Significance of bacteriophages in fermented soybeans: A review
-
Ekachai Chukeatirote
 ,
Wallapat Phongtang
,
Wallapat Phongtang

Abstract
Bacteriophages are ubiquitous and have been reported to have been found in many food products. Their presence is important as they have the ability to interact with their bacterial host in food matrices. Fermented soybean products, one of the most widely consumed ethnic foods among Asian people, are prepared naturally and include Japanese Natto, Indian Kinema, Korean Chongkukjang and Thai Thua Nao. This review highlights bacteriophages which have been isolated from fermented soybean products and also includes an overview of their diversity, occurrence as well as their significance.
Introduction
Fermented foods are defined as those which have been processed by microbial activity. In general, fermented food products can be classified on the basis of the raw materials used (i.e. beverages, dairy, fish or meat or fruit and vegetable products) (1). Well known fermented foods which have been popularly consumed include fermented dairy products and fermented sausages. Others may be considered ‘indigenous’ and consumed only by particular consumer groups; these include fermented soybeans and fish sauce. According to the definition, fermented foods must be produced by microbes (as well as their enzymes), it is therefore not surprising that fermented food products are a reliable reservoir for microbiota, including bacteriophages (especially for those products produced by the action of bacteria).
Bacteriophages or phages, are viruses which can infect bacteria (Figure 1A). They are ubiquitous and may be the most abundant entities on Earth (2). Once denied, it is now widely accepted that phages have played a major role in the regulation of bacterial communities in every ecosystem (3, 4). Phages can interact with their host by either the lytic pathway or lysogenic mode. Lytic phages are known to disrupt bacterial metabolism and result in cell lysis. In contrast, lysogenic phages (also known as temperate phages) can integrate their DNA into the host genome, replicate together, and thus establish a longterm relationship with their bacterial host (Figure 1B) (5). Due to their simple structure and host specificity, bacteriophages have played a pivotal role in the field of molecular biology and biotechnology. Many biological concepts can be resolved by the use of bacteriophages, for example the Hershey–Chase experiment (6). More recently, bacteriophages have captured interest among the scientific community specifically for phage therapy, phage display systems, vaccine delivery vehicles and even for diagnostic use (7).
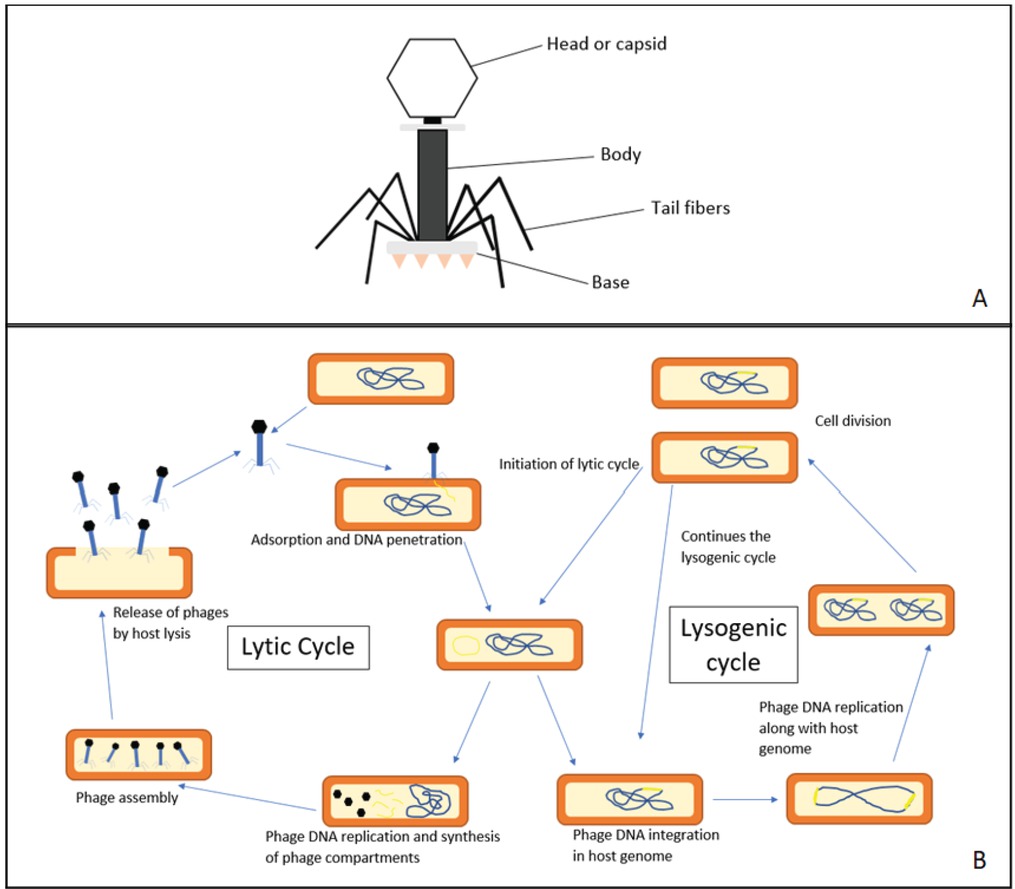
Bacteriophage and its life cycle. (A) The structure of a tailed bacteriophage; (B) The phage life cycle: lytic and lysogenic cycle.
Phages have also contributed considerably to the food industry. Many phages have been isolated and studied from various food products. Table 1 gives examples of fermented food products in which bacteriophages were isolated and studied. Based on different food substrates, the abundance of phages has been demonstrated in a wide range of fermented food products. Most bacteriophages commonly isolated and studied to date are specific to lactic acid bacteria (LAB) (23). It should be noted however, that other bacteriophages infecting other bacterial groups are also present and remain to be studied, for example the bacteriophage groups present in fermented foods not associated with LAB. In general, the presence of phages appears to be detrimental to bacterial starter cultures used and often results in food production failure (24). On the other hand, phage infecting bacterial pathogens can be considered beneficial, as many have
Fermented food products and their associated bacteriophages.
| Fermented foods | Bacterial host | Phage effect | References |
|---|---|---|---|
| Dairy-based | |||
| - Cheese | LAB | Lytic activity to LAB strains and commercial starter culture | 8 - 11 |
| - Yoghurt | S. thermophilus L. delbrueckii | Lytic activity and temperate phage | 12 - 14 |
| Vegetable-based | |||
| - Fermenting cucumber | LAB | Lytic phage against commercial starter culture | 15 - 17 |
| - Kimchi | LAB Weissella cibaria | Lytic activity to LAB population | 18, 19 |
| - Sauekraut Animal-based | LAB | Lytic activity to starter culture | 20 |
| - Fermented sausages | LAB | Lytic activity to starter culture | 21 |
| - Nham (fermented pork) | Weissella cibaria | Lytic activity | 22 |
Notes: LAB = Lactic acid bacteria; S. = Streptococcus; L. = Lactobacillus.
reported the ability to control foodborne pathogenic bacteria (25, 26). Fermented soybeans (FSB) are another type of fermented food for which soybeans are used as the raw material. Interestingly, several studies have shown that the predominant bacterial group of this product is Bacillus species, and not LAB (1, 27). With only a few studies having been performed to date regarding phages infecting the Bacillus species in fermented soybean products, the aim of this review was to increase awareness of phage occurrence, and its potential role and significance during the soybean fermentation process.
Fermented soybeans (FSB)
Soybeans [Glycine max (L.) Merrill] belong to the family Leguminosae and are one of the most important crops worldwide (28). Soybean seeds are protein-rich and contain many beneficial nutrients, however in order for human consumption, the soybeans must be cooked to destroy anti-nutritional factors such as trypsin inhibitor and hemaglutinins (lectins) (29). The soybeans can then be safely consumed or further processed into various products. The fermentation of soybeans is a process which results in the formation of novel foods products (fermented soybeans) with benefits including prolonging the shelf life.
There are many kinds of FSB products including Japanese Natto, Korean Chongkukjang, Indian Kinema and Thai Thua Nao (30). Figure 2 shows representative images of Thai Thua Nao, FSB products. In addition, detailed characteristics of these FSB products are provided in Table 2. To perform the fermentation process, three major ingredients are used: soybeans, water and naturally occurring bacteria. The fermentation process in brief consists of soaking, boiling, fermenting and post-fermenting (27). The procedure typically starts with the soaking and boiling of soybean seeds for 3-4 h until soft. The cooked soybeans are then placed in a lined bamboo basket and covered with banana leaves. The soybean fermentation occurs naturally at ambient temperatures for 2-3 days. Post-fermentation processes have been developed by locals with the aim of prolonging the products shelf-life (Figure 3).

Representative images of Thai Thua Nao fermented soybean (FSB) products. (A - B) Fresh Thua Nao products. (C - D) Dried Thua Nao products.
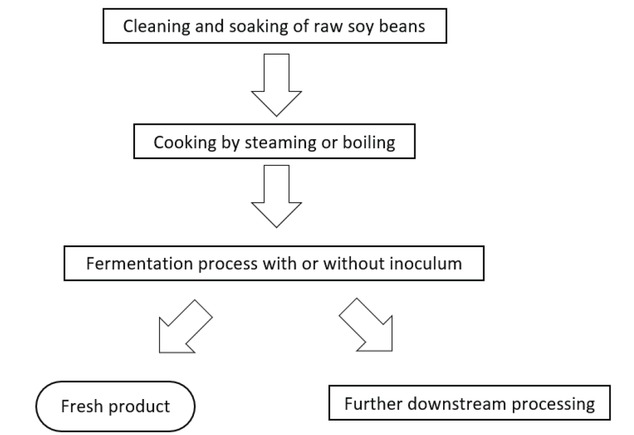
The typical production process of fermented soybean (FSB) products.
Fermented soybean products and their characteristics.
| Products / Country | Uses | Isolated bacteria | References |
|---|---|---|---|
| Natto / Japan | Directly eaten; flavor enhancer | Bacillus subtilis (natto) | 31 |
| Cheonggukjang / Korea | Flavor enhancer | B. subtilis, B. cereus | 32, 33 |
| Kinema / India | Main dish | B. subtilis, B. licheniformis, | 34, 35 |
| B. cereus, B. circulans, | |||
| B. thuringensis, B. sphaericus, Enterococcus | |||
| faecum, Escherichia coli and other | |||
| Enterobaceriaceae | |||
| Thua Nao / Thailand | Flavor enhancer | B. subtilis, B. licheniformis, | 36, 37 |
| B. pumilus, B. megaterium, | |||
| B. cereus, Lactobacillus sp., | |||
| Gram-positive cocci |
The majority of FSB products are traditionally produced, using naturally occurring microbes. Natto is one of the best-known examples and is prepared using a pure starter culture of B. subtilis strain natto, for commercial purposes (31). The Bacillus species are predominant in FSB products and thus considered to play a key role during fermentation (38, 39). Other bacterial strains have also been detected, although their presence is often considered a ‘contaminant’ as they do not appear to play a role in the fermentation process (27).
Historical study of bacteriophages in relation to FSB products
Since their discovery by Twort (40) and d’Herelle (41), bacteriophages have been isolated and studied by many and are of great interest due to their simple structure, useful not only for fundamental biological studies but also for their potential use in biotechnological applications (42, 43). In terms of food-related bacteriophages, their study can be traced back to 1946, where they were used for staphylococci typing (44, 45).
The bacteriophages related to FSB products were initially isolated from Natto; where the presence of phages resulted in the abnormal production of Natto whereby the viscous polymer polyglutamic acid (PGA) was absent (46). Natto products affected by phages, were then considered ‘spoiled’ and lead to a tremendous loss to the Natto manufacturing business. Since 1967, many bacteriophages infecting Natto bacterial starter cultures have been isolated and characterized, and include the work of Yoshimoto and Hongo (47), Yoshimoto et al. (48), Fujii et al. (49, 50), Yamamoto (51), Tsutsumi et al. (52), Nagai and Yamasaki (53) and Umene et al. (54). In addition, further research relating to the influence of phages on PGA degradation have been performed and include the work of Hara et al. (55), Hongo and Yoshimoto (56-58), Nagai and Itoh (59), Kimura and Itoh (60), Kimura (61), Fujimoto et al. (62) and Ozaki et al. (63). It should be noted however, that although most references relating to the isolation and characterization of Natto phages (between 1967 and 1986) were written in Japanese, there are two excellent English reviews related to Natto bacteriophages (64, 65). Based on these studies, a report describing guidelines to improve the fermentation procedure of Natto was proposed (66).
Subsequent research on FSB-associated phages has been performed on Korean Cheonggukjang (or Jang). The first record detailing phage isolation from Korean soybean paste came from the work of Lee in 1978 (67). Lee reported that this phage could infect B. subtilis var. 816, and thus result in fermentation failure. However, there has been no further work on this topic until 2011 by Korean scientists and to date, there have been only seven research papers on this topic (between 2011 and 2018). In contrast, the bacteriophage findings have been related to Bacillus, normal flora strains in Cheonggukjang products. This was expected, as no specific inoculum is required for Cheonggukjang fermentation. Cheonggukjang and other FSB products (except Natto) are typically produced in households using naturally occurring bacterial cultures. Much work has focused on phages infecting B. cereus and aim to employ them as biocontrol agents (32, 68 - 71). The bacteriophages specific to B. subtilis have been found (72) and confirmed to play a detrimental role in the quality of resulting Cheonggukjang products (73). For other FSB products, there are currently no reports describing phages. Figure 4 shows a timeline of the major studies associated with phages isolated from FSB products.
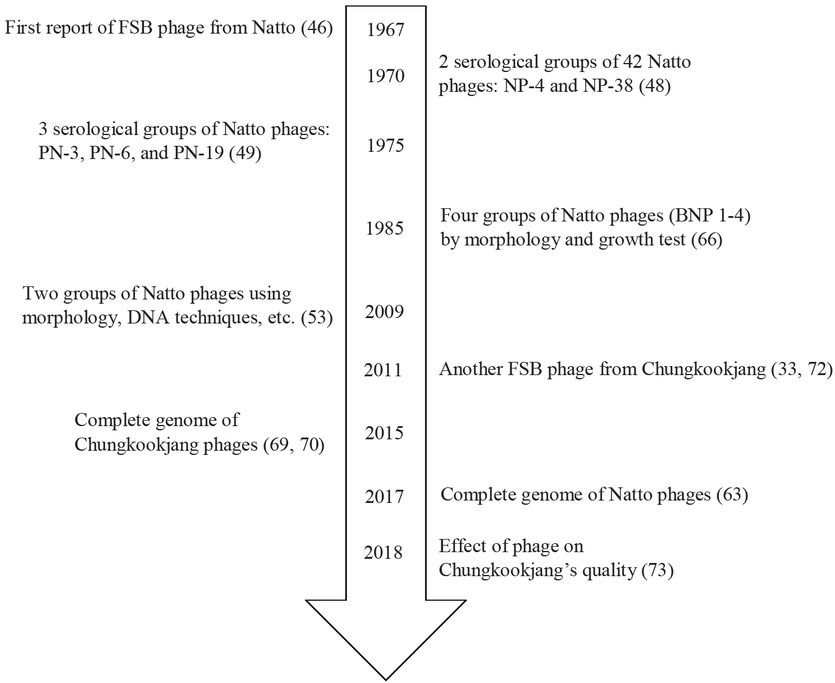
Major events relating to fermented soybean (FSB) phage research. References shown in brackets.
Classification and occurrence of fermented soybean bacteriophages: an overview
One of the distinct features of Natto is the presence of a mucous slime substance known as polyglutamic acid (PGA) (74). Notably, Natto affected by phages fail to produce PGA and it was this observation that initiated a focus on the presence of phages in FSB products. In 1967, Fujii et al. reported on the isolation of a phage known as PN-1 infecting the B. subtilis (natto) starter cultural strain from an abnormal Natto product (reduced or no PGA present). From there, many Japanese groups found further evidence of Natto phages from such products, as well as Natto factories and have studied their characteristics (64, 65).
During this period, isolated Natto phages were screened and classified by a serological method. Work by several research groups resulted in the classification of Natto phages into between 2 and 4 groups (46, 48). There has been no further studies describing the relationship of these phages at this time. The phages were also named differently. It was not until 2009, after a detailed study of the Natto bacteriophages was performed by Nagai and Yamasaki, that a reclassification occurred. The Natto phages were analysed using a DNA hybridization technique, and their morphology uncovered, as well as other physicochemical analyses. Based on this data, many Natto phages previously isolated were then grouped into two types: Group 1 (represented by the JNDMP isolate) and Group 2 (represented by the ONPA isolate) (53). Key features of the two Natto phage groups are summarised in Table 3. In addition, based on the phage PM1 genome, a specific primer set was developed, designed for the selective amplification of a 0.53kb DNA region of the 1.1kb EcoRI fragment of the PM1 DNA (54). The existence of this amplified product, was distinct and strongly associated with the PM1 and PM1-relaed phages, and proved useful for the detection and distribution of FSB phages. It is expected that this finding may be useful in the FSB production pipeline (such as in the Natto’s factory) to prevent and control phage infection (54).
Key characteristics of the two Natto phage groups (adapted from 64, 65).
| Group 1 (JNDMP) | Group 2 (ONPA) | |
|---|---|---|
| Head (diameter, nm) | 60 | 89 |
| Tail (width x length, nm) | 7 x 200 | 9 x 200 |
| Sheath (width, nm) | Absent | 23 |
| Genomic size (kb) | 42 | 91 |
| Latent time (min) | 35 | 50 |
| Burst size | 46 | 72 |
| Heat stability (oC) | 53 | 63 |
| Mg2+ requirement | Required | Not required |
| Host range | ||
| - B. subtilis (natto) | ||
| - B. subtilis Marburg | ||
| - B. cereus | ||
| - B. brevis | ||
| - B. megaterium |
At present, bacteriophage nomenclature and classification are governed by the International Committee on Taxonomy of Viruses (ICTV). Many criteria have been proposed for phage classification, including nucleic acid nature, particle structure and nucleotide or amino acid sequences (75), although it should be noted that viral morphology observed by electron microscopy remains important (75). Based on this aspect, phages are either tailed, polyhedral, filamentous or pleomorphic. Of these structures, tailed phages (Order Caudovirales) constitute the largest, most widespread group of bacterial viruses (ca. 96%) and can be classified into three families: Myoviridae (with contractile tail), Siphoviridae (with long, non-contractile tail), and Podoviridae (with extremely short, non-contractile tail) (75). A diagram showing the morphological structures of these three tailed phage families is presented in Figure 5. All phages isolated from the FSB products to date have been tailed phages, grouped into Myoviridae or Siphoviridae. For the two Natto phage groups, Group 1 is Siphoviridae, and Group 2 is Myoviridae (Figure 5 and Table 4).
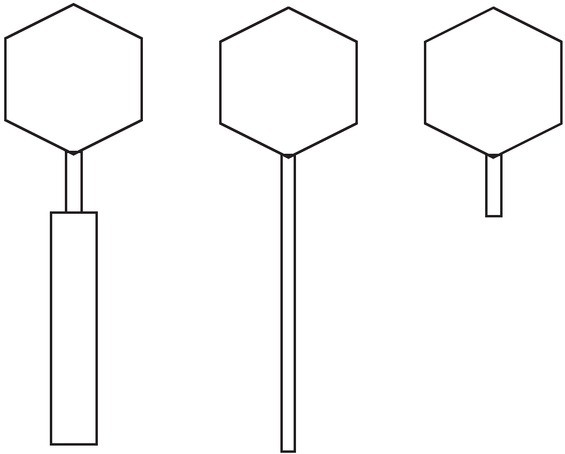
A diagram to show the three most common tailed bacteriophage morphotypes: Myoviridae (left), Siphoviridae (middle), and Podoviridae (right).
Other studies of FSB phages have come from Cheonggukjang products. In 2011, the phages infecting B. subtilis and B. cereus have been isolated from Korean Cheonggukjang (33, 72). Since then, further work regarding this topic has been extended and most studies have focused on phages infecting B. cereus, aiming to use these phages as a biocontrol agent for controlling foodborne pathogenic bacteria (33, 68, 71). Recently, Ghosh et al. (73) has shown that the phages infecting B. subtilis are abundant in Cheonggukjang products and may cause detrimental effect to the fermentation process, as well as the products quality. The majority of phages isolated from Cheonggukjang products are Myoviridae and only the phages reported by Oh et al. (71) are Siphoviridae (Table 4).
Detailed characteristics of FSB-related phages.
| FSB-related phages | Host | Family | Genome size | Morphology | References | |
|---|---|---|---|---|---|---|
| Natto | ||||||
| - JNDMP | BS | S | 42 kb | Hexagonal head (diameter, 60 nm) | 53 | |
| Non-contractile tail (7 x 200 nm) | ||||||
| - ONPA | BS | M | 91 kb | Hexagonal head (diameter, 89 nm) | 53 | |
| Contractile tail (9 x 200 nm) with a sheath (23 nm) | ||||||
| - ϕNIT1 | BS | M | 155,631 bp | Isometric head (100nm in diameter) | 63 | |
| Contractile tail (250 nm long) | ||||||
| - PM1 | BS | S | 50 kb | A long, non-contracted tail virus (similar to JNDMP) | 54 | |
| - BN100 | BS | S | 42 kb | Hexagonal head (60 by 67 nm) | 59 | |
| Non-contractile tail (7 x 200 nm) (similar to JNDMP) | ||||||
| - PBND8 | BS | M | ND | Hexagonal head (diameter, 39 nm) | 52 | |
| Contractile tail | ||||||
| Cheonggukjang - 816 phage | BS | ND | ND | Hexagonal head (diameter, 160 - 240 nm) | 67 | |
| - JBP901 | BC | M | 159, 492 bp | Hexagonal head (diameter, 95 nm) | 33, 69 | |
| Contractile tail (170 nm long) | 33, 69 | |||||
| - BCP1-1 | BC | M | 150 kb | Icosahedral head (diameter, 95 nm) | 68 | |
| Contractile tail (220 nm long) | ||||||
| - BCP8-2 | BC | M | 159,071 bp | Similar to BCP1-1, with 210 nm long contractile tail | 68, 70 | |
| - Bp-K2 | BS | M | 21 kb | Icosahedral head (50 by 80 nm) | 72 | |
| Contractile tail (85 - 90 nm long) with a basal plate | ||||||
| - BSP18 | BS | M | ND | Isometric head (90 nm in diameter) | 73 | |
| Contractile tail (200 nm long) | ||||||
| - CAU series | BC | S | ND | Hexagonal head (100 nm in diameter) | 71 | |
| Non-contractile tail (197 nm long, 11 nm wide) | ||||||
| Soy sauce - ϕD10 | TH | S | ND | Hexagonal head (55 nm in diameter) | 76 | |
| Non-contractile tail (200 nm long, 9 nm wide) | ||||||
Notes: BS = Bacillus subtilis; BC = B. cereus; TH = Tetragenococcus halophila; S = Siphoviridae; M = Myoviridae; ND = No data.
Significance of phages in soybean fermentation and the FSB products
Phages are abundant, found in both natural and man-made environments. This includes the fermentation of soybeans which provides a unique niche for the Bacillus-phages. As mentioned above, many phages related to the Bacillus bacterial group can be isolated during the soybean fermentation process, where a particular group of Bacillus species is present. The occurrence of phages is usually considered detrimental as they have the potential to cause failure (or delay) in food fermentation, ultimately leading to poor quality products. However, phages found naturally in food can also represent an alternative means to combat foodborne pathogenic bacteria—a major concern within the food industry. Similarly, significance of the phages found during soybean fermentation and within FSB products can be viewed both positively and negatively, and this will be discussed below.
Degradation risk to FSB products
From the beginning, phages within FSB products have been isolated from failed soybean fermentations or from poor quality FSB products. In 1967, Fujii et al. reported the first isolation of bacteriophages from abnormal Natto (i.e. due to the absence of the viscous biopolymer). These abnormal Natto products were considered ‘spoiled’ and thus discarded, a major problem for the Natto production industry. During 1960 - 1980, several Japanese researchers used abnormal Natto as a major source for the isolation and screening of phages (46, 48, 49). It was shown that these isolated phages could infect B. subtilis strain natto, an inoculum used for Natto preparation. The Natto production process relies on the metabolic capabilities of the B. subtilis starter culture, which could then be delayed, or even worse disrupted, resulting in poor quality Natto products.
One of the best-known examples of the phage effect on Natto products is the degradation of γ-polyglutamic acid (γ-PGA). PGA is a water-soluble, biodegradable substance comprising of D- and L-glutamic acids polymerized via γ-glutamyl bonds (77). In general, PGA is produced predominantly by bacteria belonging to Bacillus species, such as B. licheniformis, B. subtilis, B. megaterium, B. pumilis, B. mojavensis and B. amyloliquefaciens. It is worth mentioning that at least one Gram-negative bacterium, Fusobacterium nucleatum, and some archaea are also able to produce the PGA (78). PGA biosynthesis is a multi-step reaction occurring via a cytoplasmic membrane-bound enzyme complex (the CapABC proteins) using L-glutamic acid as a substrate (77). Physiological functions of PGA in these bacteria have been proposed; potentially as a barrier to attack via phages, or perhaps as a virulence factor (77). For FSB products, some PGA-producing Bacillus strains have been isolated from Natto, Doenjang and Kinema (54-63, 79-81). It should be noted however, that the ability to produce PGA is strain-specific, for example B. subtilis strain natto can produce PGA, whereas B. subtilis Marburg cannot (64). The presence of PGA is one of the key characteristics of Natto generating its sticky appearance. Although there is no reported range for optimum PGA quantities, it is generally acknowledged that the higher the PGA content, the higher the products quality (especially for Natto products) (59, 64, 65).
PGA degradation in Natto was initially described by Fujii et al. (46), followed by isolation of the phage culprit. It has been shown that B. subtilis strain natto infected with the phage was not able to accumulate PGA and resulted in ‘spoiled’ Natto. Further research revealed that this phage could produce extracellular PGA depolymerase enzyme which digested the PGA into di- and tri-γ-glutamate, and hence a reduction in viscosity (57, 58). Subsequently, Kimura and Itoh reported another PGA-degrading enzyme, the so-called ‘PghP’, from the phage ΦNIT1 (60). The PghP (γ-PGA hydrolase of phage) is a monomeric enzyme of 22.9 kDa able to digest the PGA to tri-, tetra-, and penta-γ-glutamate, suggesting that the PghP enzyme catalyzes the PGA differently from the PGA depolymerase of Hongo et al. (58). Unfortunately, Hongo’s enzyme has not been purified or studied further. The PghP work also showed that the phage ΦNIT1 has an apparent advantage over a phage that did not produce PghP when infecting the PGA-forming Bacillus cells (60). This finding supports the possible role of PGA as a host barrier, preventing the host cells from phage infection. Additionally, the PghP has been crystallized to determine its 3D-structure, in which the enzyme was found to belong to space group P3121 or P3221 (62).
Apart from the phage’s effect on PGA hydrolysis, a study of the phage ΦNIT1 genome revealed a putative levanase gene (orf477) encoding a possible levansucrase (63). Levan, a fructan polysaccharide that has also been found in FSB products as another biopolymer (82-84). Based on the comparative genomic analysis, the existence of the phgP and levP (orf477) was found in several Natto phages (including the ΦNIT1) (63). The role of the PghP enzyme produced by the ΦNIT1 phage as a strategy to infect the PGA-forming host cells was confirmed in this study (63). LevP was expected to degrade fructan, however, this was not clear from the phage growth and infection, and thus requires further investigation (63).
As a result, phage contamination poses a serious threat to the Natto production process, as sensitive B. subtilis host cells were lysed when encountering the phages. It has been hypothesized that the phages were brought into the factory and contaminated the subsequent fermentation process via the raw soybeans or dust in the air (64). Further investigations revealed that apart from abnormal Natto, the phages were also found in the factory and in the effluent (48, 49). It should also be noted that the phages were often isolated from the old factories where the factory walls were made of clay (an old style of Japanese building). As expected, the phages were found abundantly in the fermentation room where the bacterial starter cultures were inoculated (49). With this knowledge, a guideline to improve the Natto fermentation process was compiled and proposed by Nakajima (66) and Takiguchi et al. (89). Critical regulations to prevent phage contamination included i) the strict separation of the entrance and exit; ii) implementation of correct hand washing; iii) the immediate removal of abnormal Natto; iv) the use of sterilized water; v) UV installation; vi) the use of stainless steel equipment (no wooden tools); vii) regular cleaning of the factory; and viii) educating the workers about hygiene practices. Since the implication of these rules and regulations, phage detection has decreased enormously and in fact is rarely detected in the modern Natto factories (64).
Information regarding the effect of bacteriophages on other FSB products however, is limited. Apart from Natto, the only FSB phages known to date were isolated from Korean Cheonggukjang. The first report of phage presence and its effect on Korean soybean paste can be traced back to 1978 (67), although information related to this topic is minimal. Lee (67) reported a phage which could infect B. subtilis var. 816, a starter culture in manufacturing the soybean paste. This bacterial host-phage infection caused an occasional failure in soybean fermentation, subsequently leading to poor quality products (67). It was not until 2011, that Kim et al. (72) further described this event in which the phage Bp-K2 isolated from Chungkookjang exhibited lytic activity on various strains of B. subtilis. However, only the phages characteristics were determined with no report of its effect on soybean fermentation. Detailed information relating to this scope has recently been published in 2018, describing a similar phenomenon which was also found in Korean FSB products. Ghosh et al. (73) revealed that there was a high prevalence of phages infecting B. subtilis in Cheonggukjang. Phages in contact with bacterial host cells stall bacterial growth and yield products of poor quality (73).
Beneficial role as biocontrol agents
Among the naturally produced FSB products, currently Natto is the only product for which a pure starter culture of B. subtilis strain Natto is used in the production process (31). Due to Natto’s popularity among Japanese consumers, the production process of Natto has been strictly controlled to guarantee the products quality and safety. Therefore, the microbial contamination of Natto product is considered to be ‘minimal’. However, unlike Natto, other FSB products are conventionally produced by artisanal techniques. These FSB products can be considered ‘indigenous’ and are produced locally from knowledge passed from generation to generation. More importantly, their fermentation process occurs naturally by mixed microbial cultures, although many studies confirm that Bacillus species are predominant (27). As a result, these FSB production processes (except Natto) can be regarded as ‘non-sterile’. Microbial contamination, especially from pathogens can be expected and once developed generates a serious issue for consumers.
There are no specific regulations regarding the microbiological safety of FSB products, and varies from country to country. For example, in Thailand, there is only a guideline of ‘Community Product Standards’ for Thua Nao products, which does not include enough detail for microbiological analyses (85). Nevertheless, to ensure public safety, the FDA of Thailand has established guidelines related to microbiological contaminates of foods and related food products (86) in accordance with other international standards such as Codex and ICMSF (87, 88). Noted specifications to indicate acceptable quality of foods includes: i) total aerobic count of less than 4 log CFU/g; ii) Enterobacteriaceae count of less than 100 CFU/g; iii) Escherichia, Listeria, Vibrio, and Staphylococcus count of less than 20 CFU/g; and iv) pathogenic Bacillus including B. cereus count of less than 3 log CFU/g (88).
Of these FSB products (except Natto), there have been many reports describing events of microbial contamination. These include an occurrence of the Enterobacteriaceae members in Kinema (34), Escherichia coli, and B. cereus in Thua Nao (90), and B. cereus in Korean soybean paste (71). In these cases, the FSB-isolated phages can be acknowledged as ‘beneficial’ against such foodborne pathogenic bacteria. In terms of food safety, the presence of these allied phages is useful and once studied could be used as alternative means to eliminate these pathogens. To date, many research groups have focused on phages which exhibit lytic activity against B. cereus from FSB products. This is not surprising, considering that Bacillus species are widely found in these FSB foods. In particular, many FSB phages infecting B. cereus have been isolated from Korean Cheonggukjang (33, 68 - 71). These FSB phages specific to B. cereus exhibit strong lytic activity and are therefore a promising tool to tackle B. cereus contamination. Although most of the present studies have focused on the FSB phages of B. cereus, there are other contaminated bacterial strains which include E. coli, Salmonella and Staphylococcus (35, 90). Further work screening and isolating phages against these foodborne pathogenic bacteria is therefore of great importance when considering the impact on food safety.
Concluding remarks
Phages are versatile and can be found in all ecosystems where their bacterial hosts are present. In this review, we have focused on the occurrence of the phages relating to FSB products. The majority of phages isolated to date are associated with the Bacillus species. Although FSB phages are a viable risk factor to the bacterial starter cultures used in FSB preparations, they may offer a promising tool in combating foodborne pathogenic bacteria. Up until recently, FSB phages have only been isolated and studied from Natto and Cheonggukjang. It would be of great interest if FSB phages were isolated and studied from other FSB products. Further studies related to phage-host interactions are challenging, but important when considering that the starter culture could be selected or developed for optimal FSB production. In terms of food safety issues, some FSB phages against the foodborne pathogenic bacteria can be applied potentially as biocontrol agents.
Acknowledgements
The work in our laboratory was supported by Mae Fah Luang University. Additionally, this study was part of an academic collaboration between Mae Fah Luang University and Kangwon National University.
References
1 Steinkraus KH. Fermentation in world food processing. Compr Rev Food Sci Food Safety. 2002;1:23-32.10.1111/j.1541-4337.2002.tb00004.xSearch in Google Scholar
2 Clokie MRJ, Millard AD, Letarov AV and Heaphy S. Phages in nature. Bacteriophage. 2011;1:31-45.10.4161/bact.1.1.14942Search in Google Scholar
3 Weinbauer MG and Rassoulzadegan F. Are viruses driving microbial diversification and diversity? Environ Microbiol. 2004;6:1-11.10.1046/j.1462-2920.2003.00539.xSearch in Google Scholar
4 Abedon ST. Phages, ecology, evolution. In: Abedon ST, editor. Bacteriophage ecology: Population growth, evolution, and impact of bacterial viruses. Cambridge University Press; 2008, pp.1-28.10.1017/CBO9780511541483.004Search in Google Scholar
5 Orlova EV. 2012. Bacteriophages and their structural organization. In: Kurtboke I (ed.) Bacteriophages, InTech. Rijeka, Croatia; 2012, pp. 1-30.Search in Google Scholar
6 Hershey A, Chase M. Independent functions of viral protein and nucleic acid in growth of bacteriophage. J Gen Physiol. 1952;36:39-56.10.1007/978-3-662-47150-0_3Search in Google Scholar
7 Clark JR, March JB. Bacteriophages and biotechnology: vaccines, gene therapy and antibacterials. Trends Biotechnol. 2006;24:212-8.10.1016/j.tibtech.2006.03.003Search in Google Scholar
8 McIntyre K, Heap HA, Davey GP, Limsowtin KY. The distribution of lactococcal bacteriophage in the environment of a cheese manufacturing plant. Int Dairy J. 1991;1:183-97.10.1016/0958-6946(91)90010-6Search in Google Scholar
9 Quiberoni A, Tremblay D, Ackermann HW, Moineau S, Reinheimer JA. Diversity of Streptococcus thermophilus phages in a large-production cheese factory in Argentina. J Dairy Sci. 2006;89:3791-9.10.3168/jds.S0022-0302(06)72420-1Search in Google Scholar
10 Gebreselassie HK. Diversity of bacterial strains and bacteriophages in Norwegian cheese production. MSc Thesis. Norwegian University of Life Sciences. 2014.Search in Google Scholar
11 Kleppen HP, Nes IF, Holo H. Characterization of a Leuconostoc bacteriophage infecting flavor producers of cheese starter cultures. Appl Environ Microbiol. 2012;78:6769-72.10.1128/AEM.00562-12Search in Google Scholar
12 Auad L, de Ruiz Holgado AAP, Forsman P, Alatossava T, Raya RR. Isolation and characterization of a new Lactobacillus delbrueckii ssp. bulgaricus temperate bacteriophage. J Dairy Sci. 1997;80:2706-12.10.3168/jds.S0022-0302(97)76231-3Search in Google Scholar
13 Ma C, Pan N, Chen Z, Liu Z, Gong G, Ma A. Geographical diversity of Streptococcus thermophilus phages in Chinese yoghurt plants. Int. Dairy J. 2014;35:32-7.10.1016/j.idairyj.2013.10.007Search in Google Scholar
14 Ma C, Chen Z, Gong G, Huang L, Li S, Ma A. Starter culture design to overcome phage infection during yogurt fermentation. Food Science and Biotechnology. 2015;24:521-7.10.1007/s10068-015-0068-1Search in Google Scholar
15 Lu Z, Breidt F Jr, Fleming HP, Altermann E, Klaenhammer TR. Isolation and characterization of a Lactobacillus plantarum bacteriophage, ΦJL-1, from a cucumber fermentation. International Journal of Food Microbiology. 2003;84:225-35.10.1016/S0168-1605(03)00111-9Search in Google Scholar
16 Lu Z, Perez-Díaz IM, Hayes JS and Breidt F. Bacteriophage ecology in a commercial cucumber fermentation. Appl Environ Microbiol. 2012;78:8571-8.10.1128/AEM.01914-12Search in Google Scholar PubMed PubMed Central
17 Yoon SK, Roudolphe BP, Breidt F Jr and Fleming HP. Detection and characterization of a lytic Pediococcus bacteriophage from the fermenting cucumber brine. J Microbiol Biotechnol. 2007;17:262-70.Search in Google Scholar
18 Lu Z, Breidt F, Plengvidhya V, Fleming HP. Bacteriophage ecology in commercial sauerkraut fermentations. Appl Environ Microbiol. 2003;69:3192-202.10.1128/AEM.69.6.3192-3202.2003Search in Google Scholar PubMed PubMed Central
19 Kleppen HP, Holo H, Jeon SR, Nes IF and Yoon SS. Novel Podoviridae family bacteriophage infecting Weissella cibaria isolated from Kimchi. Appl Environ Microbiol. 2012;78:7299-308.10.1128/AEM.00031-12Search in Google Scholar PubMed PubMed Central
20 Yoon SS, Barrangou-Poueys R, Breidt F Jr, Klaenhammer TR and Fleming HP. Isolation and characterization of bacteriophages from fermenting sauerkraut. Appl Environ Microbiol. 2002;68:973-6.10.1128/AEM.68.2.973-976.2002Search in Google Scholar PubMed PubMed Central
21 Trevors KE, Holley RA, Kempton AG. Effect of bacteriophage on the activity of lactic acid starter cultures used in the production of fermented sausage. J Food Sci. 1984;149:650-3.10.1111/j.1365-2621.1984.tb12492.xSearch in Google Scholar
22 Pringsulaka O, Patarasinpaiboon N, Suwannasai N, Atthakor W, Rangsiruji A. Isolation and characterisation of a novel Podoviridae-phage infecting Weissella cibaria N 22 from Nham, a Thai fermented pork sausage. Food Microbiology. 2011;28:518-25.10.1016/j.fm.2010.10.011Search in Google Scholar PubMed
23 Marco MB, Moineau S, Quiberoni A. Bacteriophages and dairy fermentations. Bacteriophage. 2012;2:149-58.10.4161/bact.21868Search in Google Scholar PubMed PubMed Central
24 Garneau JE and Moineau S. Bacteriophages of lactic acid bacteria and their impact on milk fermentations. Microb Cell Fact. 2011;10:S20.10.1186/1475-2859-10-S1-S20Search in Google Scholar PubMed PubMed Central
25 Leverentz B, Conway WS, Alavidze Z, Janisiewicz WJ, Fuchs Y, Camp MJ, Chighladze E, Sulakvelidze A. Examination of bacteriophage as a biocontrol method for Salmonella on fresh-cut fruit: a model study. J Food Prot. 2001;64:1116-21.10.4315/0362-028X-64.8.1116Search in Google Scholar PubMed
26 Bigwood T, Hudson JA, Billington C, Carey-Smith GV, Heinemann JA. Phage inactivation of foodborne pathogens on cooked and raw meat. Food Microbiol. 2008;25:400-6.10.1016/j.fm.2007.11.003Search in Google Scholar PubMed
27 Chukeatirote E. Thua nao: Thai fermented soybean. J Ethn Foods. 2015;2:115-8.10.1016/j.jef.2015.08.004Search in Google Scholar
28 Agarwal DK, Billore SD, Sharma AN, Dupare BU, Srivastava SK. Soybean: Introduction, improvement, and utilization in India—Problems and prospects. Agric Res. 2013;2:293-300.10.1007/s40003-013-0088-0Search in Google Scholar
29 Liener IE. Implications of antinutritional components in soybean foods. J Crit Rev Food Sci Nutr. 1994;34:31-67.10.1080/10408399409527649Search in Google Scholar PubMed
30 Sarkar PK, Nout MJR. Handbook of indigenous foods involving alkaline fermentation. CRC Press. 2014.10.1201/b17195Search in Google Scholar
31 Kiuchi K, Watanabe S. Industrialization of Japanese Natto. In: Steinkraus KH, editor. Industrialization of indigenous fermented foods. 2nd ed. New York, Marcel Dekker; 2004, pp. 193-246.10.1201/9780203022047.ch4Search in Google Scholar
32 Kwak CS, Lee MS, Park SC. Higher antioxidant of chungkookjang, a fermented soybean paste, may be due to increased aglycone and malonylgycoside isoflavone during fermentation. Nutr Res. 2007;27,719-27.Search in Google Scholar
33 Shin H, Bandara N, Shin E, Ryu S, Kim KP. Prevalence of Bacillus cereus bacteriophages in fermented foods and characterization of phage JBP901. Res Microbiol. 2011;162:791-7.10.1016/j.resmic.2011.07.001Search in Google Scholar PubMed
34 Sarkar PK, Tamang JP, Cook PE, Owens JD. Kinema—A traditional soybean fermented food: Proximate composition and microflora. Food Microbiol. 1994;11:47-55.10.1006/fmic.1994.1007Search in Google Scholar
35 Nout MJR, Bakshi D, Sarkar PK. Microbiological safety of kinema, a fermented soya bean food. Food Control. 1998;9:357-62.10.1016/S0956-7135(98)00126-1Search in Google Scholar
36 Leejeerajumnean A. Thua nao: Alkali fermented soybean from Bacillus subtilis Silpakorn Univ Int J. 2003;3:277-92.Search in Google Scholar
37 Chukeatirote E, Chainun C, Siengsubchart A, Moukamnerd C, Chantawannakul P, Lumyong S, Boontim N, Thakang P. Microbiological and biochemical changes in thua nao fermentation. Res J Microbiol. 2006;1:38-44.Search in Google Scholar
38 Chantawannakul P, Oncharoen A, Klanbut K, Chukeatirote E and Lumyong S. Characterisation of proteases of Bacillus subtilis strain 38 isolated from traditionally fermented soybean in Northern Thailand. ScienceAsia. 2002;28:241-5.10.2306/scienceasia1513-1874.2002.28.241Search in Google Scholar
39 Omafuvbe BO, Abiose SH and Shonukan OO. Fermentation of soybean Glycine max for soy-daddawa production by starter cultures of Bacillus Food Microbiol. 2002;19:561-6.10.1006/fmic.2002.0513Search in Google Scholar
40 Twort FW. An investigation on the nature of ultramicroscopic viruses. The Lancet. 1915;186:1241-3.10.1016/S0140-6736(01)20383-3Search in Google Scholar
41 d’Herelle F. Sur un microbe invisible antagoniste des bacilles dysenteriques. CR Acad. Sci. Paris. 1917;165:373-5. (in French)Search in Google Scholar
42 Haq IU, Chaudhry WN, Akhtar MN, Andleeb S, Qadri I. Bacteriophages and their implications on future biotechnology: a review. Virol J. 2012;9:9.10.1186/1743-422X-9-9Search in Google Scholar PubMed PubMed Central
43 Keen EC. A century of phage research: Bacteriophages and the shaping of modern biology. Bioessays 2015;37:6-9.10.1002/bies.201400152Search in Google Scholar PubMed PubMed Central
44 Macdonald A. Staphylococcus aureus in cows’ milk; the results of phage-typing. Mon Bull Minist Health Emerg Public Health Lab Serv. 1946;5:230-3.Search in Google Scholar
45 McClure WB, Miller AM. Identification of identical strains of staphylococci in food poisoning and other infections by bacteriophage typing. Can Med Assoc J. 1946;55:36-9.Search in Google Scholar
46 Fujii H, Oki M, Makihara M, Keshino J, Takeya R. On the formation of mucilage by Bacillus natto Part VII. Isolation and characterization of a bacteriophage active against “Natto”-producing bacteria, J Agric Chem Soc Jpn. 1967;41:39-43. (in Japanese)10.1271/nogeikagaku1924.41.39Search in Google Scholar
47 Yoshimoto A, Hongo M. Bacteriophages of Bacillus natto 1. Some characteristics of phage NP-1. J Fac Agric, Kyushu U. 1970;16:41-58. (in Japanese)10.5109/22801Search in Google Scholar
48 Yoshimoto A, Nomura S, Hongo M. Bacteriophages of Bacillus natto (IV) Natto plant pollution by bacteriophages. J Ferment Technol. 1970;48:660-8. (in Japanese)Search in Google Scholar
49 Fujii H, Shiraishi A, Kaba K, Shibagaki M, Takahashi S, Honda A. Abnormal fermentation in Natto production and Bacillus natto phages. J Ferment Technol. 1975;53:424-8. (in Japanese)Search in Google Scholar
50 Fujii H, Shiraishi A, Kiryu K, Fujimoto Y. Isolation and some characteristics of Bacillus natto phage PM. Bull Fac Home Life Sci, Fukuoka Women’s U. 1983;14:1-5. (in Japanese)Search in Google Scholar
51 Yamamoto T. Report on the phage contamintion to commercial Natto. Iwate-ken Jozo Shokuhin Shikenjyo Hokoku. 1986;20:106-7. (in Japanese)Search in Google Scholar
52 Tsutsumi T, Hirokawa H, Shishido K. A new defective phage containing a randomly selected 8 kilobase-pairs fragment of host chromosomal DNA inducible in a strain of Bacillus natto FEMS Microbiol Lett. 1990;72:41-6.10.1111/j.1574-6968.1990.tb03858.xSearch in Google Scholar
53 Nagai T, Yamazaki F. Bacillus subtilis (natto) bacteriophages isolated in Japan. Food Sci Technol Res. 2009; 15: 293-8.10.3136/fstr.15.293Search in Google Scholar
54 Umene K, Oohasi S, Yamanaka F, Shiraishi A. Molecular characterization of the genome of Bacillus subtilisnatto bacteriophage PM1, a phage associated with disruption of food production. World J Microbiol Biotechnol. 2009; 25: 1877-81.10.1007/s11274-009-0086-3Search in Google Scholar
55 Hara T, Shiraishi A, Fujii H, Ueda, S. Specific host range of Bacillus subtilisnatto phages associated with polyglutamate production. Agric Biol Chem. 1984;48:2373-4.Search in Google Scholar
56 Hongo M, Yoshimoto A. Formation of phage-induced γ-polyglutamic acid depolymerase in lysogenic strain of Bacillus natto Agric Biol Chem. 1968;32:525-7.10.1080/00021369.1968.10859090Search in Google Scholar
57 Hongo M, Yoshimoto A. Bacteriophages of Bacillus natto Part II. Induction of γ-polyglutamic acid depolymerase following phage infection. Agric Biol Chem. 1970;34:1047-54.Search in Google Scholar
58 Hongo M, Yoshimoto A. Bacteriophages of Bacillus natto Part III. Action of phage-induced γ-polyglutamic acid depolymerase on γ-polyglutamic acid and the enzymatic hydrolyzates. Agric Biol Chem. 1970;34:1055-63.10.1271/bbb1961.34.1055Search in Google Scholar
59 Nagai T, Itoh Y. Characterization of a generalized transducing phage of poly-γ-glutamic acid-producing Bacillus subtilis and its application for analysis of Tn917-LTV1 insertional mutants defective in poly-γ-glutamic acid production. Appl Environ Microbiol. 1997;63:4087-9.10.1128/aem.63.10.4087-4089.1997Search in Google Scholar PubMed PubMed Central
60 Kimura K, Itoh Y. Characterization of poly-γ-glutamate hydrolase encoded by a bacteriophage genome: Possible role in phage infection of Bacillus subtilis encapsulated with poly-γ-glutamate. Appl Environ Microbiol. 2003;69:2491-7.10.1128/AEM.69.5.2491-2497.2003Search in Google Scholar PubMed PubMed Central
61 Kimura K. γ-PGA hydrolase of phage. In: K Kiuchi, T Nagai, K Kimura, editors. Advanced science on Natto: Japanese soybean fermented foods. Tokyo, Kenpakusha; 2008, pp. 268-70. (in Japanese)Search in Google Scholar
62 Fujimoto Z, Shiga I, Itoh Y, Kimura K. Crystallization and preliminary crystallographic analysis of poly‐γ‐glutamate hydrolase from bacteriophage ΦNIT1. Acta Cryst. 2009;65:913-6.10.1107/S1744309109029881Search in Google Scholar PubMed PubMed Central
63 Ozaki T, Abe N, Kimura K, Suzuki A, Kaneko J. Genomic analysis of Bacillus subtilis lytic bacteriophage ϕNIT1 capable of obstructing natto fermentation carrying genes for the capsule-lytic soluble enzymes poly-γ-glutamate hydrolase and levanase. Biosci Biotechnol Biochem. 2017;81:135-46.10.1080/09168451.2016.1232153Search in Google Scholar PubMed
64 Nagai T. Bacteriophages of Bacillus subtilisnatto and their contamination in Natto factories. In: I Kurtboke, editor. Bacteriophages. Shanghai, InTech; 2012a, pp. 95-110.10.5772/33555Search in Google Scholar
65 Nagai T. Overview of studies on Bacillus subtilisnatto bacteriophages and the prospects. Jpn Agric Res Q. 2012b; 46: 305-10.10.6090/jarq.46.305Search in Google Scholar
66 Nakajima J. Inprovement of quality of Natto: Research on contamination of Bsubtilisnatto phages in Natto factories. Reports of the Ibaraki Prefectural Industrial Technology Center. 1995. (in Japanese)Search in Google Scholar
67 Lee ZS. Studies on the isolation and characterization of bacteriophage of Bacillus subtilis var. 816. Kor J Microbiol. 1978;16:71-8. (in Korean)Search in Google Scholar
68 Bandara N, Jo J, Ryu S, Kim KP. Bacteriophages BCP1-1 and BCP8-2 require divalent cations for efficient control of Bacillus cereus in fermented foods. Food Microbiol. 2012;31:9-16.10.1016/j.fm.2012.02.003Search in Google Scholar PubMed
69 Asare PT, Bandara N, Jeong TY, Ryu S, Klumpp J, Kim KP. Complete genome sequence analysis and identification of putative metallo-beta-lactamase and SpoIIIE homologs in Bacillus cereus group phage BCP8-2, a new member of the proposed Bastille-like group. Arch Virol. 2015;160:2647-50.10.1007/s00705-015-2548-2Search in Google Scholar PubMed
70 Asare PT, Ryu S, Kim KP. Complete genome sequence and phylogenetic position of the Bacillus cereus group phage JBP901. Arch Virol. 2015;160:2381-4.10.1007/s00705-015-2485-0Search in Google Scholar PubMed
71 Oh H, Seo DJ, Jeon SB, Park H, Jeong S, Chun HS, Oh M, Choi C. Isolation and characterization of Bacillus cereus bacteriophages from foods and soil. Food Env Virol. 2017;9:260-9.10.1007/s12560-017-9284-6Search in Google Scholar PubMed
72 Kim EJ, Hong JW, Yun NR, Lee YN. Characterization of Bacillus phage-K2 isolated from chungkookjang, a fermented soybean foodstuff. J Ind Microbiol Biotechnol. 2011;38:39-42.10.1007/s10295-010-0825-3Search in Google Scholar PubMed
73 Ghosh K, Kang HS, Hyun WB, Kim KP. High prevalence of Bacillus subtilis-infecting bacteriophages in soybean-based fermented foods and its detrimental effects on the process and quality of Cheonggukjang. Food Microbiol. 2018;76:196-203.10.1016/j.fm.2018.05.007Search in Google Scholar PubMed
74 Ogawa Y, Yamaguchi F, Yuasa K, Tahara Y. Efficient production of γ-polyglutamic acid by Bacillus subtilisnatto in jar fermenters. Biosci Biotech Biochem. 1997;61:1684-7.10.1271/bbb.61.1684Search in Google Scholar PubMed
75 Ackermann HW. Bacteriophage classification. In: Kutter E, Sulakvelidze A, editors. Bacteriophages: Biology and applications. Boca Raton, CRC Press; 2005, pp. 67-89.Search in Google Scholar
76 Higuchi T, Uchida K, Abe K. Preparation of phage-insensitive strains of Tetragenococcus halophile and its application for soy sauce fermentation. Biosci Biotech Biochem. 1999;63:415-7.10.1271/bbb.63.415Search in Google Scholar PubMed
77 Ogunleye A, Bhat A, Irorere VU, Hill D, Williams C, Radecka I. Poly-γ-glutamic acid: production, properties and applications. Microbiology 2015;161:1-17.10.1099/mic.0.081448-0Search in Google Scholar PubMed
78 Candela T, Fouet A. Poly-gamma-glutamate in bacteria. Mol Microbiol. 2006;60:1091-8.10.1111/j.1365-2958.2006.05179.xSearch in Google Scholar PubMed
79 Zhao XQ, Park KH, Jin YY, Lee IH, Yang YY, Suh JW. Isolation and characterization of a new γ-polyglutamic acid producer, Bacillus mesentericus MJM1, from Korean domestic Chungkukjang bean paste. J Microbiol Biotechnol. 2005;15:59-65.Search in Google Scholar
80 Lee NR, Lee SM, Cho KS, Jeong SY, Hwang DY, Kim DS, Hong CO, Son HJ. Improved production of poly-γ-glutamic acid by Bacillus subtilis D7 isolated from Doenjang, a Korean traditional fermented food, and its antioxidant activity. Appl Biochem Biotechnol. 2014;173:918-32.10.1007/s12010-014-0908-0Search in Google Scholar PubMed
81 Chettri R, Bhutia MO, Tamang JP. Poly-γ-glutamic acid (PGA)-producing Bacillus species isolated from Kinema, Indian fermented soybean food. Front Microbiol. 2016;7:971.10.3389/fmicb.2016.00971Search in Google Scholar PubMed PubMed Central
82 Choi SH, Sung C, Choi WY. Levan-producing Bacillus subtilis BS 62 and its phylogeny based on its 16S rDNA sequence. J Microbiol Biotechnol. 2001;11:428-34.Search in Google Scholar
83 Shih IL, Yu YT, Shieh CJ, Hsieh CY. Selective production and characterization of levan by Bacillus subtilis (Natto) Takahashi. J Agric Food Chem. 2005;53:8211-5.10.1021/jf058084oSearch in Google Scholar PubMed
84 Xu Q, Yajima T, Li W, Saito K, Ohshima Y, Yoshikai Y. Levan (β‐2, 6‐fructan), a major fraction of fermented soybean mucilage, displays immunostimulating properties via Toll‐like receptor 4 signalling: induction of interleukin‐12 production and suppression of T‐helper type 2 response and immunoglobulin E production. Clin Exp Allergy 2006;36:94-101.10.1111/j.1365-2222.2006.02401.xSearch in Google Scholar PubMed
85 Thai Industrial Standards Institute (TISI). Community Product Standards: Thua Nao powder and Thua Nao disc. Bangkok: TISI; 2005. Available from: http://tcps.tisi.go.th/pub/tcps1057_48.pdf, and http://tcps.tisi.go.th/pub/tcps509_47.pdf.Search in Google Scholar
86 Food and Drug Administration (FDA), Thailand. Microbiological Food Safety. Bangkok: Thai FDA; 2013. Available from: .Search in Google Scholar
87 Food and Agriculture Organization of the United Nations (FAO). Codex. Rome: FAO; 2004. Available from: www.fao.org/faowho-codexalimentarius/en.Search in Google Scholar
88 ICMSF (International Commission on Microbiological Specifications for Foods). Microorganisms in Foods 6, Microbial Ecology of Food Commodities (2nd ed.). London: Kluwer Academic/Plenum Publishers; 2005, pp. 360–72.Search in Google Scholar
89 Takiguchi T, Yoshino I, Yuasa H, Kawano I, Aoki Y. Pollution of fermentation process of Natto by some kinds of bacteriophage. Reports of Gunma Prefectural Industrial Technology Research Laboratory; 1999, pp. 35-39. (in Japanese)Search in Google Scholar
90 Suriyanoi K, Chumpurat P, Beamon RA, Suvegoon W, Suksai U, Chukeatirote E. Microbiological quality of commercial Thua Nao, a Thai fermented soybean product. Pharm Sci Asia. 2018;45:231-5.10.29090/psa.2018.04.017.0061Search in Google Scholar
© 2018 Ekachai Chukeatirote et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 License.
Articles in the same Issue
- Review
- Secretion of full-length Tau or Tau fragments in cell culture models. Propagation of Tau in vivo and in vitro
- Research Article
- DNA testing of edible crabs from seafood shops on the Odisha coast, India
- Communication
- Overexpression and pre-treatment of recombinant human Secretory Leukocyte Protease Inhibitor (rhSLPI) reduces an in vitro ischemia/reperfusion injury in rat cardiac myoblast (H9c2) cell
- Review
- Global Perspective of Novel Therapeutic Strategies for the Management of NeuroAIDS
- Research Article
- Current insights on use of growth factors as therapy for Intervertebral Disc Degeneration
- Research Article
- Treatment of SEC62 over-expressing tumors by Thapsigargin and Trifluoperazine
- Review
- Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases
- Research Article
- Polymorphisms in DNA repair genes increase the risk for type 2 diabetes mellitus and hypertension
- Review
- Small heat shock proteins and neurodegeneration: recent developments
- Review Article
- The rise of genetically engineered mouse models of pancreatitis: A review of literature
- Research Article
- Early Life Trauma Predicts Affective Phenomenology and the Effects are Partly Mediated by Staging Coupled with Lowered Lipid-Associated Antioxidant Defences
- Review Article
- Significance of bacteriophages in fermented soybeans: A review
- Review
- Biased receptor functionality versus biased agonism in G-protein-coupled receptors
- Research Article
- Neural like cells and acetyl-salicylic acid alter rat brain structure and function following transient middle cerebral artery occlusion
- Research Article
- Enhancing the anticoagulant profile of meizothrombin
- Mini review
- Can uterine secretion of modified histones alter blastocyst implantation, embryo nutrition, and transgenerational phenotype?
- Research Article
- In vitro and in vivo evaluation of colon cancer targeted epichlorohydrin crosslinked Portulaca-alginate beads
- Mini review
- Prospects of Pharmacological Interventions to Organismal Aging
- When Humans Met Superbugs: Strategies to Tackle Bacterial Resistances to Antibiotics
- Special Issue: Recent Advances in Basic and Clinical Medicine
- Beneficial effects of Spirogyra Neglecta Extract on antioxidant and anti-inflammatory factors in streptozotocin-induced diabetic rats
Articles in the same Issue
- Review
- Secretion of full-length Tau or Tau fragments in cell culture models. Propagation of Tau in vivo and in vitro
- Research Article
- DNA testing of edible crabs from seafood shops on the Odisha coast, India
- Communication
- Overexpression and pre-treatment of recombinant human Secretory Leukocyte Protease Inhibitor (rhSLPI) reduces an in vitro ischemia/reperfusion injury in rat cardiac myoblast (H9c2) cell
- Review
- Global Perspective of Novel Therapeutic Strategies for the Management of NeuroAIDS
- Research Article
- Current insights on use of growth factors as therapy for Intervertebral Disc Degeneration
- Research Article
- Treatment of SEC62 over-expressing tumors by Thapsigargin and Trifluoperazine
- Review
- Interferon-gamma (IFN-γ): Exploring its implications in infectious diseases
- Research Article
- Polymorphisms in DNA repair genes increase the risk for type 2 diabetes mellitus and hypertension
- Review
- Small heat shock proteins and neurodegeneration: recent developments
- Review Article
- The rise of genetically engineered mouse models of pancreatitis: A review of literature
- Research Article
- Early Life Trauma Predicts Affective Phenomenology and the Effects are Partly Mediated by Staging Coupled with Lowered Lipid-Associated Antioxidant Defences
- Review Article
- Significance of bacteriophages in fermented soybeans: A review
- Review
- Biased receptor functionality versus biased agonism in G-protein-coupled receptors
- Research Article
- Neural like cells and acetyl-salicylic acid alter rat brain structure and function following transient middle cerebral artery occlusion
- Research Article
- Enhancing the anticoagulant profile of meizothrombin
- Mini review
- Can uterine secretion of modified histones alter blastocyst implantation, embryo nutrition, and transgenerational phenotype?
- Research Article
- In vitro and in vivo evaluation of colon cancer targeted epichlorohydrin crosslinked Portulaca-alginate beads
- Mini review
- Prospects of Pharmacological Interventions to Organismal Aging
- When Humans Met Superbugs: Strategies to Tackle Bacterial Resistances to Antibiotics
- Special Issue: Recent Advances in Basic and Clinical Medicine
- Beneficial effects of Spirogyra Neglecta Extract on antioxidant and anti-inflammatory factors in streptozotocin-induced diabetic rats