic colitis to colonic cancer. The C-terminal-truncated form of the receptor was shown to lack phosphorylation sites interacting with β-arrestin, a mediator of NKR trafficking in the cells, so they appear to be resistant to desensitization (19). In addition, as the truncated form does not couple to intracellular messenger, such as Ca2+ and NK-κB (60, 61), they appear to send different signals to the cells.
Roles of TKs in gastritis and gastric cancers
The NK1R antagonist was shown to exert antitumor action in human gastric adenocarcinoma cell lines (62). This report coincides with the observation that SP application accelerated proliferation and migration of the human poorly differentiated adenocarcinoma MKN45 cell line in vitro (63). Therefore, SP neurons appear to be involved not only in the carcinogenic process but also in the proliferation and invasion of gastric carcinoma cells. It is worth noting that the density of SP-expressing nerves increased in inverse proportion to the differentiation of tumor cells in human gastric cancer (63), indicating an undefined inverse relationship; that is, a high density of SP-positive nerves may inhibit tumor differentiation. Such a protective effect of PANs has been demonstrated in human chronic gastritis by using low doses of capsaicin (64). In patients with gastritis, the neuronal SP level decreased in normally appearing mucosa, whereas nonneuronal SP increased in the diseased area in gastritis and ulcer patients (65), with Helicobactor pylori infection further decreasing neuronal SP levels. The decrease in SP, a gastroprotective factor, appears to predispose patients to mucosal cellular damage in cases of gastritis. In the CNS, an intracerebroventricular (ICV) injection of SP inhibited the formation of ethanol-induced gastric ulcers, although intravenously (IV) injected SP had no effect (66). The protective effect of central SP was inhibited by pretreatment with antagonists of the three NKR subtypes, as well as μ-opioid receptor and endomorphin-2 antagonists. In addition, centrally injected SP reversed the ethanol-induced reduction of gastric mucosal CGRP content. Hence, SP may induce centrally initiated gastric mucosal protection through the peripheral action of CGRP.
Role of TKs in hypersensitivity and inflammation in airway
Neurogenic inflammation in the airway has been reviewed (67, 68) and, as regards asthma, it was demonstrated that SP and NKA released from airway sensory nerves cause bronchoconstriction through NK2R in humans (69) and plasma exudation through NK1R in guinea pigs (70). Elevation of tracheal temperature by humidified warm air induced bronchospasms in ovalbumin-sensitized rats, and this response was abolished by NK1R and NK2R antagonists (71). In another study, it was shown that hypochlorite-ovalbumin-induced hypersensitivity was not induced in TRPA1 knockout mice (72). These reports imply that TRPA1 and bronchopulmonary vagal C-fibers releasing SP and NKA are key mediators of airway hypersensitivity in asthma. Moreover, nasal antigen exposure rapidly induced bronchial hyper-responsiveness through SP release and binding to NK1R without direct stimulation of the bronchial mucosa by antigens in mice (73), implying that united but remote interactions can be triggered between the upper and lower airways through TK release, presumably mediated by the vagal C-fibers.
TKs have been shown to be involved in the regulation of the immunological process of inflammation in the airways and immuno-mediating cells, such as lymphocytes and mast cells, express NKRs (74). SP was shown to be a potent chemoattractant for basophils through NK1R activation in a human in vitro study (75). With regard to environmental pollution and its biological effect on the airway, diesel exhaust particles were shown to induce edema in the rat trachea and bronchus via the up-regulation of TRPV1, NK1R, and NK2R (76), implying that treatment with receptor antagonists can ameliorate the symptoms caused by air pollutants such as particulate matter <2.5 μm.
Role of TKs in inflammation in the brain
SP is abundant not only in peripheral PANs but also in the CNS, where it was shown to regulate inflammation (77). NK1R is predominantly distributed in glial cells and astrocytes, and inflammatory cytokines released from these cells after bacterial exposure were shown to be inhibited by an NK1R receptor antagonist both in vivo and in vitro (78). Moreover, in response to streptococcal infection in the CNS of mice in vivo, a nonlethal dose of bacteria increased the permeability of the blood-brain barrier (BBB) in wild-type mice, while no such response was induced in NK1R knockout mice (79). Pretreatment with an NK1R antagonist abolished the BBB response and attenuated the release of inflammatory cytokines such as TNF-α and IL-6, as well as further augmented the release of the immunosuppressive cytokine IL-10. Thus, NK1R is definitely involved in the modulation of BBB permeability in sepsis, suggesting the possibility of the prophylactic use of NK1R inhibition for encephalitis induced by bacterial infection.
Acute brain injury, such as neurotrauma or stroke, increases intracranial pressure (ICP), which causes perivascular accumulation of SP in association with increased BBB permeability and formation of vasogenic edema. Administration of an NK1R antagonist was found to reduce potently the increase in BBB permeability and edema formation (80). Therefore, NK1R antagonists seem to be an effective candidate for a novel therapeutic approach to the treatment of acute brain injury. A secondary increase in ICP within 2 h of subarachnoid hemorrhage (SAH) was followed by brain edema. In a rat model of SAH, the administration of the NK1R antagonist n-acetyl-tryptophan (NAT) after 30 min did not improve any outcome parameters. However, NAT administration at 4 h after ischemic stroke in rat was effective in reducing BBB permeability and cerebral edema (81), suggesting that the SP-NK1R axis makes a crucial contribution to cerebral edema after ischemic stroke, presumably in association with temporal changes in its degree of contribution.
NK1R antagonists have the potential to promote recovery following traumatic brain injury (TBI) (82). Infusion of SP promoted cellular proliferation in the subventricular zone and dentate gyrus following TBI in rats, which was largely associated with microglial proliferation but did not correspond with any functional improvements (83). As NAT treatment reduces microglial proliferation and improves motor outcome, NAT appears to be an effective neuroprotector following TBI. Although the neuroprotective effect of NAT was limited to 5 h after TBI, administration of a highly lipid soluble NK1R antagonist L-732,138 provided effective neuroprotection even after a delay of 12 h in rats (84). This suggests that SP is involved in the pathogenesis of TBI and that NK1R antagonist treatment may ameliorate TBI, although the lipid solubility of chemicals greatly influences the efficacy of the antagonists.
Craniotomy and associated injury to the somatosensory cortex was shown to cause transient periorbital allodynia and elevate nociceptive neuropeptides, such as SP, CGRP, and glial fibrillary acidic protein, in the brainstem of mice without change in macrophage/microglial cells (85). Therefore, the mechanism underlying persistent allodynia does not appear to involve activation of these cells, and the source of the elevation of neuropeptides remains to be clarified.
In contrast to the brain, no comparable changes in the blood-spinal cord barrier after spinal cord injury were induced by NAT (86), and SP immunoreactivity was decreased in the injured spinal cord (87) in contrast to the cerebrum. Hence, the involvement of the SP-NK1R axis in inflammation presumably differs between the brain and the spinal cord.
Remodeling of tendon and TKs in tendinopathy
SP synthesis has been found in nonnervous cells. Tendon primarily consists of tenocytes, which play a crucial role in tendinopathy. The overloading of tendons induces tendinopathy leading to their thickening, with a subsequent induction of tenocyte proliferation. Primary cultures of human tenocytes were immuno-positively stained for SP, and mechanical load flexion on the cells increased SP mRNA expression while decreasing NK1R mRNA expression (88). SP application to the cells accelerated tenocyte proliferation through activation of NK1R, suggesting the presence of an autocrine positive feedback loop in tenocytes in response to mechanical load.
Thickening of tendon associated with tendinopathy involves remodeling of the interstitial matrices. Interestingly, in a three-dimensional culture of human tenocytes, SP treatment induced an enhanced mRNA expression of collagen type III, smooth muscle actin, and matrix-metalloproteinase type 3 (MMP-3), which was blocked by an NK1R antagonist (89). As collagen type III is a substrate of MMP-3, this observation implies that SP induces the acceleration of interstitial collagen matrix remodeling in the tendon. It is possible that autocrine feedback regulation via the SP-NK1R axis underlies human tendinosis involving tenocyte proliferation. Conversely, SP injection into rabbit tendon increased tendon blood vessel formation and reduced peritendinous inflammation (90).
TKs and hematopoiesis
The majority of TKs are expressed in neurons, whereas some TKs are expressed in nonneuronal tissues such as blood cells. According to an in vitro study in humans, SP and NKA exert opposite, that is, stimulatory and inhibitory effects on hematopoiesis through NK1R and NK-2R, respectively (91). A study showed high levels of Tac4 expression in the thymus and bone marrow (9), implying a possible relation to lymphocyte functions. Tac4 encodes HK-1 and EK, and HK-1 is a full agonist of NK1R in immunoresponses (15). As for lymphocyte functions, HK-1 was shown to enhance the proliferation of T-cell precursors and increase the number of thymocytes in fetal thymus organ cultures (92). As the proliferation of T-cell precursors in the fetal thymus is important to subsequent selection of T-cells in ontogenesis, HK-1 appears to play a crucial role in the development of T-cell-mediated acquired immunity. Moreover, HK-1 was reported to be an activator of B-cell proliferation and survival through the synergic action of the mitogen-activated protein kinase (MAPK) cascade in response to TLR signals such as LPS in mice (93). In humans, the expression of hHK-1 and NK1R, but not SP, was similarly detected in B cells, and in contrast to SP, hHK-1 was able to induce human pre-B-cells proliferation through an NK1R-independent mechanism (94). By contrast, the knockout of Tac4 (Tac4-/-) increased the pro-B-cell population twofold in mouse bone marrow in vivo and in in vitro culture (95). HK-1 application to cell cultures of long-term reconstituting stem cells led to a more than 80% decrease in de novo generated pro-B cells (in terms of absolute cell number), without inducing cell death. Therefore, the effects of HK-1 on B-cell hematopoiesis remain controversial. Indeed, hHK-1 was up-regulated in B cells from chronic lymphocytic leukemia and non-Hodgkin’s lymphoma, whereas it was down-regulated in cells from acute lymphoblastic leukemia. The role of HK-1 in each pathological process remains, therefore, to be clarified.
In a long-term bone marrow repopulation study, a cell culture taken from Tac1r-/- knockout mice showed a lower population of B and T cells (96), implying that NK1R has a role in hematopoiesis. However, B- and T-cell engraftment deficiencies in Tac1r-/- knockout mice in vivo indicate that NK1R signaling has a role in hematopoiesis of lymphocytes, although this role is not indispensable as Tac1r-/- knockout mice are still capable of repopulation with B and T cells.
Bone marrow supplies progenitor cells (PCs) to peripheral blood. In mice, sympathetic sensory nerve fibers containing SP innervate the bone marrow and mediate a variety of responses related to injury via the release of TKs (97). The sympathetic fibers accelerate migration of PCs from the bone marrow to the peripheral blood (98), and inflammatory pain causes the migration of PCs into the circulation (99). The sympathetic fibers in the bone marrow express SP, and topoietic cells in the bone marrow abundantly express NK1R (100). An in vivo study in mice demonstrated that artificial myocardial infarction increased the levels of SP in the circulation and NK1R-positive granulocytes in the blood, and a single treatment with systemic injection of SP increased granulocytes in the blood (99). Thus, SP in association with ischemia appears to increase the number of granulocytes in circulation. Morphine treatment attenuated the mobilization of cell in circulation, and NK1R-/- knockout cells transplanted into wild-type mice did not show any increase in granulocytes, supporting the notion that pain and NK1R have roles in cell mobilization (99).
It is worthy of note that pretreatment to cultured human mesenchymal stem cells with SP led to a decrease in apoptotic cells after irradiation (101), implying the potential therapeutic use of SP pretreatment in minimizing radiation damage to patients.
TKs and venous thromboembolism
TKs, particularly SP and EKA/B, play crucial roles in the regulation of platelet functions, and they are known to stimulate platelet aggregation, presumably through NK1R and NK3R activation and intracellular Ca2+ mobilization (102). Among the various TKs, SP was shown to be stored in platelets (103), and SP acts in an autocrine or paracrine manner on aggregation (102).
Prevention of thromboembolism is crucial not only to the surgical treatment and postoperative care of patients but also to venous thrombosis after long-haul flights (traveler’s syndrome) (104) and long hours of sedentary office work common to the modern lifestyle among urban workers. Regarding clot formation, SP was shown to accelerate platelet-dependent clot formation via NK1R (105), while an NK1R antagonist upregulated tissue-plasminogen activator, enhancing cleavage of clots (106). SP and NK1R knockout reduced thrombus formation in response to immobilized collagen (107). Taken together, these facts indicate that the blockade of SP-NK1R can prevent clot formation. Moreover, full-length transcript variants of NK1R mRNA are known to have a higher affinity for SP than the truncated form (108). The expression ratio of the full-length form of NK1R is lower than that of the truncated form, and the full-length form was not observed in every sample (109). In fact, clot formation was enhanced in people who express the full-length form of the mRNA. Therefore, the full-length variant of NK1R has the potential to be a biomarker for clot formation in humans and could aid in the prevention of thromboembolism (109).
TKs and taste cells
Taste receptors (TRs) are classified into type I (T1R) and type II, with the former further classified into subtypes T1R1, T1R2, and T1R3 (110). A heterodimer of T1R1 plus T1R3 functions as an umami receptor, while a heterodimer of T1R2 plus T1R3 functions as a sweet receptor. Taste cells are also classified into type I (glial cells), type II (taste cells), and type III (presynaptic cells), and TRs are located in type II cells. Ca2+ imaging of isolated taste cells demonstrated that SP in a nanomole range induces cytosolic fluctuations of Ca2+ in some populations of taste cells, and the response was inhibited by an NK1R antagonist and, to a lesser extent, by an NK2R antagonist at higher concentration ranges (111). Although mouse taste cells express both NK1R and NK2R mRNA, single-cell reverse transcription and polymerase chain reaction revealed that a Ca2+ response was induced in type-I glial cells and type-II umami-receptor cells, suggesting that spicy foods stimulate SP release to enhance umami taste. The involvement of TKs in taste cells implies the revolutionary development of TK neurons not only as irritant receptor against risk factors and toxins but also to play a role in the palatability of foods.
NKB and gonadotropin secretion
Pulsatile secretion of gonadotropin-releasing hormone (GnRH) triggers the onset of puberty in males and females (112). GnRH pulses gradually potentiate the secretion of pituitary gonadotropins, luteinizing hormone (LH), and follicle stimulating hormone, which stimulate estradiol (E2) synthesis and secretion in the gonadotroph to elicit humoral and behavioral changes in animals. E2 regulates feedback to the hypothalamus, whereas E2 has dual actions on the hypothalamus either to reduce or to enhance gonadotropin secretion depending on the stage of the ovarian cycle.
Previous reports concerning the regulation of GnRH secretion in the hypothalamus are summarized in Figure 2. In immunohistochemical studies, estrogen receptor α (ERα) was found in NKB-immunopositive neurons proximally to GnRH neurons in the mouse and ovine arcuate nucleus (ARC) where the GnRH pulse generator exists (113). The number of NKB-positive cells is high in the ARC of females, implying that NKB has a role as an epistatic factor in the regulation of GnRH secretion. Indeed, the NK3R agonist senktide has been shown to increase LH secretion after IV and ICV injection in mice (114), rats (115), sheep (116), and monkey (117).
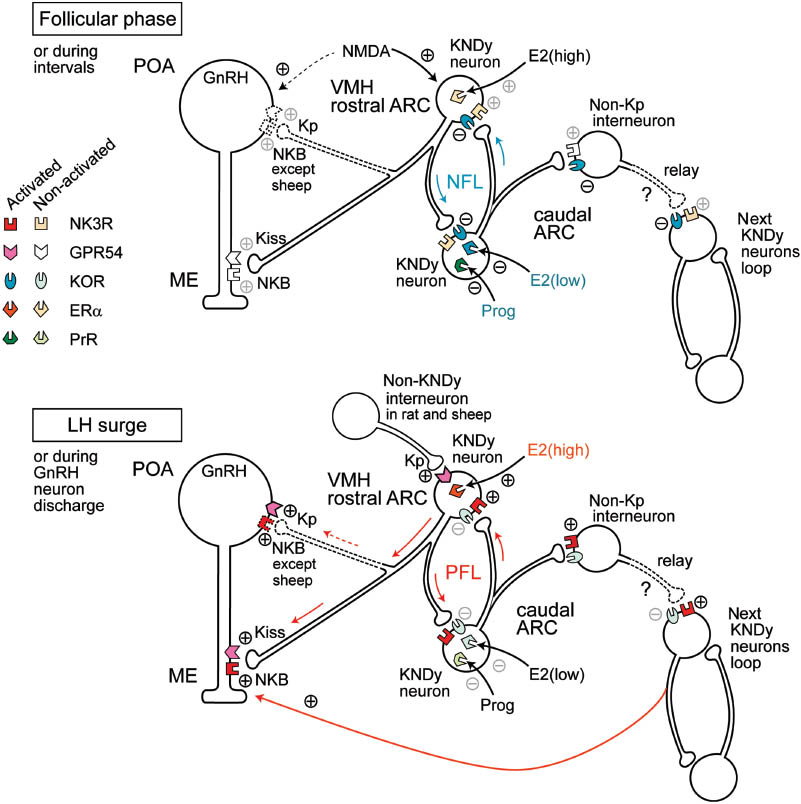
A model for the regulatory mechanism of GnRH secretion in the hypothalamus.
The upper illustration shows the hypothetized regulatory mechanism of the negative feedback loop during the follicular phase by low levels of peripheral estradiol or the interval period of GnRH neuron discharge. The lower illustration shows the mechanism of the positive feedback loop during the luteinizing hormone (LH) surge by high levels of peripheral estradiol or the discharge period of GnRH neurons. The colored symbols indicate the activated forms of receptors. Localizations of ERα remain speculative. ER α, estrogen receptor α; GnRH, gonadotropin releasing hormone; GPR54, G-protein coupled receptor 54 (as kisspeptin receptor); KOR, κ-opioid receptor; Kp, kisspeptin; NK3R, neurokinin receptor type 3; NMDA, N-methyl-d-asparate; PrR, progesterone receptor; ARC, arcuate nucleus; ME, median eminence, POA, preoptic area.
The Kiss1 gene product kisspeptin1 and its G-protein-coupled receptor type 54 (GPR54) (118) also plays a crucial role in the regulation of GnRH secretion (119) (Figure 2). Kisspeptin has a potent stimulatory effect on GnRH secretion in the male mouse (120), and a mutant of GPR54 causes hypogonadtropic hypogonadism (HH) in mice (121).
The cell bodies of GnRH neurons are in the preoptic area (POA) of the hypothalamus and extend fibers to the median eminence (ME) (Figure 2). NKB-immunopositive neurons are abundant in the caudal ARC (113), and they possess kisspeptin and dynorphin (Dyn) (122–124). Thus, kisspeptin-, NKB-, and Dyn-positive neurons, referred to as KNDy neurons, have been shown to play a central role in the ARC for the central regulation of GnRH secretion. As KNDy neurons are in contact with cell bodies and the nerve fibers of GnRH neurons, kisspeptin and NKB can stimulate both cell bodies and the nerve fibers of GnRH neurons through GPR54 and NK3R, respectively (125). However, as an exception, NKB cannot stimulate the cell bodies of GnRH neurons in sheep, because NK3R is not found there (126). Moreover, KNDy neurons can contact each other via the varicosities of nerve fibers to form a feedback loop in the ARC (125).
KNDy neurons possess both NK3R and κ-opioid receptors (KOR) (127). In general, hypothalamic NKB neurons enhance GnRH secretion, which has been supported by observations that mutations of the Tac3 (Ppt-b) gene encoding NKB play a role in HH in humans (128). Further, NKB neurons in the hypothalamus have been shown to possess Dyn immunoreactivity (127), and Dyn was shown to inhibit electrical bursts in the ARC in sheep and goats (129, 130), suggesting inhibitory autocrine regulation of KNDy neurons. Taken together, it can be said that NKB triggers a positive excitatory feedback loop of KNDy neurons, while Dyn triggers a negative inhibitory feedback loop (Figure 2). The former accelerates GnRH secretion, whereas the latter reduces it. As KNDy neurons are abundant in the ARC (123), there is presumably a number of neuron pairs forming feedback loops. Non-kisspeptin interneurons presumably relay acceleration of the positive feedback loops in rats and sheep (131).
In mice, kisspeptin neurons in the ARC are inhibited by Dyn via KOR and stimulated by NKB. However, the effect of NKB has been shown to be blocked by a mixture of three NKR subtype antagonists, not only by those NK3R antagonists that inhibit the effect of senktide in the male mouse (132), suggesting that NKB activates kisspeptin neurons through multiple subtypes of NKR in the male mouse. In addition, the three major tachykinin peptides directly activate nearly all kisspeptin neurons in the ARC, indicating the existence of a complex regulatory system for GnRH secretion.
Although it remains unclear as to what initiates the positive and negative feedback loops as epistatic factors to KNDy neurons in the ARC, recent studies have revealed a peripheral feedback system to KNDy neurons. The knockout of aromatase, which synthesizes E2 in the female mouse, was shown to reduce kisspeptin expression in the rostral periventricular region of the third ventricle (133), suggesting that E2 input to the hypothalamus potentiates kisspeptin neurons, presumably to generate the pre-ovulatory LH surge in the adult female mouse. Kisspeptin is expressed in NKB- and Dyn-negative neurons in the hypothalamus, particularly in the POA in ewes (124), and such a neuron may relay the E2 excitatory signal to the GnRH neurons in the POA or KNDy neurons in the ARC immediately before the LH surge, as is the case of rodents (133). In contrast to kisspeptin, E2 inhibits the expression of NKB in the ARC in mice (127) and ovarinectomized goat (130), suggesting that the NKB-NK3R axis is, in part, involved in the negative feedback regulation of GnRH secretion by E2. As ERα is found abundantly in KNDy neurons, peripheral E2 signals from the ovary are likely to inhibit or activate the feedback loop (Figure 2); that is, low levels of E2 stimulate the negative feedback loop in the follicular phase, and high level of E2 stimulates the positive feedback loop leading to the LH surge. Progesterone receptors (PrR) found in KNDy neurons are likely to be involved in the stimulation of the negative feedback loop in the luteal phase and, presumably, in pregnancy (134).
Figure 2 shows the hypothesized model of the relation between KNDy neurons in the ARC and peripheral feedback signals, and it is drawn as if different types of KNDy neurons receive the inhibitory and excitatory input of E2 at low and high levels, respectively, although this is based simply on the notion that E2 and progesterone inhibits GnRH secretion in early follicular phase and luteal phase, respectively, while a high level of E2 accelerates GnRH secretion in late follicular phase. However, it is possible that identical neurons in the ARC receive all of these signals. A very high percentage of the KNDy neurons possess both the receptors (124, 135), whereas it remains unclear which KNDy neurons receive E2 signals that trigger either the positive or negative feedback loop in the ARC. At present, no dimorphism of KNDy neurons that respond differently to E2 depending on levels have been distinguished by immunohistochemical studies, and their morphological distinction remains uncertain. It is possible that there are functional differences in KNDy neurons in the ARC (Figure 2), while another possibility is that other NKB-positive neurons from outside the ARC trigger the positive response leading to the LH surge, because NKB-immunoreactivity is also concentrated in other hypothalamic regions (113, 131), and ERα and PrR are located in hypothalamic neurons other than KNDy neurons in the ARC (135). Further investigations are needed to elucidate the superior signals for the hypothalamic regulatory mechanism underlying GnRH secretion.
Central administration of senktide evoked LH secretion during prepuberty in male and female rats (136), while the response was abolished in the postpubertal male rat only. Gonadectomy without sex steroid supplementation in adult mice reversed the effects of senktide in both sexes. Thus, the sex steroids can be seen to influence the responsiveness of GnRH neurons to NKB, and these neurons maintain their positive responsiveness to NKB in females but not in males after the onset of puberty. On the other hand, neonatal E2 treatment reduced the number of NKB-immunopositive neurons in the ARC as well as the level of LH circulating in both sexes in rats, whereas senktide injection recovered the LH level to the normal range in both sexes (136). These results imply that the NKB-NK3R axis has a key role in facilitating GnRH secretion and, simultaneously, the fragility of the signal system to exposure to sex steroids and presumably environmental hormones in neonates. Indeed, ICV injection of senktide increased LH secretion during diestrus and proestrus phases of the menstrual cycle in intact female rats (that is, it triggers a premature LH surge). However, recent studies have also revealed an opposite action; that is, ICV injection of senktide reduces the plasma concentration of LH in ovarinectomized rats (115, 137). Conversely, senktide increases LH secretion in male mice (114, 138), whereas it inhibits LH secretion in ovarinectomized female mice (127). In sheep, senktide implantation in the retrochiasmatic area elevates plasma LH concentration in the anestrous and follicular phases of ewes, although it has no effect in the luteal phase (116). In contrast, senktide was found to inhibit LH secretion but increase frequency of electrical impulses in the ARC in ovarinectomized goats (130). These conflicting effects imply that the NKB-NK3R axis modulates GnRH secretion via multiple pathways, depending not only on animal species but also on gender and the hormonal background of the follicular cycle (139).
In sheep, prenatal testosterone treatment reduces the number of NKB-, Dyn-, and progesterone-positive cells in the female ARC to levels comparable to those in normal males but does not affect the number of kisspeptin cells (135). Prenatal exposure of the brain to testosterone is known to play a key role in sex differentiation and the maintenance of the response to the negative feedback of progesterone. In normal adult rams, the number of KNDy neurons in the ARC decreases to half of that in adult females in association with a decrease in responsiveness to the feedback of progesterone. Hence, the aforementioned report implies that persistent kisspeptin cells might maintain responsiveness to progesterone in females, thus salvaging GnRH secretion.
The postnatal development of the kisspeptin-GPR54 signaling system was reviewed by Clarkson et al. (140). Although both NKB and kisspeptin stimulate GnRH secretion in mice, the individual roles remain to be clarified. The expression of Kiss1 mRNA encoding for kisspeptin increased in the ARC prior to the onset of puberty in GnRH/sex steroid-deficient hpg mice, whereas Tac3 encoding NKB increased at puberty in both wild-type and hpg mice (141), suggesting that neurons involved in negative feedback regulation have different sensitivities to sex steroid. Indeed, E2 administration revealed that Kiss1 gene expression was more sensitive than Tac2 expression in the ARC (141). However, the prepubertal treatment of wild-type and hpg mice with an NK3R antagonist did not alter the onset of puberty. Thus, the NKB-NK-3R axis alone seems insufficient to induce puberty in mice. On the other hand, whereas acute ablation of kisspeptin neurons inhibits fertility in adult female mice, neither ablation of kisspeptin neurons nor that of GPR54-expressing neurons in knockout mice altered the onset of puberty or fertility in female mice (142). Thus, compensation for the kisspeptin-GPR54 axis might occur via genetic ablation, but not via acute ablation, at an early stage of development in the female mouse.
The role of the NKB-NK3R axis was recently evaluated in male and female mice in relation to postnatal nutrient status (143). Body weight of pups increased depending on the amount of feed before and after weaning, and the density of kisspeptin and NKB-immunopositive neurons in the median preoptic nucleus projecting to the hypothalamic regions was lower in the under-nourished group at 32 days and in adult diestrus female mice, indicating that the ARC circuits are highly plastic and develop in response to food availability. The observation confirmed the impact of nutrition status at the early neonatal stages on the plasticity of the hypothalamic neural circuits regulating mammalian reproduction in adults.
Pheromone exposure, by the introduction of male sheep, caused an elevation in LH level in ewes during seasonal anestrus; however, pretreatment with a kisspeptin antagonist into the lateral ventricles in the ewes abolished this elevation in LH (144). After male exposure, Kiss1 mRNA was increased in the ARC, whereas NKB mRNA was decreased. These observations imply that kisspeptin neurons, but not NKB neurons, mediate the effect of pheromone on the activation of GnRH and LH secretion.
Distribution and roles of TKs in the brain stem
The brainstem plays a crucial role in the regulation of the autonomic nervous system innervating a variety of organs and tissues. The distribution of SP and NKA encoded by the same Tac1 gene has been reported in the human brainstem, and, in general, these two TKs show quite similar distributions (145).
The paratrigeminal respiratory group in the brainstem is related to the rhythm generation of respiration in lampreys, in which the glutamatergic neurons are surrounded by SP-immunopositive neurons (146). A glutamate receptor blockade induces apnea, whereas microinjection of SP into this region restored respiratory rhythm, indicating that SP contributes to the generation of respiratory rhythm in vertebrates.
TKs, SP in particular, are known to be cough inducers in the CNS, and NKR antagonist treatment has been shown to suppress cough in several species (147). Cough is induced by a series of complex mechanisms involving integration of peripheral sensory input from the airways and reconfiguration of inspiratory and expiratory motor neuron firing patterns (148). Various endogenous and exogenous factors are involved in the cough reflex, with afferent C-fiber activation in the airways being insufficient itself to induce the cough reflex. However, the micro injection of SP in the commissural subnucleus of the nucleus of the solitary tract (NST) enhances cough reflex sensitivity to topical citric acid exposure in the guinea pig trachea (149), indicating that signals inducing the cough reflex through C-fibers coming from the airways converge in the NST, and NKR is involved in the regulation of the reflex threshold.
In comparison with SP and NKA, which are widely distributed in both the CNS and PNS, NKB is predominantly distributed in the CNS (150). The distribution of NKB is restricted to 19 nuclei/regions of the brainstem (151) with the major focus being the pons. Moderate distribution has also been in the reticular formation, periaqueductal central gray, inferior salivatory nucleus, NST, gelatinosa trigeminal nucleus, and spinal trigeminal nucleus. This distribution pattern implies the functional involvement of NKB in the regulation of the cardiovascular, respiratory, and gastrointestinal systems. However, the roles of NKB in the brainstem remain to be determined.
Roles of TKs in neurogenic diseases
Schizophrenia
NK3R modulates the activity of dopaminergic neurons of the ventral tegmental area. Systemic administration of the dopaminergic receptor agonist, apomorphine, was shown to induce human schizophrenia-like behavior in rats, in which internalization of NK3R was induced (152). An NK-3R antagonist SB222200 prevented the apomorphine-induced increase in cytosolic density of NK3R, implying a potential therapeutic use of NK3R antagonists in the treatment of schizophrenia.
Alzheimer’s disease (AD)
AD is a neurodegenerative disorder in which a characteristic amyloid plaque in the brain is formed by aggregated β sheets (fibrils) of amyloid β (Aβ). Aβ is a peptide consisting of approximately 40 amino acid residues and exhibits neurotoxicity. TKs are known to inhibit the toxicity of Aβ. Although the precise mechanism of TK activity remains unknown, the similarity of residues 25–35 region of Aβ to those of the three major TK peptides (SP, NKA, and NKB) suggest that it plays a crucial role (153). In particular, residues 31 and 33–35 are identical; that is, FxGLM-NH2 (with a hydrophobic x residue), which contributes to its amyloidogenicity through strong aromatic amino acid interactions. Indeed, the addition of tryptophan (W) to the carboxyl terminal of TKs effectively attenuates the formation of Aβ fibrils (153), implying that modification of the three TK peptides could lead to the development of anti-AD compounds.
On the other hand, because the NK2R gene is located in the linkage region for AD on chromosome 10p21, it was regarded as a candidate gene for AD. However, gene analysis has demonstrated that it has no association with AD (154). Hence, the NK2R gene was excluded as a risk factor for AD.
Huntington’s disease (HD)
The amygdala may be involved in the development of motion sicknesses such as HD. In rats, hypergravity-induced motion sickness increases Tac1 mRNA, but not NK1R mRNA, in the amygdala and NST, whereas motion sickness was inhibited by the systemic administration of the NK1R antagonist CP-99,994 (155). This suggests that SP neuron stimulation by hypergravity in the amygdala may be involved in the development of motion sickness in this rat model.
TKs and metal ions in the CNS
The redox-active metals such as copper are known to increase in concentration with aging, and they have been shown to be the cause of various neurological diseases (156). The inhibition of copper uptake appears to protect against copper-induced intracellular calcium fluctuations. Although SP does not exert any copper-binding activity, a novel protective role of NKB in AD was recently demonstrated (157); that is, two NKB molecules can bind a copper ion, preventing copper from entering astrocytes. In the cerebrospinal fluid of HD, free copper ion concentration was shown to increase, while copper/superoxide dismutase (SOD) activity decreased according to the stage of the disease (158). The latter change increased oxidative stress, resulting in tissue damage. Recently, NPK as a major TK in the cerebral cortex and hippocampus was also shown to be able to bind copper ions (159), implying that local changes in NPK in the brain may alter copper concentrations as a potential causative factor for neurological disease such as HD. Indeed, SP, NKA, NKB, and NPK were decreased in the substantia nigra of the HD (160). NPγ also binds copper ions with increased pH (161).
Conclusions
While the roles of TKs have been investigated for over 80 years, the frontiers of TK study around the globe continue to expand. In the last decade, the physiological roles of hypothalamic NKB in relation to puberty and GnRH secretion have been determined in several mammalian species, although they have yielded conflicting results and the mechanisms have not yet been theoretically integrated. Other physiological and pathophysiological roles of TKs in the CNS are not sufficiently clarified, though their localization has been determined. Thus, further investigations are needed and are expected to elucidate the roles of TKs. On the other hand, the representative role of the SP and NK1R axis in pain and inflammation seems to be more or less established, and many NK1R antagonists have been shown to have potential therapeutic uses in the control of disease. However, they have not yet fulfilled the criteria for establishment as clinical drugs, and no NK1R antagonist has yet emerged as a clinically available drug other than as an antiemetic. This is presumably in part due to the range of activities of most TKs, which can lead to adverse effects. The situation with TKs contrasts markedly with that of other drugs, such as cyclooxygenase inhibitors, opioids agonists, and NMDA receptor antagonists currently available for clinical use for the control of pain and inflammation. Thus, further development of selective and safe NKR antagonists is needed as part of the future study of TKs. Although much hard work is yet required, future studies on the physiological and pathophysiological roles of TKs should simultaneously be beneficial, and it will simultaneously open the door to the clinical use of TK antagonists.
Highlights
SP plays crucial roles in pain processing and inflammation in the peripheral tissue in cooperation with TRP and in the regulation of BBB permeability and microglia in inflammation in the CNS.
TKs are involved in hematopoiesis in the bone marrow, and the molecular structure of NKR influences the onset of venous thromboembolism.
NKB binds copper ions in the CNS, which may be helpful for the prevention of neurogenic movement diseases such as AD and HD.
KNDy neurons in the positive and negative feedback loops in the ARC might consist of neuron couples and regulate GnRH secretion from the ME.
About the author

Takenori Onaga received his Bachelor’s degree of Doctor of Veterinary Medicine from the Rakuno Gakuen University (Ebetsu, Japan) in 1990. He started his research on bioactive peptides in the laboratory of veterinary physiology in the same university and, in this way, received research training with Dr. Satoru Naruse and Dr. Atsukazu Kuwahara in the Institute of Physiological Science (Okazaki, Japan). He earned his PhD in Veterinary Medicine from the Hokkaido University (Sapporo, Japan) under the supervision of Dr. Yoshiaki Habara in 2001 and continued his research in the aforementioned laboratory. He is a Professor in the present Laboratory since 2009.
Acknowledgments
I sincerely appreciate all the work performed by researchers in association with the various studies on TKs. I thank Dr. Garry Heterick for his help in correcting the original manuscript. The author has no conflicts of interest to disclose.
References
1. Chang MM, Leeman SE, Niall HD. Amino-acid sequence of substance P. Nat New Biol 1971; 232: 86–7.10.1038/newbio232086a0Suche in Google Scholar
2. Nawa H, Hirose T, Takashima H, Inayama S, Nakanishi S. Nucleotide sequences of cloned cDNAs for two types of bovine brain substance P precursor. Nature 1983; 306: 32–6.10.1038/306032a0Suche in Google Scholar
3. Tatemoto K, Lundberg JM, Jornvall H, Mutt V. Neuropeptide K: isolation, structure and biological activities of a novel brain tachykinin. Biochem Biophys Res Commun 1985; 128: 947–53.10.1016/0006-291X(85)90138-XSuche in Google Scholar
4. Roth KA, Makk G, Beck O, Faull K, Tatemoto K, Evans CJ, Barchas JD. Isolation and characterization of substance P, substance P 5-11, and substance K from two metastatic ileal carcinoids. Regul Pept 1985; 12: 185–99.10.1016/0167-0115(85)90060-6Suche in Google Scholar
5. Kage R, McGregor GP, Thim L, Conlon JM. Neuropeptide-gamma: a peptide isolated from rabbit intestine that is derived from gamma-preprotachykinin. J Neurochem 1988; 50: 1412–7.10.1111/j.1471-4159.1988.tb03024.xSuche in Google Scholar
6. Kurtz MM, Wang R, Clements MK, Cascieri MA, Austin CP, Cunningham BR, Chicchi GG, Liu Q. Identification, localization and receptor characterization of novel mammalian substance P-like peptides. Gene 2002; 296: 205–12.10.1016/S0378-1119(02)00861-2Suche in Google Scholar
7. Nawa H, Doteuchi M, Igano K, Inouye K, Nakanishi S. Substance K: a novel mammalian tachykinin that differs from substance P in its pharmacological profile. Life Sci 1984; 34: 1153–60.10.1016/0024-3205(84)90087-0Suche in Google Scholar
8. Biran J, Palevitch O, Ben-Dor S, Levavi-Sivan B. Neurokinin Bs and neurokinin B receptors in zebrafish-potential role in controlling fish reproduction. Proc Natl Acad Sci USA 2012; 109: 10269–74.10.1073/pnas.1119165109Suche in Google Scholar PubMed PubMed Central
9. Zhang Y, Lu L, Furlonger C, Wu GE, Paige CJ. Hemokinin is a hematopoietic-specific tachykinin that regulates B lymphopoiesis. Nat Immunol 2000; 1: 392–7.10.1038/80826Suche in Google Scholar PubMed
10. Page NM. Hemokinins and endokinins. Cell Mol Life Sci 2004; 61: 1652–63.10.1007/s00018-004-4035-xSuche in Google Scholar PubMed
11. Munekata E, Kubo K, Tanaka H, Osakada F. Structure-activity studies of heptapeptide derivatives related to substance P, neurokinin A, B and other tachykinins on smooth muscles. Peptides 1987; 8: 169–73.10.1016/0196-9781(87)90182-3Suche in Google Scholar
12. Pennefather JN, Patak E, Pinto FM, Candenas ML. Mammalian tachykinins and uterine smooth muscle: the challenge escalates. Eur J Pharmacol 2004; 500: 15–26.10.1016/j.ejphar.2004.07.007Suche in Google Scholar
13. Regoli D, Rhaleb NE, Rouissi N, Tousignant C, Jukic D, Drapeau C. Activation of sensory nerves by kinins: pharmacologic tools for studying kinin receptors. Adv Exp Med Biol 1991; 298: 63–74.10.1007/978-1-4899-0744-8_6Suche in Google Scholar
14. Bellucci F, Carini F, Catalani C, Cucchi P, Lecci A, Meini S, Patacchini R, Quartara L, Ricci R, Tramontana M, Giuliani S, Maggi CA. Pharmacological profile of the novel mammalian tachykinin, hemokinin 1. Br J Pharmacol 2002; 135: 266–74.10.1038/sj.bjp.0704443Suche in Google Scholar
15. Morteau O, Lu B, Gerard C, Gerard NP. Hemokinin 1 is a full agonist at the substance P receptor. Nat Immunol 2001; 2: 1088.10.1038/ni1201-1088Suche in Google Scholar
16. Dam TV, Takeda Y, Krause JE, Escher E, Quirion R. Gamma-preprotachykinin-(72-92)-peptide amide: an endogenous preprotachykinin I gene-derived peptide that preferentially binds to neurokinin-2 receptors. Proc Natl Acad Sci USA 1990; 87: 246–50.10.1073/pnas.87.1.246Suche in Google Scholar
17. Dike A, Cowsik SM. Three-dimensional structure of neuropeptide k bound to dodecylphosphocholine micelles. Biochemistry 2006; 45: 2994–3004.10.1021/bi052287oSuche in Google Scholar
18. Chandrashekar IR, Dike A, Cowsik SM. Membrane-induced structure of the mammalian tachykinin neuropeptide gamma. J Struct Biol 2004; 148: 315–25.10.1016/j.jsb.2004.08.008Suche in Google Scholar
19. Steinhoff MS, von Mentzer B, Geppetti P, Pothoulakis C, Bunnett NW. Tachykinins and their receptors: contributions to physiological control and the mechanisms of disease. Physiol Rev 2014; 94: 265–301.10.1152/physrev.00031.2013Suche in Google Scholar
20. Polak JM, Bloom SR. The peripheral substance P-ergic system. Peptides 1981; 2:(Suppl 2): 133–48.10.1016/0196-9781(81)90024-3Suche in Google Scholar
21. Mandadi S, Roufogalis BD. Thermo TRP channels in nociceptors: taking a lead from capsaicin receptor TRPV1. Curr Neuropharmacol 2008; 6: 21–38.10.2174/157015908783769680Suche in Google Scholar PubMed PubMed Central
22. Cattaruzza F, Spreadbury I, Miranda-Morales M, Grady EF, Vanner S, Bunnett NW. Transient receptor potential ankyrin-1 has a major role in mediating visceral pain in mice. Am J Physiol Gastrointest Liver Physiol 2010; 298: G81–91.10.1152/ajpgi.00221.2009Suche in Google Scholar PubMed PubMed Central
23. Heng YJ, Saunders CI, Kunde DA, Geraghty DP. TRPV1, NK1 receptor and substance P immunoreactivity and gene expression in the rat lumbosacral spinal cord and urinary bladder after systemic, low dose vanilloid administration. Regul Pept 2011; 167: 250–8.10.1016/j.regpep.2011.02.004Suche in Google Scholar PubMed
24. Hoffmeister C, Trevisan G, Rossato MF, de Oliveira SM, Gomez MV, Ferreira J. Role of TRPV1 in nociception and edema induced by monosodium urate crystals in rats. Pain 2011; 152: 1777–88.10.1016/j.pain.2011.03.025Suche in Google Scholar PubMed
25. Fernandes ES, Liang L, Smillie SJ, Kaiser F, Purcell R, Rivett DW, Alam S, Howat S, Collins H, Thompson SJ, Keeble JE, Riffo-Vasquez Y, Bruce KD, Brain SD. TRPV1 deletion enhances local inflammation and accelerates the onset of systemic inflammatory response syndrome. J Immunol 2012; 188: 5741–51.10.4049/jimmunol.1102147Suche in Google Scholar PubMed
26. Rivat C, Vera-Portocarrero LP, Ibrahim MM, Mata HP, Stagg NJ, De Felice M, Porreca F, Malan TP. Spinal NK-1 receptor-expressing neurons and descending pathways support fentanyl-induced pain hypersensitivity in a rat model of postoperative pain. Eur J Neurosci 2009; 29: 727–37.10.1111/j.1460-9568.2009.06616.xSuche in Google Scholar PubMed
27. Beaudry H, Dubois D, Gendron L. Activation of spinal mu- and delta-opioid receptors potently inhibits substance P release induced by peripheral noxious stimuli. J Neurosci 2011; 31: 13068–77.10.1523/JNEUROSCI.1817-11.2011Suche in Google Scholar PubMed PubMed Central
28. Hamity MV, White SR, Hammond DL. Effects of neurokinin-1 receptor agonism and antagonism in the rostral ventromedial medulla of rats with acute or persistent inflammatory nociception. Neuroscience 2010; 165: 902–13.10.1016/j.neuroscience.2009.10.064Suche in Google Scholar PubMed PubMed Central
29. Andoh T, Katsube N, Maruyama M, Kuraishi Y. Involvement of leukotriene B(4) in substance P-induced itch-associated response in mice. J Invest Dermatol 2001; 117: 1621–6.10.1046/j.0022-202x.2001.01585.xSuche in Google Scholar PubMed
30. Jeffry J, Kim S, Chen ZF. Itch signaling in the nervous system. Physiology (Bethesda) 2011; 26: 286–92.10.1152/physiol.00007.2011Suche in Google Scholar PubMed PubMed Central
31. Sun YG, Chen ZF. A gastrin-releasing peptide receptor mediates the itch sensation in the spinal cord. Nature 2007; 448: 700–3.10.1038/nature06029Suche in Google Scholar PubMed
32. Fleming MS, Ramos D, Han SB, Zhao J, Son YJ, Luo W. The majority of dorsal spinal cord gastrin releasing peptide is synthesized locally whereas neuromedin B is highly expressed in pain- and itch-sensing somatosensory neurons. Mol Pain 2012; 8: 52.10.1186/1744-8069-8-52Suche in Google Scholar PubMed PubMed Central
33. Akiyama T, Tominaga M, Davoodi A, Nagamine M, Blansit K, Horwitz A, Carstens MI, Carstens E. Roles for substance P and gastrin-releasing peptide as neurotransmitters released by primary afferent pruriceptors. J Neurophysiol 2013; 109: 742–8.10.1152/jn.00539.2012Suche in Google Scholar PubMed PubMed Central
34. Koga K, Chen T, Li XY, Descalzi G, Ling J, Gu J, Zhuo M. Glutamate acts as a neurotransmitter for gastrin releasing peptide-sensitive and insensitive itch-related synaptic transmission in mammalian spinal cord. Mol Pain 2011; 7: 47.10.1186/1744-8069-7-47Suche in Google Scholar PubMed PubMed Central
35. Akiyama T, Tominaga M, Takamori K, Carstens MI, Carstens E. Roles of glutamate, substance P, and gastrin-releasing peptide as spinal neurotransmitters of histaminergic and nonhistaminergic itch. Pain 2014; 155: 80–92.10.1016/j.pain.2013.09.011Suche in Google Scholar PubMed PubMed Central
36. Fukamachi S, Mori T, Sakabe J, Shiraishi N, Kuroda E, Kobayashi M, Bito T, Kabashima K, Nakamura M, Tokura Y. Topical cholecystokinin depresses itch-associated scratching behavior in mice. J Invest Dermatol 2011; 131: 956–61.10.1038/jid.2010.413Suche in Google Scholar PubMed
37. Raybould HE, Gayton RJ, Dockray GJ. Mechanisms of action of peripherally administered cholecystokinin octapeptide on brain stem neurons in the rat. J Neurosci 1988; 8: 3018–24.10.1523/JNEUROSCI.08-08-03018.1988Suche in Google Scholar
38. Newbold P, Brain SD. An investigation into the mechanism of capsaicin-induced oedema in rabbit skin. Br J Pharmacol 1995; 114: 570–7.10.1111/j.1476-5381.1995.tb17177.xSuche in Google Scholar PubMed PubMed Central
39. Bost KL. Tachykinin-mediated modulation of the immune response. Front Biosci 2004; 9: 3331–2.10.2741/1484Suche in Google Scholar PubMed
40. Bie B, Zhao ZQ. Peripheral inflammation alters desensitization of substance P-evoked current in rat dorsal root ganglion neurons. Eur J Pharmacol 2011; 670: 495–9.10.1016/j.ejphar.2011.09.004Suche in Google Scholar PubMed
41. Castillo C, Norcini M, Baquero-Buitrago J, Levacic D, Medina R, Montoya-Gacharna JV, Blanck TJ, Dubois M, Recio-Pinto E. The N-methyl-D-aspartate-evoked cytoplasmic calcium increase in adult rat dorsal root ganglion neuronal somata was potentiated by substance P pretreatment in a protein kinase C-dependent manner. Neuroscience 2011; 177: 308–20.10.1016/j.neuroscience.2010.12.040Suche in Google Scholar
42. Hui Y, Du J, Tang C, Bin G, Jiang H. Changes in arterial hydrogen sulfide (H(2)S) content during septic shock and endotoxin shock in rats. J Infect 2003; 47: 155–60.10.1016/S0163-4453(03)00043-4Suche in Google Scholar
43. Ang SF, Moochhala SM, MacAry PA, Bhatia M. Hydrogen sulfide and neurogenic inflammation in polymicrobial sepsis: involvement of substance P and ERK-NF-kappaB signaling. PLoS One 2011; 6: e24535.10.1371/journal.pone.0024535Suche in Google Scholar PubMed PubMed Central
44. Tamizhselvi R, Moore PK, Bhatia M. Hydrogen sulfide acts as a mediator of inflammation in acute pancreatitis: in vitro studies using isolated mouse pancreatic acinar cells. J Cell Mol Med 2007; 11: 315–26.10.1111/j.1582-4934.2007.00024.xSuche in Google Scholar PubMed PubMed Central
45. Trevisani M, Siemens J, Materazzi S, Bautista DM, Nassini R, Campi B, Imamachi N, Andre E, Patacchini R, Cottrell GS, Gatti R, Basbaum AI, Bunnett NW, Julius D, Geppetti P. 4-Hydroxynonenal, an endogenous aldehyde, causes pain and neurogenic inflammation through activation of the irritant receptor TRPA1. Proc Natl Acad Sci USA 2007; 104: 13519–24.10.1073/pnas.0705923104Suche in Google Scholar PubMed PubMed Central
46. Barbara G, Wang B, Stanghellini V, de Giorgio R, Cremon C, Di Nardo G, Trevisani M, Campi B, Geppetti P, Tonini M, Bunnett NW, Grundy D, Corinaldesi R. Mast cell-dependent excitation of visceral-nociceptive sensory neurons in irritable bowel syndrome. Gastroenterology 2007; 132: 26–37.10.1053/j.gastro.2006.11.039Suche in Google Scholar PubMed
47. Cottrell GS, Amadesi S, Schmidlin F, Bunnett N. Protease-activated receptor 2: activation, signalling and function. Biochem Soc Trans 2003; 31: 1191–7.10.1042/bst0311191Suche in Google Scholar PubMed
48. Divito SJ, Morelli AE, Larregina AT. Role of neurokinin-1 receptor in the initiation and maintenance of skin chronic inflammatory diseases. Immunol Res 2011; 50: 195–201.10.1007/s12026-011-8219-9Suche in Google Scholar PubMed
49. Cunin P, Caillon A, Corvaisier M, Garo E, Scotet M, Blanchard S, Delneste Y, Jeannin P. The tachykinins substance P and hemokinin-1 favor the generation of human memory Th17 cells by inducing IL-1beta, IL-23, and TNF-like 1A expression by monocytes. J Immunol 2011; 186: 4175–82.10.4049/jimmunol.1002535Suche in Google Scholar PubMed
50. Song IS, Bunnett NW, Olerud JE, Harten B, Steinhoff M, Brown JR, Sung KJ, Armstrong CA, Ansel JC. Substance P induction of murine keratinocyte PAM 212 interleukin 1 production is mediated by the neurokinin 2 receptor (NK-2R). Exp Dermatol 2000; 9: 42–52.10.1034/j.1600-0625.2000.009001042.xSuche in Google Scholar PubMed
51. Donnerer J, Schuligoi R, Stein C. Increased content and transport of substance P and calcitonin gene-related peptide in sensory nerves innervating inflamed tissue: evidence for a regulatory function of nerve growth factor in vivo. Neuroscience 1992; 49: 693–8.10.1016/0306-4522(92)90237-VSuche in Google Scholar
52. Joachim RA, Kuhlmei A, Dinh QT, Handjiski B, Fischer T, Peters EM, Klapp BF, Paus R, Arck PC. Neuronal plasticity of the “brain-skin connection”: stress-triggered up-regulation of neuropeptides in dorsal root ganglia and skin via nerve growth factor-dependent pathways. J Mol Med (Berl) 2007; 85: 1369–78.10.1007/s00109-007-0236-8Suche in Google Scholar PubMed
53. Grip L, Lonne-Rahm SB, Holst M, Johansson B, Nordlind K, Theodorsson E, El-Nour H. Substance P alterations in skin and brain of chronically stressed atopic-like mice. J Eur Acad Dermatol Venereol 2013; 27: 199–205.10.1111/j.1468-3083.2011.04443.xSuche in Google Scholar PubMed
54. Simpson J, Sundler F, Humes DJ, Jenkins D, Wakelin D, Scholefield JH, Spiller RC. Prolonged elevation of galanin and tachykinin expression in mucosal and myenteric enteric nerves in trinitrobenzene sulphonic acid colitis. Neurogastroenterol Motil 2008; 20: 392–406.10.1111/j.1365-2982.2007.01037.xSuche in Google Scholar PubMed
55. Engel MA, Leffler A, Niedermirtl F, Babes A, Zimmermann K, Filipovic MR, Izydorczyk I, Eberhardt M, Kichko TI, Mueller-Tribbensee SM, Khalil M, Siklosi N, Nau C, Ivanovic-Burmazovic I, Neuhuber WL, Becker C, Neurath MF, Reeh PW. TRPA1 and substance P mediate colitis in mice. Gastroenterology 2011; 141: 1346–58.10.1053/j.gastro.2011.07.002Suche in Google Scholar PubMed
56. Engel MA, Khalil M, Mueller-Tribbensee SM, Becker C, Neuhuber WL, Neurath MF, Reeh PW. The proximodistal aggravation of colitis depends on substance P released from TRPV1-expressing sensory neurons. J Gastroenterol 2012; 47: 256–65.10.1007/s00535-011-0495-6Suche in Google Scholar PubMed
57. Engel MA, Khalil M, Siklosi N, Mueller-Tribbensee SM, Neuhuber WL, Neurath MF, Becker C, Reeh PW. Opposite effects of substance P and calcitonin gene-related peptide in oxazolone colitis. Dig Liver Dis 2012; 44: 24–9.10.1016/j.dld.2011.08.030Suche in Google Scholar PubMed
58. Matsumoto K, Kurosawa E, Terui H, Hosoya T, Tashima K, Murayama T, Priestley JV, Horie S. Localization of TRPV1 and contractile effect of capsaicin in mouse large intestine: high abundance and sensitivity in rectum and distal colon. Am J Physiol Gastrointest Liver Physiol 2009; 297: G348–60.10.1152/ajpgi.90578.2008Suche in Google Scholar PubMed
59. Gillespie E, Leeman SE, Watts LA, Coukos JA, O’Brien MJ, Cerda SR, Farraye FA, Stucchi AF, Becker JM. Truncated neurokinin-1 receptor is increased in colonic epithelial cells from patients with colitis-associated cancer. Proc Natl Acad Sci USA 2011; 108: 17420–5.10.1073/pnas.1114275108Suche in Google Scholar PubMed PubMed Central
60. Lai JP, Lai S, Tuluc F, Tansky MF, Kilpatrick LE, Leeman SE, Douglas SD. Differences in the length of the carboxyl terminus mediate functional properties of neurokinin-1 receptor. Proc Natl Acad Sci USA 2008; 105: 12605–10.10.1073/pnas.0806632105Suche in Google Scholar PubMed PubMed Central
61. Douglas SD, Leeman SE. Neurokinin-1 receptor: functional significance in the immune system in reference to selected infections and inflammation. Ann NY Acad Sci 2011; 1217: 83–95.10.1111/j.1749-6632.2010.05826.xSuche in Google Scholar
62. Rosso M, Robles-Frias MJ, Covenas R, Salinas-Martin MV, Munoz M. The NK-1 receptor is expressed in human primary gastric and colon adenocarcinomas and is involved in the antitumor action of L-733,060 and the mitogenic action of substance P on human gastrointestinal cancer cell lines. Tumour Biol 2008; 29: 245–54.10.1159/000152942Suche in Google Scholar
63. Feng F, Yang J, Tong L, Yuan S, Tian Y, Hong L, Wang W, Zhang H. Substance P immunoreactive nerve fibres are related to gastric cancer differentiation status and could promote proliferation and migration of gastric cancer cells. Cell Biol Int 2011; 35: 623–9.10.1042/CBI20100229Suche in Google Scholar
64. Domotor A, Kereskay L, Szekeres G, Hunyady B, Szolcsanyi J, Mozsik G. Participation of capsaicin-sensitive afferent nerves in the gastric mucosa of patients with Helicobacter pylori-positive or-negative chronic gastritis. Dig Dis Sci 2007; 52: 411–7.10.1007/s10620-006-9180-7Suche in Google Scholar
65. Erin N, Turker S, Elpek O, Yildirim B. Differential changes in substance P, VIP as well as neprilysin levels in patients with gastritis or ulcer. Peptides 2012; 35: 218–24.10.1016/j.peptides.2012.03.018Suche in Google Scholar
66. Brancati SB, Zadori ZS, Nemeth J, Gyires K. Substance P induces gastric mucosal protection at supraspinal level via increasing the level of endomorphin-2 in rats. Brain Res Bull 2013; 91: 38–45.10.1016/j.brainresbull.2013.01.004Suche in Google Scholar
67. Barnes PJ. Neurogenic inflammation in the airways. Respir Physiol 2001; 125: 145–54.10.1016/S0034-5687(00)00210-3Suche in Google Scholar
68. Carr MJ, Undem BJ. Inflammation-induced plasticity of the afferent innervation of the airways. Environ Health Perspect 2001; 109: (Suppl 4): 567–71.10.1289/ehp.01109s4567Suche in Google Scholar
69. Sheldrick RL, Rabe KF, Fischer A, Magnussen H, Coleman RA. Further evidence that tachykinin-induced contraction of human isolated bronchus is mediated only by NK2-receptors. Neuropeptides 1995; 29: 281–92.10.1016/0143-4179(95)90037-3Suche in Google Scholar
70. Rogers DF, Belvisi MG, Aursudkij B, Evans TW, Barnes PJ. Effects and interactions of sensory neuropeptides on airway microvascular leakage in guinea-pigs. Br J Pharmacol 1988; 95: 1109–16.10.1111/j.1476-5381.1988.tb11745.xSuche in Google Scholar PubMed PubMed Central
71. Hsu CC, Lin RL, Lin YS, Lee LY. Bronchoconstriction induced by increasing airway temperature in ovalbumin-sensitized rats: role of tachykinins. J Appl Physiol (1985) 2013; 115: 688–96.10.1152/japplphysiol.00491.2013Suche in Google Scholar PubMed PubMed Central
72. Hox V, Vanoirbeek JA, Alpizar YA, Voedisch S, Callebaut I, Bobic S, Sharify A, De Vooght V, Van Gerven L, Devos F, Liston A, Voets T, Vennekens R, Bullens DM, De Vries A, Hoet P, Braun A, Ceuppens JL, Talavera K, Nemery B, Hellings PW. Crucial role of transient receptor potential ankyrin 1 and mast cells in induction of nonallergic airway hyperreactivity in mice. Am J Respir Crit Care Med 2013; 187: 486–93.10.1164/rccm.201208-1358OCSuche in Google Scholar PubMed
73. Hens G, Raap U, Vanoirbeek J, Meyts I, Callebaut I, Verbinnen B, Vanaudenaerde BM, Cadot P, Nemery B, Bullens DM, Ceuppens JL, Hellings PW. Selective nasal allergen provocation induces substance P-mediated bronchial hyperresponsiveness. Am J Respir Cell Mol Biol 2011; 44: 517–23.10.1165/rcmb.2009-0425OCSuche in Google Scholar PubMed
74. Joos GF, Germonpre PR, Pauwels RA. Role of tachykinins in asthma. Allergy 2000; 55: 321–37.10.1034/j.1398-9995.2000.00112.xSuche in Google Scholar PubMed
75. Cima K, Vogelsinger H, Kahler CM. Sensory neuropeptides are potent chemoattractants for human basophils in vitro. Regul Pept 2010; 160: 42–8.10.1016/j.regpep.2009.12.013Suche in Google Scholar PubMed
76. Teles AT, Kumagai Y, Brain SD, Teixeira SA, Varriano AA, Barreto MA, de Lima WT, Antunes E, Muscara MN, Costa SK. Involvement of sensory nerves and TRPV1 receptors in the rat airway inflammatory response to two environment pollutants: diesel exhaust particles (DEP) and 1,2-naphthoquinone (1,2-NQ). Arch Toxicol 2010; 84: 109–17.10.1007/s00204-009-0427-xSuche in Google Scholar PubMed
77. Nimmo AJ, Cernak I, Heath DL, Hu X, Bennett CJ, Vink R. Neurogenic inflammation is associated with development of edema and functional deficits following traumatic brain injury in rats. Neuropeptides 2004; 38: 40–7.10.1016/j.npep.2003.12.003Suche in Google Scholar PubMed
78. Chauhan VS, Sterka DG Jr., Gray DL, Bost KL, Marriott I. Neurogenic exacerbation of microglial and astrocyte responses to Neisseria meningitidis and Borrelia burgdorferi. J Immunol 2008; 180: 8241–9.10.4049/jimmunol.180.12.8241Suche in Google Scholar PubMed PubMed Central
79. Chauhan VS, Kluttz JM, Bost KL, Marriott I. Prophylactic and therapeutic targeting of the neurokinin-1 receptor limits neuroinflammation in a murine model of pneumococcal meningitis. J Immunol 2011; 186: 7255–63.10.4049/jimmunol.1100721Suche in Google Scholar PubMed PubMed Central
80. Zacest AC, Vink R, Manavis J, Sarvestani GT, Blumbergs PC. Substance P immunoreactivity increases following human traumatic brain injury. Acta Neurochir Suppl 2010; 106: 211–6.10.1007/978-3-211-98811-4_39Suche in Google Scholar PubMed
81. Turner RJ, Helps SC, Thornton E, Vink R. A substance P antagonist improves outcome when administered 4 h after onset of ischaemic stroke. Brain Res 2011; 1393: 84–90.10.1016/j.brainres.2011.03.066Suche in Google Scholar PubMed
82. Donkin JJ, Vink R. Mechanisms of cerebral edema in traumatic brain injury: therapeutic developments. Curr Opin Neurol 2010; 23: 293–9.10.1097/WCO.0b013e328337f451Suche in Google Scholar PubMed
83. Carthew HL, Ziebell JM, Vink R. Substance P-induced changes in cell genesis following diffuse traumatic brain injury. Neuroscience 2012; 214: 78–83.10.1016/j.neuroscience.2012.04.028Suche in Google Scholar PubMed
84. Donkin JJ, Cernak I, Blumbergs PC, Vink R. A substance P antagonist reduces axonal injury and improves neurologic outcome when administered up to 12 h after traumatic brain injury. J Neurotrauma 2011; 28: 217–24.10.1089/neu.2010.1632Suche in Google Scholar PubMed
85. Elliott MB, Oshinsky ML, Amenta PS, Awe OO, Jallo JI. Nociceptive neuropeptide increases and periorbital allodynia in a model of traumatic brain injury. Headache 2012; 52: 966–84.10.1111/j.1526-4610.2012.02160.xSuche in Google Scholar PubMed PubMed Central
86. Leonard AV, Vink R. The effect of an NK1 receptor antagonist on blood spinal cord barrier permeability following balloon compression-induced spinal cord injury. Acta Neurochir Suppl 2013; 118: 303–6.Suche in Google Scholar
87. Leonard AV, Thornton E, Vink R. Substance P as a mediator of neurogenic inflammation after balloon compression induced spinal cord injury. J Neurotrauma 2013; 30: 1812–23.10.1089/neu.2013.2993Suche in Google Scholar PubMed
88. Backman LJ, Fong G, Andersson G, Scott A, Danielson P. Substance P is a mechanoresponsive, autocrine regulator of human tenocyte proliferation. PLoS One 2011; 6: e27209.10.1371/journal.pone.0027209Suche in Google Scholar PubMed PubMed Central
89. Fong G, Backman LJ, Hart DA, Danielson P, McCormack B, Scott A. Substance P enhances collagen remodeling and MMP-3 expression by human tenocytes. J Orthop Res 2013; 31: 91–8.10.1002/jor.22191Suche in Google Scholar PubMed PubMed Central
90. Andersson G, Backman LJ, Scott A, Lorentzon R, Forsgren S, Danielson P. Substance P accelerates hypercellularity and angiogenesis in tendon tissue and enhances paratendinitis in response to Achilles tendon overuse in a tendinopathy model. Br J Sports Med 2011; 45: 1017–22.10.1136/bjsm.2010.082750Suche in Google Scholar PubMed
91. Rameshwar P, Gascon P. Hematopoietic modulation by the tachykinins. Acta Haematol 1997; 98: 59–64.10.1159/000203593Suche in Google Scholar PubMed
92. Zhang Y, Paige CJ. T-cell developmental blockage by tachykinin antagonists and the role of hemokinin 1 in T lymphopoiesis. Blood 2003; 102: 2165–72.10.1182/blood-2002-11-3572Suche in Google Scholar PubMed
93. Wang W, Li Q, Zhang J, Wu H, Yin Y, Ge Q, Zhang Y. Hemokinin-1 activates the MAPK pathway and enhances B cell proliferation and antibody production. J Immunol 2010; 184: 3590–7.10.4049/jimmunol.0901278Suche in Google Scholar PubMed
94. Grassin-Delyle S, Buenestado A, Vallat L, Naline E, Marx S, Decocq J, Debre P, Bernard OA, Advenier C, Devillier P, Merle-Beral H. Expression and proliferative effect of hemokinin-1 in human B-cells. Peptides 2012; 32: 1027–34.10.1016/j.peptides.2011.02.012Suche in Google Scholar PubMed
95. Berger A, Benveniste P, Corfe SA, Tran AH, Barbara M, Wakeham A, Mak TW, Iscove NN, Paige CJ. Targeted deletion of the tachykinin 4 gene (TAC4-/-) influences the early stages of B lymphocyte development. Blood 2010; 116: 3792–801.10.1182/blood-2010-06-291062Suche in Google Scholar PubMed
96. Berger A, Frelin C, Shah DK, Benveniste P, Herrington R, Gerard NP, Zuniga-Pflucker JC, Iscove NN, Paige CJ. Neurokinin-1 receptor signalling impacts bone marrow repopulation efficiency. PLoS One 2013; 8: e58787.10.1371/journal.pone.0058787Suche in Google Scholar PubMed PubMed Central
97. Imai S, Tokunaga Y, Maeda T, Kikkawa M, Hukuda S. Calcitonin gene-related peptide, substance P, and tyrosine hydroxylase-immunoreactive innervation of rat bone marrows: an immunohistochemical and ultrastructural investigation on possible efferent and afferent mechanisms. J Orthop Res 1997; 15: 133–40.10.1002/jor.1100150120Suche in Google Scholar PubMed
98. Katayama Y, Battista M, Kao WM, Hidalgo A, Peired AJ, Thomas SA, Frenette PS. Signals from the sympathetic nervous system regulate hematopoietic stem cell egress from bone marrow. Cell 2006; 124: 407–21.10.1016/j.cell.2005.10.041Suche in Google Scholar PubMed
99. Amadesi S, Reni C, Katare R, Meloni M, Oikawa A, Beltrami AP, Avolio E, Cesselli D, Fortunato O, Spinetti G, Ascione R, Cangiano E, Valgimigli M, Hunt SP, Emanueli C, Madeddu P. Role for substance P-based nociceptive signaling in progenitor cell activation and angiogenesis during ischemia in mice and in human subjects. Circulation 2012; 125: 1774–86, S1–19.10.1161/CIRCULATIONAHA.111.089763Suche in Google Scholar PubMed PubMed Central
100. Nance DM, Sanders VM. Autonomic innervation and regulation of the immune system (1987–2007). Brain Behav Immun 2007; 21: 736–45.10.1016/j.bbi.2007.03.008Suche in Google Scholar PubMed PubMed Central
101. An YS, Lee E, Kang MH, Hong HS, Kim MR, Jang WS, Son Y, Yi JY. Substance P stimulates the recovery of bone marrow after the irradiation. J Cell Physiol 2011; 226: 1204–13.10.1002/jcp.22447Suche in Google Scholar PubMed
102. Graham GJ, Stevens JM, Page NM, Grant AD, Brain SD, Lowry PJ, Gibbins JM. Tachykinins regulate the function of platelets. Blood 2004; 104: 1058–65.10.1182/blood-2003-11-3979Suche in Google Scholar
103. Nakano T, Shimomura T, Takahashi K, Ikawa S. Platelet substance P and 5-hydroxytryptamine in migraine and tension-type headache. Headache 1993; 33: 528–32.10.1111/j.1526-4610.1993.hed3310528.xSuche in Google Scholar
104. Schwarz T, Siegert G, Oettler W, Halbritter K, Beyer J, Frommhold R, Gehrisch S, Lenz F, Kuhlisch E, Schroeder HE, Schellong SM. Venous thrombosis after long-haul flights. Arch Intern Med 2003; 163: 2759–64.10.1001/archinte.163.22.2759Suche in Google Scholar
105. Azma T, Matsubara Y, Kinoshita H, Hidaka I, Shiraishi S, Nakao M, Kawamoto M, Yuge O, Hatano Y. Prothrombotic roles of substance-P, neurokinin-1 receptors and leukocytes in the platelet-dependent clot formation in whole blood. J Thromb Thrombolysis 2009; 27: 280–6.10.1007/s11239-008-0215-0Suche in Google Scholar
106. Reed GL. Platelet secretory mechanisms. Semin Thromb Hemost 2004; 30: 441–50.10.1055/s-2004-833479Suche in Google Scholar
107. Jones S, Gibbins JM. The neurokinin 1 receptor: a potential new target for anti-platelet therapy? Curr Opin Pharmacol 2008; 8: 114–9.10.1016/j.coph.2008.01.004Suche in Google Scholar
108. Fong TM, Yu H, Huang RR, Strader CD. The extracellular domain of the neurokinin-1 receptor is required for high-affinity binding of peptides. Biochemistry 1992; 31: 11806–11.10.1021/bi00162a019Suche in Google Scholar
109. Azma T, Sugimoto Y, Kinoshita H, Ito T, Tsukamoto M, Hoshijima H, Nakao M, Kikuchi H. Detection of the full-length transcript variant for neurokinin-1 receptor in human whole blood associated with enhanced reinforcement of clot by substance-P. J Thromb Thrombolysis 2012; 33: 329–37.10.1007/s11239-011-0650-1Suche in Google Scholar
110. Zhao GQ, Zhang Y, Hoon MA, Chandrashekar J, Erlenbach I, Ryba NJ, Zuker CS. The receptors for mammalian sweet and umami taste. Cell 2003; 115: 255–66.10.1016/S0092-8674(03)00844-4Suche in Google Scholar
111. Grant J. Tachykinins stimulate a subset of mouse taste cells. PLoS One 2012; 7: e31697.10.1371/journal.pone.0031697Suche in Google Scholar PubMed PubMed Central
112. Williams CL, Thalabard JC, O’Byrne KT, Grosser PM, Nishihara M, Hotchkiss J, Knobil E. Duration of phasic electrical activity of the hypothalamic gonadotropin-releasing hormone pulse generator and dynamics of luteinizing hormone pulses in the rhesus monkey. Proc Natl Acad Sci USA 1990; 87: 8580–2.10.1073/pnas.87.21.8580Suche in Google Scholar PubMed PubMed Central
113. Goubillon ML, Forsdike RA, Robinson JE, Ciofi P, Caraty A, Herbison AE. Identification of neurokinin B-expressing neurons as an highly estrogen-receptive, sexually dimorphic cell group in the ovine arcuate nucleus. Endocrinology 2000; 141: 4218–25.10.1210/endo.141.11.7743Suche in Google Scholar PubMed
114. Navarro VM, Gottsch ML, Wu M, Garcia-Galiano D, Hobbs SJ, Bosch MA, Pinilla L, Clifton DK, Dearth A, Ronnekleiv OK, Braun RE, Palmiter RD, Tena-Sempere M, Alreja M, Steiner RA. Regulation of NKB pathways and their roles in the control of Kiss1 neurons in the arcuate nucleus of the male mouse. Endocrinology 2011; 152: 4265–75.10.1210/en.2011-1143Suche in Google Scholar PubMed PubMed Central
115. Navarro VM, Castellano JM, McConkey SM, Pineda R, Ruiz-Pino F, Pinilla L, Clifton DK, Tena-Sempere M, Steiner RA. Interactions between kisspeptin and neurokinin B in the control of GnRH secretion in the female rat. Am J Physiol Endocrinol Metab 2011; 300: E202–10.10.1152/ajpendo.00517.2010Suche in Google Scholar PubMed PubMed Central
116. Billings HJ, Connors JM, Altman SN, Hileman SM, Holaskova I, Lehman MN, McManus CJ, Nestor CC, Jacobs BH, Goodman RL. Neurokinin B acts via the neurokinin-3 receptor in the retrochiasmatic area to stimulate luteinizing hormone secretion in sheep. Endocrinology 2010; 151: 3836–46.10.1210/en.2010-0174Suche in Google Scholar PubMed PubMed Central
117. Ramaswamy S, Seminara SB, Ali B, Ciofi P, Amin NA, Plant TM. Neurokinin B stimulates GnRH release in the male monkey (Macaca mulatta) and is colocalized with kisspeptin in the arcuate nucleus. Endocrinology 2010; 151: 4494–503.10.1210/en.2010-0223Suche in Google Scholar PubMed PubMed Central
118. Ohtaki T, Shintani Y, Honda S, Matsumoto H, Hori A, Kanehashi K, Terao Y, Kumano S, Takatsu Y, Masuda Y, Ishibashi Y, Watanabe T, Asada M, Yamada T, Suenaga M, Kitada C, Usuki S, Kurokawa T, Onda H, Nishimura O, Fujino M. Metastasis suppressor gene KiSS-1 encodes peptide ligand of a G-protein-coupled receptor. Nature 2001; 411: 613–7.10.1038/35079135Suche in Google Scholar PubMed
119. Seminara SB, Messager S, Chatzidaki EE, Thresher RR, Acierno JS Jr., Shagoury JK, Bo-Abbas Y, Kuohung W, Schwinof KM, Hendrick AG, Zahn D, Dixon J, Kaiser UB, Slaugenhaupt SA, Gusella JF, O’Rahilly S, Carlton MB, Crowley WF Jr., Aparicio SA, Colledge WH. The GPR54 gene as a regulator of puberty. N Engl J Med 2003; 349: 1614–27.10.1056/NEJMoa035322Suche in Google Scholar PubMed
120. Gottsch ML, Cunningham MJ, Smith JT, Popa SM, Acohido BV, Crowley WF, Seminara S, Clifton DK, Steiner RA. A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology 2004; 145: 4073–7.10.1210/en.2004-0431Suche in Google Scholar PubMed
121. de Roux N. GnRH receptor and GPR54 inactivation in isolated gonadotropic deficiency. Best Pract Res Clin Endocrinol Metab 2006; 20: 515–28.10.1016/j.beem.2006.10.005Suche in Google Scholar PubMed
122. Burke MC, Letts PA, Krajewski SJ, Rance NE. Coexpression of dynorphin and neurokinin B immunoreactivity in the rat hypothalamus: morphologic evidence of interrelated function within the arcuate nucleus. J Comp Neurol 2006; 498: 712–26.10.1002/cne.21086Suche in Google Scholar PubMed
123. Foradori CD, Amstalden M, Goodman RL, Lehman MN. Colocalisation of dynorphin a and neurokinin B immunoreactivity in the arcuate nucleus and median eminence of the sheep. J Neuroendocrinol 2006; 18: 534–41.10.1111/j.1365-2826.2006.01445.xSuche in Google Scholar PubMed
124. Goodman RL, Lehman MN, Smith JT, Coolen LM, de Oliveira CV, Jafarzadehshirazi MR, Pereira A, Iqbal J, Caraty A, Ciofi P, Clarke IJ. Kisspeptin neurons in the arcuate nucleus of the ewe express both dynorphin A and neurokinin B. Endocrinology 2007; 148: 5752–60.10.1210/en.2007-0961Suche in Google Scholar PubMed
125. Lehman MN, Coolen LM, Goodman RL. Minireview: kisspeptin/neurokinin B/dynorphin (KNDy) cells of the arcuate nucleus: a central node in the control of gonadotropin-releasing hormone secretion. Endocrinology 2010; 151: 3479–89.10.1210/en.2010-0022Suche in Google Scholar PubMed PubMed Central
126. Amstalden M, Coolen LM, Hemmerle AM, Billings HJ, Connors JM, Goodman RL, Lehman MN. Neurokinin 3 receptor immunoreactivity in the septal region, preoptic area and hypothalamus of the female sheep: colocalisation in neurokinin B cells of the arcuate nucleus but not in gonadotrophin-releasing hormone neurones. J Neuroendocrinol 2010; 22: 1–12.10.1111/j.1365-2826.2009.01930.xSuche in Google Scholar PubMed PubMed Central
127. Navarro VM, Gottsch ML, Chavkin C, Okamura H, Clifton DK, Steiner RA. Regulation of gonadotropin-releasing hormone secretion by kisspeptin/dynorphin/neurokinin B neurons in the arcuate nucleus of the mouse. J Neurosci 2009; 29: 11859–66.10.1523/JNEUROSCI.1569-09.2009Suche in Google Scholar PubMed PubMed Central
128. Topaloglu AK, Reimann F, Guclu M, Yalin AS, Kotan LD, Porter KM, Serin A, Mungan NO, Cook JR, Ozbek MN, Imamoglu S, Akalin NS, Yuksel B, O’Rahilly S, Semple RK. TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for Neurokinin B in the central control of reproduction. Nat Genet 2009; 41: 354–8.10.1038/ng.306Suche in Google Scholar PubMed PubMed Central
129. Goodman RL, Coolen LM, Anderson GM, Hardy SL, Valent M, Connors JM, Fitzgerald ME, Lehman MN. Evidence that dynorphin plays a major role in mediating progesterone negative feedback on gonadotropin-releasing hormone neurons in sheep. Endocrinology 2004; 145: 2959–67.10.1210/en.2003-1305Suche in Google Scholar PubMed
130. Wakabayashi Y, Nakada T, Murata K, Ohkura S, Mogi K, Navarro VM, Clifton DK, Mori Y, Tsukamura H, Maeda K, Steiner RA, Okamura H. Neurokinin B and dynorphin A in kisspeptin neurons of the arcuate nucleus participate in generation of periodic oscillation of neural activity driving pulsatile gonadotropin-releasing hormone secretion in the goat. J Neurosci 2010; 30: 3124–32.10.1523/JNEUROSCI.5848-09.2010Suche in Google Scholar PubMed PubMed Central
131. Goodman RL, Coolen LM, Lehman MN. A role for neurokinin B in pulsatile GnRH secretion in the ewe. Neuroendocrinology 2013. DOI:10.1159/000355285.10.1159/000355285Suche in Google Scholar PubMed PubMed Central
132. de Croft S, Boehm U, Herbison AE. Neurokinin B activates arcuate kisspeptin neurons through multiple tachykinin receptors in the male mouse. Endocrinology 2013; 154: 2750–60.10.1210/en.2013-1231Suche in Google Scholar PubMed
133. Clarkson J, Herbison AE. Oestrogen, kisspeptin, GPR54 and the pre-ovulatory luteinising hormone surge. J Neuroendocrinol 2009; 21: 305–11.10.1111/j.1365-2826.2009.01835.xSuche in Google Scholar PubMed
134. Foradori CD, Coolen LM, Fitzgerald ME, Skinner DC, Goodman RL, Lehman MN. Colocalization of progesterone receptors in parvicellular dynorphin neurons of the ovine preoptic area and hypothalamus. Endocrinology 2002; 143: 4366–74.10.1210/en.2002-220586Suche in Google Scholar PubMed
135. Cheng G, Coolen LM, Padmanabhan V, Goodman RL, Lehman MN. The kisspeptin/neurokinin B/dynorphin (KNDy) cell population of the arcuate nucleus: sex differences and effects of prenatal testosterone in sheep. Endocrinology 2010; 151: 301–11.10.1210/en.2009-0541Suche in Google Scholar PubMed PubMed Central
136. Ruiz-Pino F, Navarro VM, Bentsen AH, Garcia-Galiano D, Sanchez-Garrido MA, Ciofi P, Steiner RA, Mikkelsen JD, Pinilla L, Tena-Sempere M. Neurokinin B and the control of the gonadotropic axis in the rat: developmental changes, sexual dimorphism, and regulation by gonadal steroids. Endocrinology 2012; 153: 4818–29.10.1210/en.2012-1287Suche in Google Scholar PubMed PubMed Central
137. Kinsey-Jones JS, Grachev P, Li XF, Lin YS, Milligan SR, Lightman SL, O’Byrne KT. The inhibitory effects of neurokinin B on GnRH pulse generator frequency in the female rat. Endocrinology 2012; 153: 307–15.10.1210/en.2011-1641Suche in Google Scholar PubMed
138. Garcia-Galiano D, van Ingen Schenau D, Leon S, Krajnc-Franken MA, Manfredi-Lozano M, Romero-Ruiz A, Navarro VM, Gaytan F, van Noort PI, Pinilla L, Blomenrohr M, Tena-Sempere M. Kisspeptin signaling is indispensable for neurokinin B, but not glutamate, stimulation of gonadotropin secretion in mice. Endocrinology 2012; 153: 316–28.10.1210/en.2011-1260Suche in Google Scholar PubMed
139. Grachev P, Li XF, Kinsey-Jones JS, di Domenico AL, Millar RP, Lightman SL, O’Byrne KT. Suppression of the GnRH pulse generator by neurokinin B involves a kappa-opioid receptor-dependent mechanism. Endocrinology 2012; 153: 4894–904.10.1210/en.2012-1574Suche in Google Scholar PubMed
140. Clarkson J, Shamas S, Mallinson S, Herbison AE. Gonadal steroid induction of kisspeptin peptide expression in the rostral periventricular area of the third ventricle during postnatal development in the male mouse. J Neuroendocrinol 2012; 24: 907–15.10.1111/j.1365-2826.2012.02294.xSuche in Google Scholar PubMed
141. Gill JC, Navarro VM, Kwong C, Noel SD, Martin C, Xu S, Clifton DK, Carroll RS, Steiner RA, Kaiser UB. Increased neurokinin B (Tac2) expression in the mouse arcuate nucleus is an early marker of pubertal onset with differential sensitivity to sex steroid-negative feedback than Kiss1. Endocrinology 2012; 153: 4883–93.10.1210/en.2012-1529Suche in Google Scholar PubMed PubMed Central
142. Mayer C, Boehm U. Female reproductive maturation in the absence of kisspeptin/GPR54 signaling. Nat Neurosci 2011; 14: 704–10.10.1038/nn.2818Suche in Google Scholar
143. Caron E, Ciofi P, Prevot V, Bouret SG. Alteration in neonatal nutrition causes perturbations in hypothalamic neural circuits controlling reproductive function. J Neurosci 2012; 32: 11486–94.10.1523/JNEUROSCI.6074-11.2012Suche in Google Scholar
144. De Bond JA, Li Q, Millar RP, Clarke IJ, Smith JT. Kisspeptin signaling is required for the luteinizing hormone response in anestrous ewes following the introduction of males. PLoS One 2013; 8: e57972.10.1371/journal.pone.0057972Suche in Google Scholar
145. Covenas R, Martin F, Belda M, Smith V, Salinas P, Rivada E, Diaz-Cabiale Z, Narvaez JA, Marcos P, Tramu G, Gonzalez-Baron S. Mapping of neurokinin-like immunoreactivity in the human brainstem. BMC Neurosci 2003; 4: 3.10.1186/1471-2202-4-3Suche in Google Scholar
146. Cinelli E, Robertson B, Mutolo D, Grillner S, Pantaleo T, Bongianni F. Neuronal mechanisms of respiratory pattern generation are evolutionary conserved. J Neurosci 2013; 33: 9104–12.10.1523/JNEUROSCI.0299-13.2013Suche in Google Scholar
147. Bolser DC. Central mechanisms II: pharmacology of brainstem pathways. Handb Exp Pharmacol 2009; 187: 203–17.10.1007/978-3-540-79842-2_10Suche in Google Scholar
148. Canning BJ. Central regulation of the cough reflex: therapeutic implications. Pulm Pharmacol Ther 2009; 22: 75–81.10.1016/j.pupt.2009.01.003Suche in Google Scholar
149. Canning BJ, Mori N. Encoding of the cough reflex in anesthetized guinea pigs. Am J Physiol Regul Integr Comp Physiol 2011; 300: R369–77.10.1152/ajpregu.00044.2010Suche in Google Scholar
150. Merchenthaler I, Maderdrut JL, O’Harte F, Conlon JM. Localization of neurokinin B in the central nervous system of the rat. Peptides 1992; 13: 815–29.10.1016/0196-9781(92)90192-6Suche in Google Scholar
151. Duque E, Mangas A, Salinas P, Diaz-Cabiale Z, Narvaez JA, Covenas R. Mapping of alpha-neo-endorphin- and neurokinin B-immunoreactivity in the human brainstem. Brain Struct Funct 2013; 218: 131–49.10.1007/s00429-012-0388-3Suche in Google Scholar PubMed
152. Hether S, Misono K, Lessard A. The neurokinin-3 receptor (NK3R) antagonist SB222200 prevents the apomorphine-evoked surface but not nuclear NK3R redistribution in dopaminergic neurons of the rat ventral tegmental area. Neuroscience 2013; 247: 12–24.10.1016/j.neuroscience.2013.05.006Suche in Google Scholar PubMed
153. Flashner E, Raviv U, Friedler A. The effect of tachykinin neuropeptides on amyloid beta aggregation. Biochem Biophys Res Commun 2011; 407: 13–7.10.1016/j.bbrc.2011.02.067Suche in Google Scholar PubMed
154. Friedrich P, Feulner TM, Laws SM, Eckart K, Perneczky R, Kurz A, Forstl H, Riemenschneider M. No association of tachykinin receptor 2 (TACR2) polymorphisms with Alzheimer’s disease. Neurobiol Aging 2011; 32: 544–5.10.1016/j.neurobiolaging.2009.03.007Suche in Google Scholar PubMed
155. Horii A, Nakagawa A, Uno A, Kitahara T, Imai T, Nishiike S, Takeda N, Inohara H. Implication of substance P neuronal system in the amygdala as a possible mechanism for hypergravity-induced motion sickness. Brain Res 2012; 1435: 91–8.10.1016/j.brainres.2011.11.047Suche in Google Scholar PubMed
156. Rivera-Mancia S, Perez-Neri I, Rios C, Tristan-Lopez L, Rivera-Espinosa L, Montes S. The transition metals copper and iron in neurodegenerative diseases. Chem Biol Interact 2010; 186: 184–99.10.1016/j.cbi.2010.04.010Suche in Google Scholar PubMed
157. Russino D, McDonald E, Hejazi L, Hanson GR, Jones CE. The tachykinin peptide neurokinin B binds copper forming an unusual [Cu(NKB)] complex and inhibits copper uptake into 1321N1 astrocytoma cells. ACS Chem Neurosci 2013; 4: 1371–81.10.1021/cn4000988Suche in Google Scholar PubMed PubMed Central
158. Boll MC, Alcaraz-Zubeldia M, Montes S, Rios C. Free copper, ferroxidase and SOD1 activities, lipid peroxidation and NO(x) content in the CSF. A different marker profile in four neurodegenerative diseases. Neurochem Res 2008; 33: 1717–23.10.1007/s11064-008-9610-3Suche in Google Scholar PubMed
159. Blaszak M, Jankowska E, Kowalik-Jankowska T. Acid-base properties of the (1–4,18–36) fragments of neuropeptide K and their mono- and polynuclear copper(II) complexes products of metal-catalyzed oxidation. Inorg Chem 2013; 52: 130–43.10.1021/ic301476pSuche in Google Scholar PubMed
160. Arai H, Emson PC, Carrasco LH. Huntington’s disease: changes in tachykinin content in postmortem brains. Ann Neurol 1987; 22: 587–94.10.1002/ana.410220505Suche in Google Scholar PubMed
161. Kowalik-Jankowska T, Jankowska E, Kasprzykowski F. Coordination abilities of a fragment containing D1 and H12 residues of neuropeptide gamma and products of metal-catalyzed oxidation. Inorg Chem 2010; 49: 2182–92.10.1021/ic902021jSuche in Google Scholar PubMed
©2014 by Walter de Gruyter Berlin/Boston
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Artikel in diesem Heft
- Frontmatter
- Review
- Mammalian genome evolution as a result of epigenetic regulation of transposable elements
- Neuroglobin – recent developments
- The Jumonji family: past, present and future of histone demethylases in cancer
- Tachykinin: recent developments and novel roles in health and disease
- Circadian angiogenesis
- Short Conceptual Overview
- Transthyretin microheterogeneity and molecular interactions: implications for amyloid formation
Artikel in diesem Heft
- Frontmatter
- Review
- Mammalian genome evolution as a result of epigenetic regulation of transposable elements
- Neuroglobin – recent developments
- The Jumonji family: past, present and future of histone demethylases in cancer
- Tachykinin: recent developments and novel roles in health and disease
- Circadian angiogenesis
- Short Conceptual Overview
- Transthyretin microheterogeneity and molecular interactions: implications for amyloid formation