Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma
-
Tianhao Sun
 und
Zhi-Li Liu
und
Zhi-Li Liu

Abstract
Osteosarcoma (OS) is the most common type of primary malignant bone tumor. The early lung metastasis of osteosarcoma is one of the main factors of poor prognosis. Therefore, searching for new targets and new mechanisms of osteosarcoma metastasis is essential for the prevention and treatment of osteosarcoma. Our previous studies suggested that fatty acid synthase (FASN) was an oncogene and promoted osteosarcoma. In addition, it is reported that the expression of miR-195 was negatively correlated with osteosarcoma. Aberrant DNA methylation can reversely regulate the expression of miRNAs. However, whether miR-195 could target FASN in osteosarcoma and whether ectopic DNA methylation is the upstream regulatory mechanism of miR-195 in metastasis of osteosarcoma are not fully studied. The expressions were detected by qPCR and western blot, and methylation level was determined by methylation-specific PCR. Luciferase reporter assay, MTT, wound healing, and Transwell assay were used. We found that the expression of miR-195 was low in osteosarcoma. The methylation of miR-195 was high. miR-195 targeted and decreased the expression of FASN. In osteosarcoma, miR-195 inhibited cell proliferation, cell migration, and invasion. The methylation of miR-195 was related to decreased miR-195, it might promote osteosarcoma.
1 Introduction
Osteosarcoma (OS) is the most common bone tumor and the eighth form of cancer in children [1]. It is classified as osteoblastic, chondroblastic, or fibroblastic osteosarcoma [2]. The morbidity of osteosarcoma accounts for 35% of all the primary bone cancers. Recurrence and lung metastasis are the leading cause of death in patients [3]. Migration is the primary step in the process of metastasis, with the main event involving epithelial–mesenchymal transition that allows the cells to become more motile [4]. Metastasis remains the most vital complication of osteosarcoma. The survival rate of patients with metastasis is low [5]. Osteosarcoma usually exhibits locally invasive growth. Pulmonary metastasis is frequently found in patients [6]. While only 20% of patients with metastasis of osteosarcoma are detectable clinically, the majority of the remaining 80% are presumed to have undetectable micro-metastases of lungs at diagnosis [7]. Precision medicine suggests one to find molecules such as proteins or miRNAs as the therapy targets. Thus, searching for effective and sensitive molecular biomarkers for osteosarcoma is urgently needed.
Fatty acids include substantial functions in cells from membrane structure and energy storage to signaling transduction and protein acylation [8]. Fatty acids are the main ingredient of phospholipids and glycolipids, which are also the main ingredient of cell membranes. Glycolipid plays important role in cell growth, cell differentiation, and signal transduction [9]. In human beings, the most common type of fatty acid is palmitic acid. The fatty acid is oxygenated and degraded into acetyl-coenzyme in the mitochondria, which provide energy. A fatty acid is also responsible for the formation of certain second messengers involved in the response to extracellular signals [9].
The fatty acid synthesis pathway has two main functions: one is the storage of excess energy intake such as fat, and the other is the synthesis of fat from carbohydrates or proteins if the diet is low in fat [8]. There are correlations between abnormal fatty acid metabolism and tumor such as unlimited cell proliferation and invasiveness, consistent with the hypothesis that fatty acid metabolism disorder might be one of the factors inducing carcinogenesis [10]. Fatty acid synthase (FASN) is the main synthetic enzyme that produces the free fatty acid palmitate. Because humans consuming a proper diet undergo little endogenous fatty acid synthesis, dietary lipids would provide the necessary fatty acid for survival, growth, and development.
microRNAs (miRNAs) are small, single-stranded RNA regulating gene expression [11]. They regulate substantial physiological and pathological processes including cell proliferation, cell differentiation, and cell death. One miRNA can bind to numerous mRNAs, resulting in the regulation of genes [12]. Thus, aberrant miRNA expression is associated with diseases such as osteosarcoma [13], osteoporosis [14], and osteoarthritis [15]. One of the facts of miRNAs is that they can bind to their target mRNAs and then degrade targets. During the process of degradation, argonaute family proteins play vital roles. Translational repression and 5′-to-3′ mRNA degradation lead to the repression of the target gene expression. Overexpressed miRNAs in malignant cells are related to carcinogenesis, and target and decrease the expression levels of tumor suppressors. In contrast, miRNAs lost by tumors lead to the expression of the oncogene. The next-generation sequencing technologies have made an investigation of miRNA expression profiling easy.
The CpG sites are regions of DNA where a cytosine is followed by a guanine. Cytosine in CpG can be methylated, which can change gene expression. CpG islands also occur frequently in promoters for miRNAs [16,17,18], which inhibit mRNA transcription or induce mRNA degradation and shed new light on cancer research [19,20,21,22,23,24,25]. In cancer, the CpG islands are commonly methylated and the related genes or miRNAs are downregulated.
Reports found that the expression level of miR-195 was negatively correlated with osteosarcoma metastasis, and upregulating miR-195 could inhibit the apoptosis of osteosarcoma cells. Our previous study showed silencing FASN could inhibit cell proliferation and migration of osteosarcoma [13]. However, whether miR-195 could target FASN in osteosarcoma was not fully studied. In this study, we will investigate the relationships between miR-195 and FASN, and the methylation of the promotor region of miR-195.
2 Materials and methods
2.1 Clinical tissues and cell culture
We selected six fresh osteosarcoma tissues and six normal tissues from clinical tissue biopsy in The First Affiliated Hospital of Nanchang University from September 2020 to July 2021. All patients were selected based on X-ray, CT, bone scan of the body, and pathology methods. Before the collection of samples, the patients were not treated by drugs or radiotherapy. All of the studies were approved by the Ethics Committee of the above hospital (2020) (12–103), and all patients signed the Informed Consent Form. The study methodologies conformed to the standards set by the Declaration of Helsinki. One of our authors had access to information that could identify individual participants during or after data collection. Osteosarcoma cell lines 143B and U2-OS were cultured in DMEM and DMEM/F12, respectively, with 10% FBS and penicillin/streptomycin.
2.2 Quantitative real-time PCR (qPCR)
All 100 mg tissues were mixed with 1 mL of TRIzol. The same amount of RNA was used for all the samples. Each cDNA sample was triplicated in 96-well plates. Data were analyzed using the 2−ΔΔCT relative quantification method. U6 was the internal control. The sequences of primers were as follows: miR-195: 5′-UAGCAGCACAGAA AUAUUGGC-3′; U6 forward: 5′-ATGACACGCAAATTCGTGAAGC-3′; miRNAs reverse: 5′-GCGAGCACAGAATTAATACGAC-3′.
2.3 Western blot
The lysis buffer was a RIPA buffer with a protease inhibitor cocktail. The cells were added to lysis buffer and subjected to alternating vortexing and then stored at 4°C for half an hour. Then, the cell lysis was performed by centrifugation at 16,000g for 20 min at 4°C. Laemmli buffer was used to denature the samples. The PVDF membrane was blocked by 5% BSA solution for 1 h. Primary (first) antibodies included anti-FASN antibody (sc-55580, mouse anti-human, 1:100) and GAPDH antibody (Abcam, ab9485, 1:2,500). The membrane was incubated with the primary antibody overnight at 4°C, rinsed with TBST, and then incubated with the secondary antibody for 1 h at room temperature. ECL Prime Western Blotting Reagents (GE Healthcare) were used to develop.
2.4 DNA extraction and methylation-specific PCR (MSP)
A DNA extraction kit (Biomed, Beijing, China) was used to extract DNA from osteosarcoma cells, and the EZ Methylation kit (Zymo Research, Orange, CA) was used for bisulfite modification of genomic DNA (500 ng). The DNA methylation status was analyzed by MSP using primers specific for either the methylated or bisulfate modified unmethylated DNA. Ten microliters of PCR products was loaded onto 6% polyacrylamide gels, electrophoresed, stained with ethidium bromide, and visualized under ultraviolet light. The MSP primers for miR-195 were as follows: left M primer: AGTGTTTTTTGTTTTTAGGAGAGAC, right M primer: TCTAATCTCAACCTTTAATTTCCGT; left U primer: GTTTTTTGTTTTTAGGAGAGATGT; right U primer: TCTCTAATCTCAACCTTTAATTTCCAT.
2.5 Luciferase reporter assays
First, we designed sequences including the two sequences of FASN as follows: UTR primers (5′-3′) U: CCCCTCGAGCCTGCCACCGGAGGTCACT, L: CGGGCGGoCGCGTGGGAGGCTGAGAGCAGCA; UTR mutant primers (5′-3′): U: TTATACAAAACAAAAGCGATAA, L: AAGCAACAGAAACCCCCTGT. We inserted the wild type and mutant fragments of the 3′-UTR of FASN containing the binding sites of miR-195 into the luciferase vector. We co-transfected the cells with the above plasmids and miR-195. The transfection method for luciferase assays is according to the instructions of LipofectamineTM 2000 (Lipo2000). One day before transfection, the OS cells in the serum-free medium were seeded in the 48-well plate. For each transfection sample, the following amounts of reagents were used. A total of 100 ng of plasmids psi-FASN/psi-FASNm (V1 = 0.25 µL) was diluted in 25 µL of serum-free medium. At the same time, miR-195 or negative controls (V2 = 6 µL) was diluted in 25 µL of serum-free medium. Lipo2000 (V3 = 1 µL) was diluted in 25 µL of serum-free medium. The above V1, V2, and V3 were mixed and incubated for 20 min, and then added into the cell medium. After 5 h, the above medium was changed to the culture medium. The cells were cultured for 48 h.
2.6 Lentiviruses silencing or overexpressing miR-195
For lentiviruses overexpressing miR-195, the vector was pSico-GFP. We first designed sequences xhoI-U 5′-3′: AATCTCGAGATGGAGEAGTATACAGCAAAC, bglII-L 5′-3′: AATAGATCTAAGGACACTCGGATGATCTGTG. The PCR fragments of miR-195 were digested with xhoI and bglII. The inserts were ligated into the vector. Plasmid construction, molecular cloning, and packaging of lentiviruses were used. For lentiviruses silencing miR-195, the vector was pSico-RNAi-GFP. The sequences of the primers used for PCR were as follows: (5′-3′) shU: ACTGACTGACCAAACAGCAAAGTTCGACAGAGCACAGGACACAAGGCCTGTTAC, shL: GGCCAAAACCAAACAGCAATAGTTCGACAGAGCACAGCATACAGCCTCTAGCAA. The packaging procedure of lentivirus contains the following steps. 24 h before transfection, the 293T cells were seeded at a concentration of 1.5 × 106 cells/mL in a 15 cm cell dish. Then, 2 h before transfection, the cell culture medium was changed to a serum-free medium. The DNA (pGC-LV vector 10 µg, pHelper 1.0 vector 10 µg, pHelper 2.0 vector 12 µg) was mixed with Opti-MEM and incubated for 5 min at room temperature. The Opti-MEM (2.4 mL) was then mixed with 100 µL of Lipofectamine 2000 and incubated for 3–5 min at room temperature. The diluted DNA and Lipofectamine 2000 were mixed and incubated and then added into the 293T cell medium. The transfection procedure of lentivirus contains the following steps: 18–24 h before transfection, the attached osteosarcoma cells were seeded at a concentration of 1 × 105 cells/well in 24 well plates. The cell medium was changed into a new medium (2 mL) containing 6 mg/mL polybrene and then lentiviruses were added. After 4 h, 2 mL of medium was added to dilute the concentration of polybrene; 24 h later, the medium containing lentiviruses was changed.
2.7 MTT cell proliferation assay
We seeded cells at a concentration of 1 × 104 cells/well with a total of 150 µL of the culture medium with FBS and antibiotics into 96-well microplates. Cell viability was checked after 1, 2, 3, 4, and 5 days. We conducted this assay by using the Cell Growth Determination Kit based on MTT (Sigma-Aldrich). In brief, we added 5 mg/mL MTT without phenol red in an amount equal to 10% of the culture volume (15 µL), incubated for 3 h, removed the culture fluid, dissolved the resulting MTT formazan crystals by adding 0.1 N HCl in isopropanol, mixed and spectrophotometrically measured the absorbance at a wavelength of 490 nm, and subtracted the background absorbance measured at 690 nm. All the wells were quadruple repetitions. Every four wells in 96-well microplates were manipulated in the same way. For each independent experiment, three 96-well microplates were used for detection on Day 1, Day 2, Day 3, Day 4, and Day 5 respectively.
2.8 Wound healing migration assay
Cells were cultured in 6-well tissue culture plates. A micropipette tip was used to scratch through the cell monolayer at the center of the plate. After two washes with phosphate-buffered saline, the medium was replaced with a medium containing 10% FBS. The cells were incubated for 24 h at 37°C and 5% CO2 and were imaged at 0 and 24 h.
2.9 Transwell invasion assay
Transwell invasion chambers (Corning Inc., Corning, NY) were coated with Matrigel (50 µL per filter) (BD Biosciences, Franklin Lakes, NJ). Cells (8 × 104/200 µL/chamber) resuspended in a medium containing 10 g/L bovine serum albumin were added to the upper chamber, and 500 µL of the medium supplemented with 10% FBS was added to the lower chamber as a chemoattractant. After 24 h, the cells in the upper chamber were removed with a cotton swab, and those that had passed through the Matrigel-coated membrane were stained with 1% crystal violet (Solarbio, Beijing, China), photographed (200× magnification), and counted.
2.10 Statistical analysis
Statistical analysis was performed by an independent sample t-test, and p < 0.05 was considered statistically significant.
3 Results
3.1 Expression of miR-195 and FASN and the methylation level of miR-195 in the osteosarcoma tissues
The expression levels of miR-195 were significantly lower in the tissues of osteosarcoma than that in nontumors (Figure 1a). The protein expression of FASN of fresh osteosarcoma tissues was significantly higher than that of fresh normal bone tissue (Figure 1b). The methylation levels in the promoter region of miR-195 were significantly higher in the tissues of osteosarcoma than that in nontumors (Figure 1c).
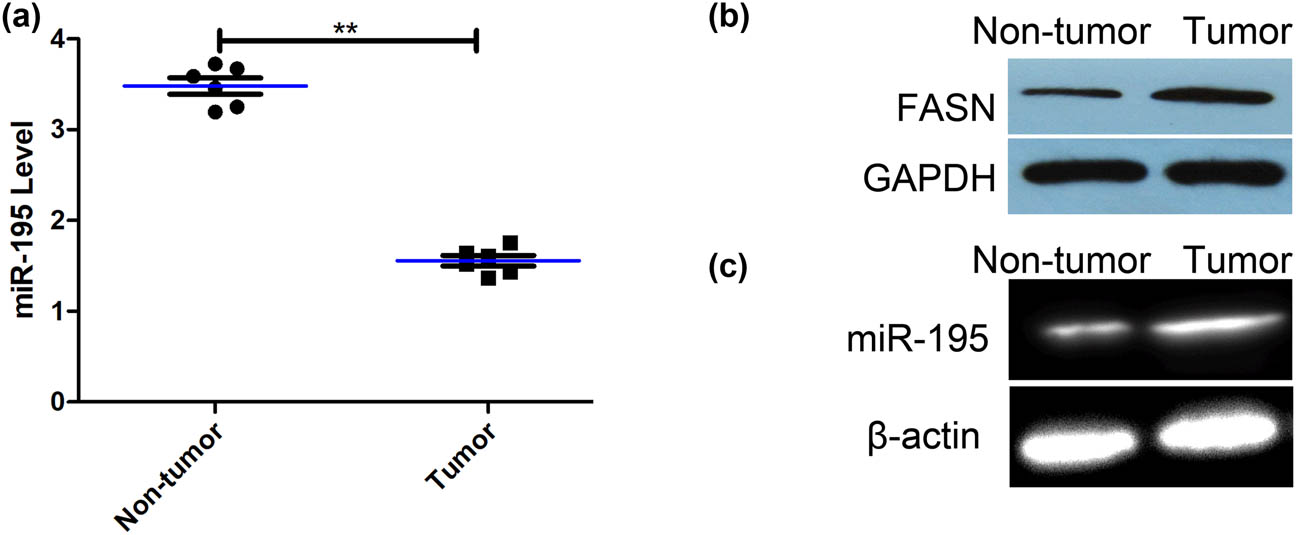
The expression of miR-195 and its methylation level in the osteosarcoma tissues. (a) The expression of miR-195 of fresh osteosarcoma tissues was significantly lower than that of fresh normal bone tissue (n = 6; p < 0.001). The error bar represents the standard variation of the miR-195 levels in patients with or without osteosarcoma. (b) The western blot assay showed that the expression of FASN of fresh osteosarcoma tissues was significantly higher than that of fresh normal bone tissues (n = 6). (c) MSP assay showed that the methylation levels of promoter regions of miR-195 in osteosarcoma tissues were significantly higher than that in fresh normal bone tissues (n = 6). *p < 0.05, **p < 0.01, n = 6/group.
3.2 miR-195 targeted and decreased the expression of FASN
The bioinformatics prediction tool Target Scan Human 6.0 indicated that there was a binding site between the miR-195 and 3′-UTR of FASN (Figure 2a). Luciferase assay verified that in the cells transfected with FASN 3′-UTR plasmids and miR-195, the luciferase activity decreased (Figure 2b), while the mutant plasmids had no effects, suggesting that miR-195 specifically targeted FASN. Next, we constructed lentiviruses silencing or overexpressing miR-195, and our transfection was successful (Figure 2c). miR-195 overexpression suppressed expression of FASN while miR-195 inhibition upregulated FASN at both mRNA and protein levels in both U2-OS and 143B cells (Figure 2d and e).
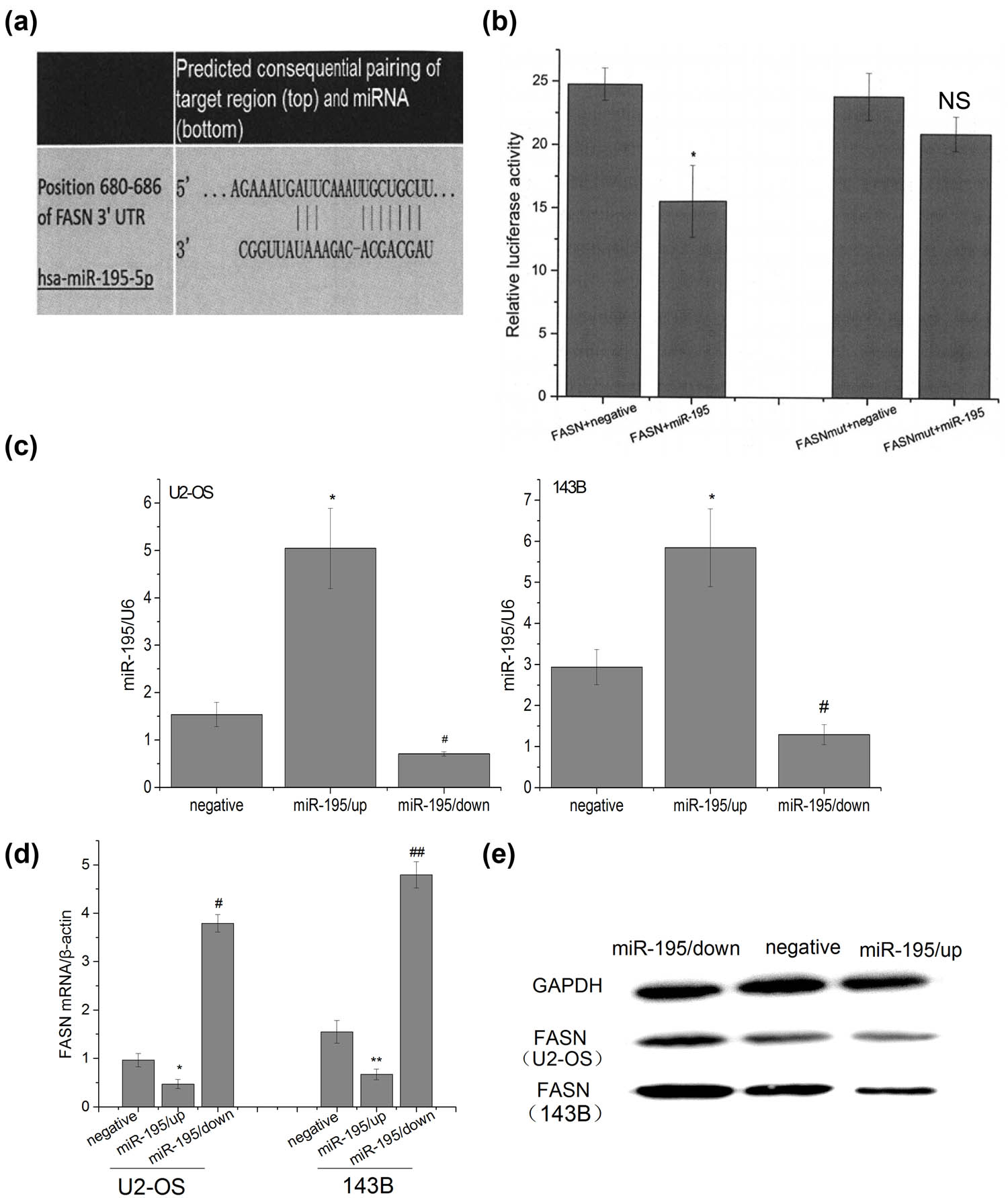
miR-195 targeted and decreased the expression of FASN. (a) Target Scan Human 6.0 software predicted that there was a target site of miR-195 on 680–686 bp of 3′-UTR of FASN. (b) Luciferase activity assay showed the following: the luciferase activity in the group psi-FASN Vector + miR-195 was significantly lower than that in the group FASN + negative controls (n = 6; p < 0.001), but there were no significant differences between the group psi-FASN mutVector + miR-195 and the group FASN mut + negative (n = 6: #p = 0.083), and the luciferase activity in the group psi-FASNmut Vector + miR-195 was significantly higher than that in the group psi-FASN Vector + miR-195 (n = 6; **p < 0.001). The error bar represents the standard variation of the luciferase activity in the cells transfected with FASN (or mutant FASN) plasmids and treated with or without miR-195. (c) The expression level of miR-195 was significantly up/downregulated in U2-OS and 143B cells transfected by miR-195/up or miR-195/down lentiviral vector (n = 6). The error bar represents the standard variation of the miR-195 levels in the U2-OS or 143B cells transfected with lentiviruses overexpressing or inhibiting miR-195. (d) The FASN mRNA expression of osteosarcoma cells decreased significantly in the group miR-195/up and significantly upregulated in the group miR-195/down in vitro by RT-PCR analysis (n = 6). The error bar represents the standard variation of the FASN levels in the U2-OS or 143B cells transfected with lentiviruses overexpressing or inhibiting miR-195. (e) The FASN protein expression of osteosarcoma cells decreased significantly in the group miR-195/up and significantly upregulated in the group miR-195/down in vitro by western blot analysis (n = 6).
3.3 miR-195 inhibited cell proliferation, cell migration, and invasion in osteosarcoma
We then studied the functions of miR-195 in osteosarcoma cells. Silence of miR-195 promoted cell proliferation while overexpression of miR-195 inhibited cell proliferation in both U2-OS and 143B cells (Figure 3a). Overexpression of miR-195 also leads to reduced cell migration in U2-OS and 143B (Figure 3b). The reduced function of miR-195 increased cell invasion while the gain of function of miR-195 inhibited cell invasion (Figure 3c).
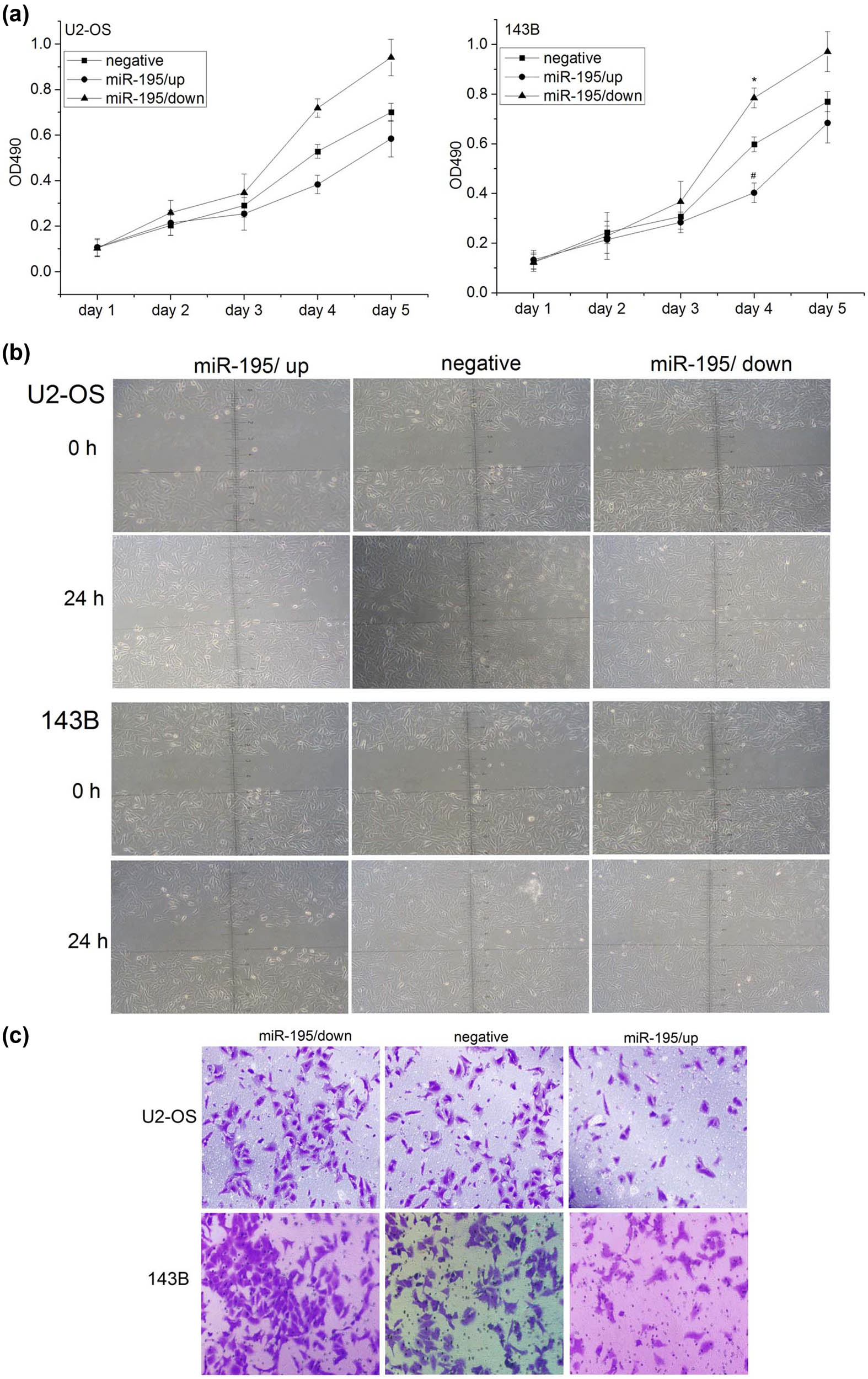
miR-195 inhibited cell proliferation, cell migration, and invasion in osteosarcoma. (a) The proliferation activity of osteosarcoma cells was detected by MTT assay. Left: the proliferation ability of U2-OS cells was significantly decreased in the group miR-195/up (n = 6; day 4: *p < 0.001) and significantly increased in the group miR-195/down in vitro (n = 6; day 4: #p < 0.001). Right: the proliferation ability of 143B cell was significantly decreased in the group miR-195/up (n = 6; day 4: *p < 0.001) and significantly increased in the group miR-195/down in vitro (n = 6; day 4: #p < 0.001). The error bar represents the standard variation of the OD490 in the U2-OS or 143B cells transfected with lentiviruses overexpressing or inhibiting miR-195. (b) The migration ability of osteosarcoma cells at 24 h using wound healing assay was significantly decreased in the group miR-195/up (n = 6, ×200). (c) The invasion ability of osteosarcoma cells at 24 h using Transwell invasion assay was significantly decreased in the miR-195/up group and significantly increased in the miR-195/down group (n = 6, ×200).
3.4 Methylation of miR-195 was one of the reasons why miR-195 decreased
We used the CpG island online prediction tool (http://cpgislands.usc.edu) to explore the possibility of methylation of the promoter region of miR-195 and found three CpG islands in the promoter of miR-195 (Figure 4a). We used methylation inhibition reagent 5-aza-2′-dc to treat U2-OS and 143B cells and detected the methylation level. The results showed that the methylation level significantly decreased after the drug was used (Figure 4b). The expression levels of miR-195 increased significantly after the cells were treated with 5-aza-2′-dc (Figure 4c), indicating that the methylation of the promoter region of miR-195 was one of the reasons why miR-195 decreased in osteosarcoma.
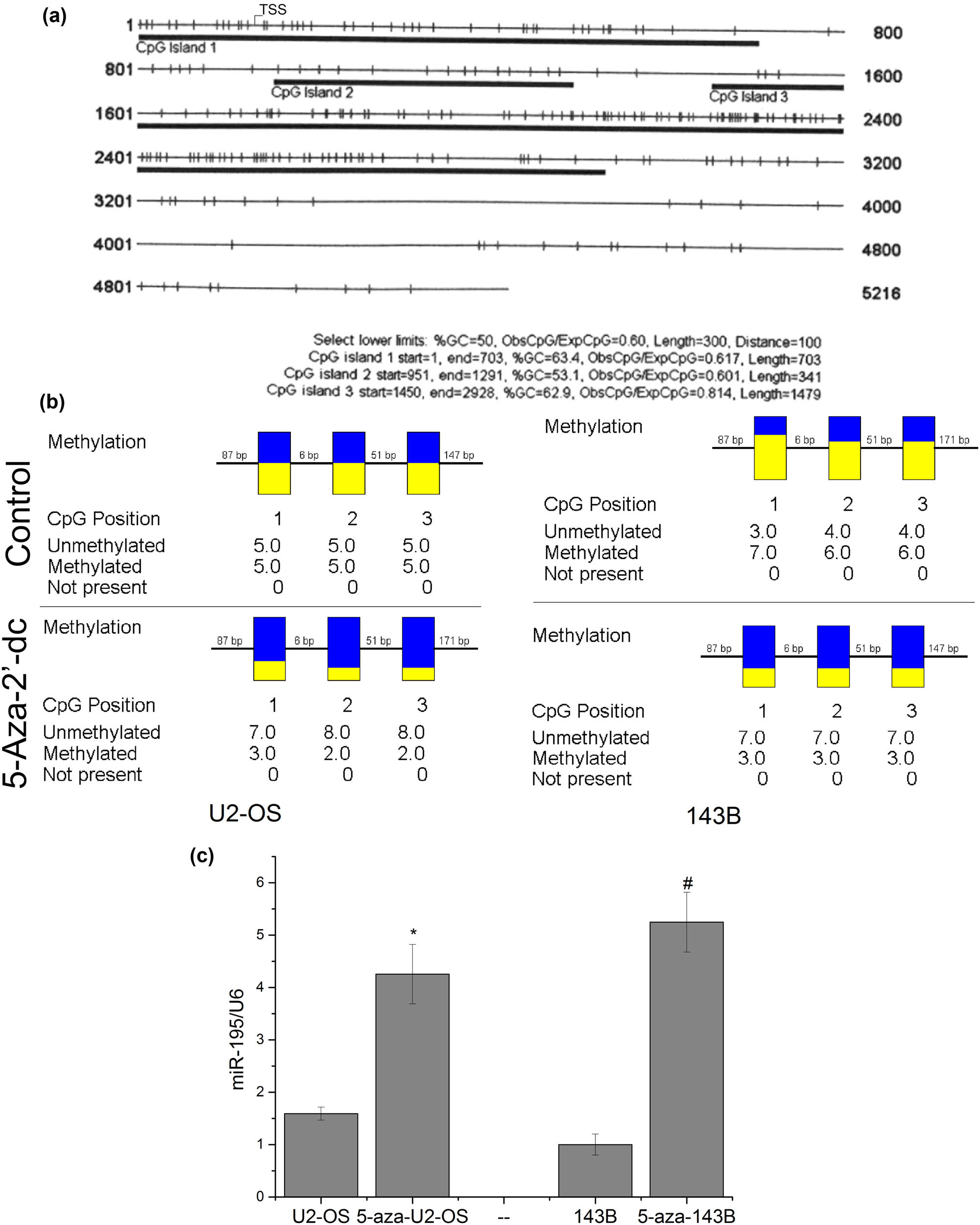
The methylation of miR-195 was one of the reasons why miR-195 decreased. (a) The prediction results showed that the promoter region of miR-195 is full of CPG islands. (b) Bisulfite sequencing PCR (BSP) assay showed that the methylation levels of the miR-195 promoter region were significantly downregulated on cells treated with 5-aza-2′-dc (n = 6). (c) The expression level of miR-195 was significantly upregulated in U2-OS and 143B cells treated with the demethylation reagent in vitro (n = 6). The error bar represents the standard variation of the miR-195 levels in the U2-OS or 143B cells treated with 5-aza.
3.5 Methylation might promote osteosarcoma cell proliferation, migration, and invasion
MTT assay showed that cell proliferation decreased significantly after 5-aza-2′-dc treatment (Figure 5a). The migration ability of cancer cells at 24 h was significantly decreased in U2-OS and 143B cells treated with 5-aza-2′-dc (Figure 5b). Transwell invasion assay demonstrated the invasion ability of cells was significantly decreased in U2-OS and 143B cells after 5-aza-2′-dc treatment (Figure 5c). Taken together, these results suggested that the cell proliferation, migration, and invasion decreased after the cells were treated with the demethylation reagent.
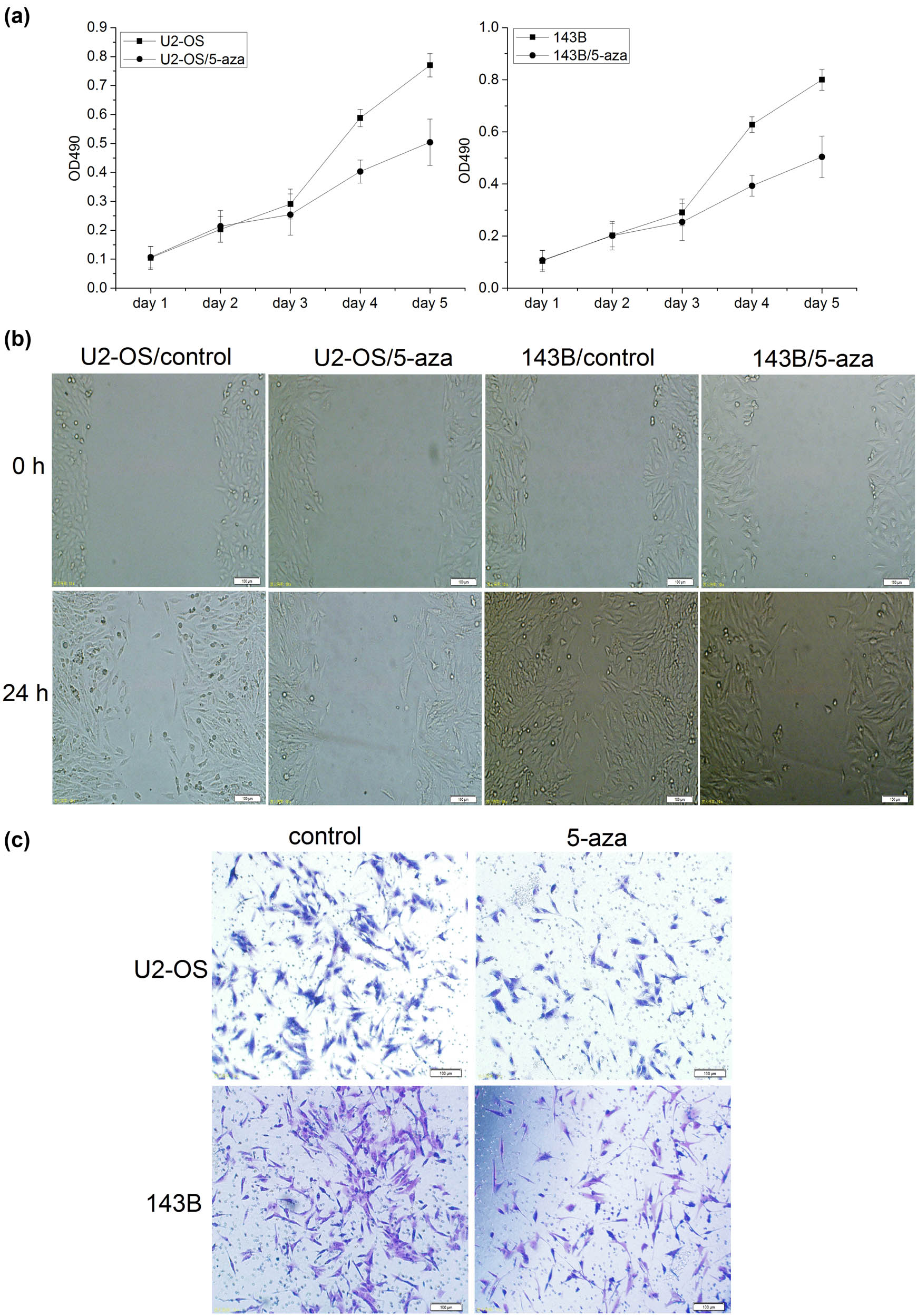
Methylation might promote osteosarcoma cell proliferation, migration, and invasion. (a) The proliferation activity of osteosarcoma cells was detected by MTT assay. MTT assay showed that the proliferation was significantly decreased in U2-OS and 143B cells treated with 5-aza-2′-dc (n = 6, day 4: p < 0.001). The error bar represents the standard variation of the OD490 in the U2-OS or 143B cells treated with 5-aza. (b) The migration ability of osteosarcoma cells at 24 h was significantly decreased in the cells treated with 5-aza-2′-dc (n = 6, ×200). (c) The invasion ability of osteosarcoma cells at 24 h was significantly decreased in the cells treated with 5-aza-2′-dc (n = 6, ×200).
4 Discussion
4.1 Summary
Our previous studies showed FASN was an oncogene. In this study, we first found the expression of FASN was reversely correlated with the expression of miR-195. Then, we demonstrated that FASN was targeted by miR-195, which inhibited osteosarcoma cell proliferation, migration, and invasion. Finally, we used the demethylation reagent to treat the cells and miR-195 increased, indicating that in cancer the expression of miR-195 was suppressed by hypermethylation. The above studies are summarized in Figure 6.
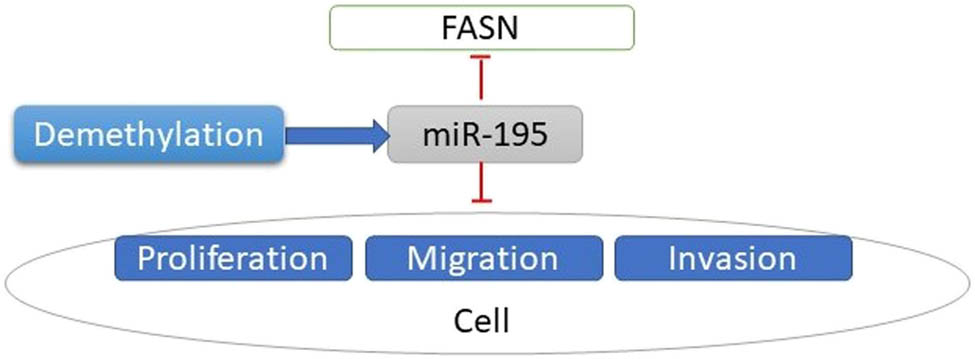
Flow chart of the summary of the main contents. Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma.
4.2 Current therapy of osteosarcoma
Currently, the combination of surgery and multiple chemotherapies is the standardized treatment for osteosarcoma. However, there are low responses to drugs in many patients [26]. Chemotherapy is often administered before surgery to prevent tumors from spreading throughout the body. However, patients with distant metastases still fare poorly, as the 5 year survival rate is about 20% [27,28]. To date, no single genetic target has proven therapeutically successful [7]. Thus, attention should be paid to the underlying molecular mechanisms, which can serve as therapeutic targets. In this study, we identified FASN and miR-195 to be the potential therapeutic targets for osteosarcoma.
4.3 FASN
Nontumor cells naturally acquire palmitate from dietary and quickly store it in triglyceride pools. Palmitate together with other saturated fatty acids has lipotoxicity [29]. To avoid this issue, no-tumor cells change the saturated fatty acids to triglycerides, thereby storing the energy and deactivating the adverse effects [30]. In contrast, tumor cells vigorously synthesize fatty acids and store very little in triglycerides. Cancerous cells typically use these fatty acids to produce phospholipids to build membranes supporting cell proliferation [31].
A high level of fat in diets has been reported in the development of many malignancies [32,33]. Breast cancer cells exhibit high expression levels of FASN and activity [34]. The overexpression of FASN is significantly detected in many types of tumors, including lung, stomach, prostate [35], ovary, bladder, oral cavity, and melanoma [36]. In other words, FASN is expressed at high levels in a variety of human tumors.
Our previous studies show that the positive expression rate of FASN in patients with lung metastasis is 86.4% but only 52.2% in those without lung metastasis [37]. We also found that the expression of FASN was significantly correlated with the tumor size. The FASN levels were higher in tumor diameter larger than 8 cm than in smaller than 8 cm [37]. In this study, based on our previous study on FASN, we further study the miRNA regulating FASN. Specifically, the miR-195 could target and inhibit FASN.
4.4 Functions of FASN in tumors
Tumors have the ability to synthesize their own fatty acid, independent of the regulatory signaling pathways inhibiting the fatty acid synthesis in normal cells [8]. All esterified fatty acids in the tumor are from de novo synthesis [38]. Endogenous fatty acid synthesis is a significant source of fatty acids for the growth of tumor cells [8]. The synthesis of fatty acids is essential for the biogenesis of cellular membranes in rapidly dividing cancer cells [39]. Long-chain fatty acids generated from the palmitate precursors (in turn generated by FASN) are essential for cell division [39,40]. Inhibition of FASN decreased the DNA synthesis and suppressed cells to progress to S-phase [41]. De novo synthesis of membrane phospholipids might be necessary for cell dividing.
The tumors with high expression levels of FASN have increased palmitoylation of Wnt-1 and stabilized β-catenin [29], which might activate the transcription factor. Besides Wnt, tubulin and Harvey rat sarcoma viral oncogene homolog (HRAS) require palmitoylation to localize and function appropriately [42]. In addition, the PI3K/Akt/mTOR pathway and protein kinase C depend on FASN [43]. It is reported that inhibition of FASN results in a decline in diacyl glycerols, those in which at least one acyl chain is palmitate. The decrease of diacyl glycerols leads to a decrease in the protein kinase C pathway, resulting in the death of the tumor cell [44]. Flow cytometry has demonstrated that, due to the inhibition of fatty acid synthesis, cells accumulated in G1, indicating a relation between FASN and cell cycle [45]. Inhibition of FASN leads to an inhibition of S-phase progression and DNA replication [41].
4.5 FASN and metastasis of osteosarcoma
During the synthesis of endogenous fatty acids, the key enzyme, FASN, is responsible for catalyzing the synthesis of long-chain fatty acids in mammals. Also, FASN is critical in sustaining the biological features of malignant tumor cells [46]. In fact, FASN has been studied as a candidate oncogene in cancer [47] such as prostate cancer [48], liver cancer [49], and ovarian cancer [50]. Recently, evidence has shown that fatty acid metabolic pathways play a critical role in carcinogenesis [51]. Inhibition of FASN expression could suppress malignant tumor cell proliferation in vitro and in vivo in oral squamous cell carcinomas [52], liver cancer [53], and neurogenesis [54]. Therefore, FASN has been considered as a promising target for anticancer treatment and management. However, the molecular roles of FASN in osteosarcoma cells remain unclear and need to be further studied.
Increasing evidence showed that FASN also contributes to colorectal cancer cell metastasis [55]. However, whether FASN could promote metastasis in OS and the molecular experimental mechanisms remain unclear. One of the most important reasons for lung metastasis is anoikis resistance [56]. Whether FASN assists lung metastasis of OS by enhancing the anoikis resistance and the detailed molecular and cellular mechanisms need to be elucidated.
-
Funding information: This work was financially supported by the National Key R&D Program of China (2018YFA0703100), NSFC/Research Grants Council (RGC) Joint Research Scheme (N_HKU753/21, 3211101031), National Natural Science Foundation of China (81802959, 81660442, 81860472 and 8156046), Guangdong Basic and Applied Basic Research Foundation (2018A030313888), Shenzhen Science and Technology Funding (JCYJ20200109150420892, JSGG20180507183242702 & JCYJ20170413162104773), HKUSZH Fund for Shenzhen Key Medical Discipline (SZXK2020084), Sanming Project of Medicine in Shenzhen “Team of Excellence in Spinal Deformities and Spinal Degeneration” (SZSM201612055), The University of Hong Kong-Shenzhen Hospital Postdoctoral Fellowship Fund, Natural Science Foundation of Jiangxi Province (No. 20192ACBL21041), and the Technology Research Development and Creative Design Program of Shenzhen, Nanshan District (2020014).
-
Author contributions: Research design: Z.L.; acquisition of data: D.L.; drafting of the article including figures and critical revision of the article: T.S.; and supervision of the project: W.L., T.W., J.W., and Y.F.
-
Conflict of interest: The authors have no conflicts of interest to disclose.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Tang H, Tang Z, Jiang Y, Wei W, Lu J. Pathological and therapeutic aspects of matrix metalloproteinases: Implications in osteosarcoma. Asia Pac J Clin Oncol. 2019;15:218–24.10.1111/ajco.13165Suche in Google Scholar
[2] Alfano L, Costa C, Caporaso A, Antonini D, Giordano A, Pentimalli F. HUR protects NONO from degradation by mir320, which is induced by p53 upon UV irradiation. Oncotarget. 2016;7(47):78127–39.10.18632/oncotarget.13002Suche in Google Scholar
[3] Mialou V, Philip T, Kalifa C, Perol D, Gentet JC, Marec-Berard P, et al. Metastatic osteosarcoma at diagnosis – prognostic factors and long-term outcome – the French pediatric experience. Cancer-Am Cancer Soc. 2005;104(5):1100–9.10.1002/cncr.21263Suche in Google Scholar
[4] Yang CX, Sedhom W, Song J, Lu SL. The role of microRNAs in recurrence and metastasis of head and neck squamous cell carcinoma. Cancers (Basel). 2019;11(3):395.10.3390/cancers11030395Suche in Google Scholar
[5] Aljubran AH, Griffin A, Pintilie M, Blackstein M. Osteosarcoma in adolescents and adults: survival analysis with and without lung metastases. Ann Oncol. 2009;20(6):1136–41.10.1093/annonc/mdn731Suche in Google Scholar
[6] Yang Z, Li X, Yang Y, He Z, Qu X, Zhang Y. Long noncoding RNAs in the progression, metastasis, and prognosis of osteosarcoma. Cell Death Dis. 2016;7(9):e2389.10.1038/cddis.2016.272Suche in Google Scholar
[7] Messerschmitt PJ, Garcia RM, Abdul-Karim FW, Greenfield EM, Getty PJ. Osteosarcoma. J Am Acad Orthop Surg. 2009;17(8):515–27.10.5435/00124635-200908000-00005Suche in Google Scholar
[8] Kuhajda FP. Fatty-acid synthase and human cancer: new perspectives on its role in tumor biology. Nutrition. 2000;16(3):202–8.10.1016/S0899-9007(99)00266-XSuche in Google Scholar
[9] Yu XH, Wang HF, Wu JB, Wang SS, Tang YJ, Tang YL, et al. Non-coding RNAs derailed: the many influences on the fatty acid reprogramming of cancer. Life Sci. 2019;231:116509.10.1016/j.lfs.2019.05.065Suche in Google Scholar PubMed
[10] Hashmi S, Wang Y, Suman DS, Parhar RS, Collison K, Conca W, et al. Human cancer: is it linked to dysfunctional lipid metabolism? Biochim Biophys Acta. 2015;1850(2):352–64.10.1016/j.bbagen.2014.11.004Suche in Google Scholar PubMed
[11] Piva R, Spandidos DA, Gambari R. From microRNA functions to microRNA therapeutics: novel targets and novel drugs in breast cancer research and treatment (Review). Int J Oncol. 2013;43(4):985–94.10.3892/ijo.2013.2059Suche in Google Scholar PubMed PubMed Central
[12] Jonas S, Izaurralde E. Towards a molecular understanding of microRNA-mediated gene silencing. Nat Rev Genet. 2015;16(7):421–33.10.1038/nrg3965Suche in Google Scholar
[13] Sun T, Zhong X, Song H, Liu J, Li J, Leung F, et al. Anoikis resistant mediated by FASN promoted growth and metastasis of osteosarcoma. Cell Death Dis. 2019;10(4):298.10.1038/s41419-019-1532-2Suche in Google Scholar
[14] Sun M, Zhou X, Chen L, Huang S, Leung V, Wu N, et al. The regulatory roles of microRNAs in bone remodeling and perspectives as biomarkers in osteoporosis. Biomed Res Int. 2016;2016:1652417.10.1155/2016/1652417Suche in Google Scholar
[15] Trachana V, Ntoumou E, Anastasopoulou L, Tsezou A. Studying microRNAs in osteoarthritis: critical overview of different analytical approaches. Mech Ageing Dev. 2018;171:15–23.10.1016/j.mad.2018.02.006Suche in Google Scholar
[16] Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function. Cell. 2004;116(2):281–97.10.1016/S0092-8674(04)00045-5Suche in Google Scholar
[17] Bartel DP. MicroRNAs: target recognition and regulatory functions. Cell. 2009;136(2):215–33.10.1016/j.cell.2009.01.002Suche in Google Scholar
[18] Ayub SG, Kaul D, Ayub T. Microdissecting the role of microRNAs in the pathogenesis of prostate cancer. Cancer Genet-Ny. 2015;208(6):289–302.10.1016/j.cancergen.2015.02.010Suche in Google Scholar
[19] Brennecke J, Hipfner DR, Stark A, Russell RB, Cohen SM. Bantam encodes a developmentally regulated microRNA that controls cell proliferation and regulates the proapoptotic gene hid in Drosophila. Cell. 2003;113(1):25–36.10.1016/S0092-8674(03)00231-9Suche in Google Scholar
[20] Xu PZ, Vernooy SY, Guo M, Hay BA. The drosophila microRNA mir-14 suppresses cell death and is required for normal fat metabolism. Curr Biol. 2003;13(9):790–5.10.1016/S0960-9822(03)00250-1Suche in Google Scholar
[21] Ambros V. The functions of animal microRNAs. Nature. 2004;431(7006):350–5.10.1038/nature02871Suche in Google Scholar PubMed
[22] Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin M, Shimizu M, et al. miR-15 and miR-16 induce apoptosis by targeting BCL2. P Natl Acad Sci USA. 2005;102(39):13944–9.10.1073/pnas.0506654102Suche in Google Scholar PubMed PubMed Central
[23] Sun TH, Cheung KSC, Liu ZL, Leung F, Lu WW. Matrix metallopeptidase 9 targeted by hsa-miR-494 promotes silybin-inhibited osteosarcoma. Mol Carcinog. 2018;57(2):262–71.10.1002/mc.22753Suche in Google Scholar PubMed
[24] Sun T, Li CT, Xiong L, Ning Z, Leung F, Peng S, et al. miR-375-3p negatively regulates osteogenesis by targeting and decreasing the expression levels of LRP5 and beta-catenin. Plos One. 2017;12(2):0171281.10.1371/journal.pone.0171281Suche in Google Scholar PubMed PubMed Central
[25] Sun TH, Leung F, Lu WW. miR-9-5p, miR-675-5p and miR-138-5p damages the strontium and LRP5-mediated skeletal cell proliferation, differentiation, and adhesion. International Journal of Molecular Sciences. 2016;17(2):236.10.3390/ijms17020236Suche in Google Scholar PubMed PubMed Central
[26] Chen R, Wang G, Zheng Y, Hua Y, Cai Z. Drug resistance-related microRNAs in osteosarcoma: translating basic evidence into therapeutic strategies. J Cell Mol Med. 2019;23(4):2280–92.10.1111/jcmm.14064Suche in Google Scholar PubMed PubMed Central
[27] Guo J, Reddick WE, Glass JO, Ji Q, Billups CA, Wu J, et al. Dynamic contrast-enhanced magnetic resonance imaging as a prognostic factor in predicting event-free and overall survival in pediatric patients with osteosarcoma. Cancer. 2012;118(15):3776–85.10.1002/cncr.26701Suche in Google Scholar PubMed PubMed Central
[28] Han G, Wang Y, Bi WZ, Wang DJ, Lu SB, Zhang L, et al. Magnetic resonance imaging is appropriate for determining the osteotomy plane for appendicular osteosarcoma after neoadjuvant chemotherapy. Med Oncol. 2012;29(2):1347–53.10.1007/s12032-011-9861-8Suche in Google Scholar PubMed
[29] Buckley D, Duke G, Heuer TS, O'farrell M, Wagman AS, McCulloch W, et al. Fatty acid synthase – modern tumor cell biology insights into a classical oncology target. Pharmacol Ther. 2017;177:23–31.10.1016/j.pharmthera.2017.02.021Suche in Google Scholar PubMed
[30] Listenberger LL, Ory DS, Schaffer JE. Palmitate-induced apoptosis can occur through a ceramide-independent pathway. J Biol Chem. 2001;276(18):14890–5.10.1074/jbc.M010286200Suche in Google Scholar PubMed
[31] Li J, Cheng JX. Direct visualization of de novo lipogenesis in single living cells. Sci Rep. 2014;4:6807.10.1038/srep06807Suche in Google Scholar PubMed PubMed Central
[32] Willett WC, Hunter DJ, Stampfer MJ, Colditz G, Manson JE, Spiegelman D, et al. Dietary fat and fiber in relation to risk of breast cancer. An 8-year follow-up. JAMA. 1992;268(15):2037–44.10.1001/jama.1992.03490150089030Suche in Google Scholar
[33] Risch HA, Jain M, Marrett LD, Howe GR. Dietary fat intake and risk of epithelial ovarian cancer. J Natl Cancer Inst. 1994;86(18):1409–15.10.1093/jnci/86.18.1409Suche in Google Scholar PubMed
[34] Kuhajda FP, Jenner K, Wood FD, Hennigar RA, Jacobs LB, Dick JD, et al. Fatty acid synthesis: a potential selective target for antineoplastic therapy. Proc Natl Acad Sci U S A. 1994;91(14):6379–83.10.1073/pnas.91.14.6379Suche in Google Scholar PubMed PubMed Central
[35] Wen S, Niu Y, Lee SO, Yeh S, Shang Z, Gao H, et al. Targeting fatty acid synthase with ASC-J9 suppresses proliferation and invasion of prostate cancer cells. Mol Carcinogen. 2016;55(12):2278–90.10.1002/mc.22468Suche in Google Scholar PubMed
[36] Zhang JS, Lei JP, Wei GQ, Chen H, Ma CY, Jiang HZ. Natural fatty acid synthase inhibitors as potent therapeutic agents for cancers: a review. Pharm Biol. 2016;54(9):1919–25.10.3109/13880209.2015.1113995Suche in Google Scholar PubMed
[37] Liu ZL, Zhou Y, Luo QF, Hu M, Wang G, Huang SH, et al. Inhibition of fatty acid synthase supresses osteosarcoma cell invasion and migration. Indian J Pathol Microbiol. 2012;55(2):163–9.10.4103/0377-4929.97849Suche in Google Scholar PubMed
[38] Ookhtens M, Kannan R, Lyon I, Baker N. Liver and adipose tissue contributions to newly formed fatty acids in an ascites tumor. Am J Physiol. 1984;247(1 Pt 2):R146–53.10.1152/ajpregu.1984.247.1.R146Suche in Google Scholar PubMed
[39] Baron A, Migita T, Tang D, Loda M. Fatty acid synthase: a metabolic oncogene in prostate cancer? J Cell Biochem. 2004;91(1):47–53.10.1002/jcb.10708Suche in Google Scholar PubMed
[40] Ayton P, Sneddon SF, Palmer DB, Rosewell IR, Owen MJ, Young B, et al. Truncation of the Mll gene in exon 5 by gene targeting leads to early preimplantation lethality of homozygous embryos. Genesis. 2001;30(4):201–12.10.1002/gene.1066Suche in Google Scholar PubMed
[41] Pizer ES, Chrest FJ, DiGiuseppe JA, Han WF. Pharmacological inhibitors of mammalian fatty acid synthase suppress DNA replication and induce apoptosis in tumor cell lines. Cancer Res. 1998;58(20):4611–5.Suche in Google Scholar
[42] Gao X, Hannoush RN. Method for cellular imaging of palmitoylated proteins with clickable probes and proximity ligation applied to Hedgehog, tubulin, and Ras. J Am Chem Soc. 2014;136(12):4544–50.10.1021/ja410068gSuche in Google Scholar PubMed
[43] Jones SF, Infante JR. Molecular pathways: fatty acid synthase. Clin Cancer Res. 2015;21(24):5434–8.10.1158/1078-0432.CCR-15-0126Suche in Google Scholar PubMed
[44] Benjamin DI, Li DS, Lowe W, Heuer T, Kemble G, Nomura DK. Diacylglycerol metabolism and signaling is a driving force underlying FASN inhibitor sensitivity in cancer cells. ACS Chem Biol. 2015;10(7):1616–23.10.1021/acschembio.5b00240Suche in Google Scholar PubMed PubMed Central
[45] Pizer ES, Wood FD, Pasternack GR, Kuhajda FP. Fatty acid synthase (FAS): a target for cytotoxic antimetabolites in HL60 promyelocytic leukemia cells. Cancer Res. 1996;56(4):745–51.Suche in Google Scholar
[46] Hess D, Igal RA. Genistein downregulates de novo lipid synthesis and impairs cell proliferation in human lung cancer cells. Exp Biol Med (Maywood). 2011;236(6):707–13.10.1258/ebm.2011.010265Suche in Google Scholar PubMed
[47] Chen XY, Ruan HB, Long XH, Peng AF, Zhou LD, Liu JM, et al. Blocking fatty acid synthase inhibits tumor progression of human osteosarcoma by regulating the human epidermal growth factor receptor 2/phosphoinositide 3-kinase/protein kinase B signaling pathway in xenograft models. Exp Ther Med. 2017;13(5):2411–6.10.3892/etm.2017.4284Suche in Google Scholar PubMed PubMed Central
[48] Migita T, Ruiz S, Fornari A, Fiorentino M, Priolo C, Zadra G, et al. Fatty acid synthase: a metabolic enzyme and candidate oncogene in prostate cancer. J Natl Cancer Inst. 2009;101(7):519–32.10.1093/jnci/djp030Suche in Google Scholar PubMed PubMed Central
[49] Qiu Z, Zhang C, Zhou J, Hu J, Sheng L, Li X, et al. Celecoxib alleviates AKT/c-Met-triggered rapid hepatocarcinogenesis by suppressing a novel COX-2/AKT/FASN cascade. Mol Carcinog. 2018;58:31–41.10.1002/mc.22904Suche in Google Scholar PubMed
[50] Jiang L, Wang H, Li J, Fang X, Pan H, Yuan X, et al. Up-regulated FASN expression promotes transcoelomic metastasis of ovarian cancer cell through epithelial-mesenchymal transition. Int J Mol Sci. 2014;15(7):11539–54.10.3390/ijms150711539Suche in Google Scholar PubMed PubMed Central
[51] Yeh CS, Wang JY, Cheng TL, Juan CH, Wu CH, Lin SR. Fatty acid metabolism pathway play an important role in carcinogenesis of human colorectal cancers by microarray-bioinformatics analysis. Cancer Lett. 2006;233(2):297–308.10.1016/j.canlet.2005.03.050Suche in Google Scholar PubMed
[52] Agostini M, Almeida LY, Bastos DC, Ortega RM, Moreira FS, Seguin F, et al. The fatty acid synthase inhibitor orlistat reduces the growth and metastasis of orthotopic tongue oral squamous cell carcinomas. Mol Cancer Ther. 2014;13(3):585–95.10.1158/1535-7163.MCT-12-1136Suche in Google Scholar PubMed
[53] Lumetti S, Manfredi E, Ferraris S, Spriano S, Passeri G, Ghiacci G, et al. The response of osteoblastic MC3T3-E1 cells to micro- and nano-textured, hydrophilic and bioactive titanium surfaces. J Mater Sci Mater Med. 2016;27(4):68.10.1007/s10856-016-5678-5Suche in Google Scholar PubMed
[54] Chorna NE, Santos-Soto IJ, Carballeira NM, Morales JL, de la Nuez J, Cátala-Valentin A, et al. Fatty acid synthase as a factor required for exercise-induced cognitive enhancement and dentate gyrus cellular proliferation. Plos One. 2013;8(11):e77845.10.1371/journal.pone.0077845Suche in Google Scholar PubMed PubMed Central
[55] Jafari N, Drury J, Morris AJ, Onono FO, Stevens PD, Gao T, et al. De novo fatty acid synthesis driven sphingolipid metabolism promotes metastatic potential of colorectal cancer. Mol Cancer Res. 2018;17:140–52.10.1158/1541-7786.MCR-18-0199Suche in Google Scholar PubMed PubMed Central
[56] Du L, Han XG, Tu B, Wang MQ, Qiao H, Zhang SH, et al. CXCR1/Akt signaling activation induced by mesenchymal stem cell-derived IL-8 promotes osteosarcoma cell anoikis resistance and pulmonary metastasis. Cell Death Dis. 2018;9(7):714.10.1038/s41419-018-0745-0Suche in Google Scholar PubMed PubMed Central
© 2022 Tianhao Sun et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Research Articles
- AMBRA1 attenuates the proliferation of uveal melanoma cells
- A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma
- Differences in complications between hepatitis B-related cirrhosis and alcohol-related cirrhosis
- Effect of gestational diabetes mellitus on lipid profile: A systematic review and meta-analysis
- Long noncoding RNA NR2F1-AS1 stimulates the tumorigenic behavior of non-small cell lung cancer cells by sponging miR-363-3p to increase SOX4
- Promising novel biomarkers and candidate small-molecule drugs for lung adenocarcinoma: Evidence from bioinformatics analysis of high-throughput data
- Plasmapheresis: Is it a potential alternative treatment for chronic urticaria?
- The biomarkers of key miRNAs and gene targets associated with extranodal NK/T-cell lymphoma
- Gene signature to predict prognostic survival of hepatocellular carcinoma
- Effects of miRNA-199a-5p on cell proliferation and apoptosis of uterine leiomyoma by targeting MED12
- Does diabetes affect paraneoplastic thrombocytosis in colorectal cancer?
- Is there any effect on imprinted genes H19, PEG3, and SNRPN during AOA?
- Leptin and PCSK9 concentrations are associated with vascular endothelial cytokines in patients with stable coronary heart disease
- Pericentric inversion of chromosome 6 and male fertility problems
- Staple line reinforcement with nebulized cyanoacrylate glue in laparoscopic sleeve gastrectomy: A propensity score-matched study
- Retrospective analysis of crescent score in clinical prognosis of IgA nephropathy
- Expression of DNM3 is associated with good outcome in colorectal cancer
- Activation of SphK2 contributes to adipocyte-induced EOC cell proliferation
- CRRT influences PICCO measurements in febrile critically ill patients
- SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis
- lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells
- circ_AKT3 knockdown suppresses cisplatin resistance in gastric cancer
- Prognostic value of nicotinamide N-methyltransferase in human cancers: Evidence from a meta-analysis and database validation
- GPC2 deficiency inhibits cell growth and metastasis in colon adenocarcinoma
- A pan-cancer analysis of the oncogenic role of Holliday junction recognition protein in human tumors
- Radiation increases COL1A1, COL3A1, and COL1A2 expression in breast cancer
- Association between preventable risk factors and metabolic syndrome
- miR-29c-5p knockdown reduces inflammation and blood–brain barrier disruption by upregulating LRP6
- Cardiac contractility modulation ameliorates myocardial metabolic remodeling in a rabbit model of chronic heart failure through activation of AMPK and PPAR-α pathway
- Quercitrin protects human bronchial epithelial cells from oxidative damage
- Smurf2 suppresses the metastasis of hepatocellular carcinoma via ubiquitin degradation of Smad2
- circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury
- Sonoclot’s usefulness in prediction of cardiopulmonary arrest prognosis: A proof of concept study
- Four drug metabolism-related subgroups of pancreatic adenocarcinoma in prognosis, immune infiltration, and gene mutation
- Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma
- LMO3 promotes proliferation and metastasis of papillary thyroid carcinoma cells by regulating LIMK1-mediated cofilin and the β-catenin pathway
- Cx43 upregulation in HUVECs under stretch via TGF-β1 and cytoskeletal network
- Evaluation of menstrual irregularities after COVID-19 vaccination: Results of the MECOVAC survey
- Histopathologic findings on removed stomach after sleeve gastrectomy. Do they influence the outcome?
- Analysis of the expression and prognostic value of MT1-MMP, β1-integrin and YAP1 in glioma
- Optimal diagnosis of the skin cancer using a hybrid deep neural network and grasshopper optimization algorithm
- miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells
- Clinical value of SIRT1 as a prognostic biomarker in esophageal squamous cell carcinoma, a systematic meta-analysis
- circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8
- miR-22-5p regulates the self-renewal of spermatogonial stem cells by targeting EZH2
- hsa-miR-340-5p inhibits epithelial–mesenchymal transition in endometriosis by targeting MAP3K2 and inactivating MAPK/ERK signaling
- circ_0085296 inhibits the biological functions of trophoblast cells to promote the progression of preeclampsia via the miR-942-5p/THBS2 network
- TCD hemodynamics findings in the subacute phase of anterior circulation stroke patients treated with mechanical thrombectomy
- Development of a risk-stratification scoring system for predicting risk of breast cancer based on non-alcoholic fatty liver disease, non-alcoholic fatty pancreas disease, and uric acid
- Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway
- circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis
- Human amniotic fluid as a source of stem cells
- lncRNA NONRATT013819.2 promotes transforming growth factor-β1-induced myofibroblastic transition of hepatic stellate cells by miR24-3p/lox
- NORAD modulates miR-30c-5p-LDHA to protect lung endothelial cells damage
- Idiopathic pulmonary fibrosis telemedicine management during COVID-19 outbreak
- Risk factors for adverse drug reactions associated with clopidogrel therapy
- Serum zinc associated with immunity and inflammatory markers in Covid-19
- The relationship between night shift work and breast cancer incidence: A systematic review and meta-analysis of observational studies
- LncRNA expression in idiopathic achalasia: New insight and preliminary exploration into pathogenesis
- Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway
- Moscatilin suppresses the inflammation from macrophages and T cells
- Zoledronate promotes ECM degradation and apoptosis via Wnt/β-catenin
- Epithelial-mesenchymal transition-related genes in coronary artery disease
- The effect evaluation of traditional vaginal surgery and transvaginal mesh surgery for severe pelvic organ prolapse: 5 years follow-up
- Repeated partial splenic artery embolization for hypersplenism improves platelet count
- Low expression of miR-27b in serum exosomes of non-small cell lung cancer facilitates its progression by affecting EGFR
- Exosomal hsa_circ_0000519 modulates the NSCLC cell growth and metastasis via miR-1258/RHOV axis
- miR-455-5p enhances 5-fluorouracil sensitivity in colorectal cancer cells by targeting PIK3R1 and DEPDC1
- The effect of tranexamic acid on the reduction of intraoperative and postoperative blood loss and thromboembolic risk in patients with hip fracture
- Isocitrate dehydrogenase 1 mutation in cholangiocarcinoma impairs tumor progression by sensitizing cells to ferroptosis
- Artemisinin protects against cerebral ischemia and reperfusion injury via inhibiting the NF-κB pathway
- A 16-gene signature associated with homologous recombination deficiency for prognosis prediction in patients with triple-negative breast cancer
- Lidocaine ameliorates chronic constriction injury-induced neuropathic pain through regulating M1/M2 microglia polarization
- MicroRNA 322-5p reduced neuronal inflammation via the TLR4/TRAF6/NF-κB axis in a rat epilepsy model
- miR-1273h-5p suppresses CXCL12 expression and inhibits gastric cancer cell invasion and metastasis
- Clinical characteristics of pneumonia patients of long course of illness infected with SARS-CoV-2
- circRNF20 aggravates the malignancy of retinoblastoma depending on the regulation of miR-132-3p/PAX6 axis
- Linezolid for resistant Gram-positive bacterial infections in children under 12 years: A meta-analysis
- Rack1 regulates pro-inflammatory cytokines by NF-κB in diabetic nephropathy
- Comprehensive analysis of molecular mechanism and a novel prognostic signature based on small nuclear RNA biomarkers in gastric cancer patients
- Smog and risk of maternal and fetal birth outcomes: A retrospective study in Baoding, China
- Let-7i-3p inhibits the cell cycle, proliferation, invasion, and migration of colorectal cancer cells via downregulating CCND1
- β2-Adrenergic receptor expression in subchondral bone of patients with varus knee osteoarthritis
- Possible impact of COVID-19 pandemic and lockdown on suicide behavior among patients in Southeast Serbia
- In vitro antimicrobial activity of ozonated oil in liposome eyedrop against multidrug-resistant bacteria
- Potential biomarkers for inflammatory response in acute lung injury
- A low serum uric acid concentration predicts a poor prognosis in adult patients with candidemia
- Antitumor activity of recombinant oncolytic vaccinia virus with human IL2
- ALKBH5 inhibits TNF-α-induced apoptosis of HUVECs through Bcl-2 pathway
- Risk prediction of cardiovascular disease using machine learning classifiers
- Value of ultrasonography parameters in diagnosing polycystic ovary syndrome
- Bioinformatics analysis reveals three key genes and four survival genes associated with youth-onset NSCLC
- Identification of autophagy-related biomarkers in patients with pulmonary arterial hypertension based on bioinformatics analysis
- Protective effects of glaucocalyxin A on the airway of asthmatic mice
- Overexpression of miR-100-5p inhibits papillary thyroid cancer progression via targeting FZD8
- Bioinformatics-based analysis of SUMOylation-related genes in hepatocellular carcinoma reveals a role of upregulated SAE1 in promoting cell proliferation
- Effectiveness and clinical benefits of new anti-diabetic drugs: A real life experience
- Identification of osteoporosis based on gene biomarkers using support vector machine
- Tanshinone IIA reverses oxaliplatin resistance in colorectal cancer through microRNA-30b-5p/AVEN axis
- miR-212-5p inhibits nasopharyngeal carcinoma progression by targeting METTL3
- Association of ST-T changes with all-cause mortality among patients with peripheral T-cell lymphomas
- LINC00665/miRNAs axis-mediated collagen type XI alpha 1 correlates with immune infiltration and malignant phenotypes in lung adenocarcinoma
- The perinatal factors that influence the excretion of fecal calprotectin in premature-born children
- Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study
- Does the use of 3D-printed cones give a chance to postpone the use of megaprostheses in patients with large bone defects in the knee joint?
- lncRNA HAGLR modulates myocardial ischemia–reperfusion injury in mice through regulating miR-133a-3p/MAPK1 axis
- Protective effect of ghrelin on intestinal I/R injury in rats
- In vivo knee kinematics of an innovative prosthesis design
- Relationship between the height of fibular head and the incidence and severity of knee osteoarthritis
- lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury during cerebral ischemic stroke via miR-186-5p/XIAP axis
- Correlation of cardiac troponin T and APACHE III score with all-cause in-hospital mortality in critically ill patients with acute pulmonary embolism
- LncRNA LINC01857 reduces metastasis and angiogenesis in breast cancer cells via regulating miR-2052/CENPQ axis
- Endothelial cell-specific molecule 1 (ESM1) promoted by transcription factor SPI1 acts as an oncogene to modulate the malignant phenotype of endometrial cancer
- SELENBP1 inhibits progression of colorectal cancer by suppressing epithelial–mesenchymal transition
- Visfatin is negatively associated with coronary artery lesions in subjects with impaired fasting glucose
- Treatment and outcomes of mechanical complications of acute myocardial infarction during the Covid-19 era: A comparison with the pre-Covid-19 period. A systematic review and meta-analysis
- Neonatal stroke surveillance study protocol in the United Kingdom and Republic of Ireland
- Oncogenic role of TWF2 in human tumors: A pan-cancer analysis
- Mean corpuscular hemoglobin predicts the length of hospital stay independent of severity classification in patients with acute pancreatitis
- Association of gallstone and polymorphisms of UGT1A1*27 and UGT1A1*28 in patients with hepatitis B virus-related liver failure
- TGF-β1 upregulates Sar1a expression and induces procollagen-I secretion in hypertrophic scarring fibroblasts
- Antisense lncRNA PCNA-AS1 promotes esophageal squamous cell carcinoma progression through the miR-2467-3p/PCNA axis
- NK-cell dysfunction of acute myeloid leukemia in relation to the renin–angiotensin system and neurotransmitter genes
- The effect of dilution with glucose and prolonged injection time on dexamethasone-induced perineal irritation – A randomized controlled trial
- miR-146-5p restrains calcification of vascular smooth muscle cells by suppressing TRAF6
- Role of lncRNA MIAT/miR-361-3p/CCAR2 in prostate cancer cells
- lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2
- Noninvasive diagnosis of AIH/PBC overlap syndrome based on prediction models
- lncRNA FAM230B is highly expressed in colorectal cancer and suppresses the maturation of miR-1182 to increase cell proliferation
- circ-LIMK1 regulates cisplatin resistance in lung adenocarcinoma by targeting miR-512-5p/HMGA1 axis
- LncRNA SNHG3 promoted cell proliferation, migration, and metastasis of esophageal squamous cell carcinoma via regulating miR-151a-3p/PFN2 axis
- Risk perception and affective state on work exhaustion in obstetrics during the COVID-19 pandemic
- lncRNA-AC130710/miR-129-5p/mGluR1 axis promote migration and invasion by activating PKCα-MAPK signal pathway in melanoma
- SNRPB promotes cell cycle progression in thyroid carcinoma via inhibiting p53
- Xylooligosaccharides and aerobic training regulate metabolism and behavior in rats with streptozotocin-induced type 1 diabetes
- Serpin family A member 1 is an oncogene in glioma and its translation is enhanced by NAD(P)H quinone dehydrogenase 1 through RNA-binding activity
- Silencing of CPSF7 inhibits the proliferation, migration, and invasion of lung adenocarcinoma cells by blocking the AKT/mTOR signaling pathway
- Ultrasound-guided lumbar plexus block versus transversus abdominis plane block for analgesia in children with hip dislocation: A double-blind, randomized trial
- Relationship of plasma MBP and 8-oxo-dG with brain damage in preterm
- Identification of a novel necroptosis-associated miRNA signature for predicting the prognosis in head and neck squamous cell carcinoma
- Delayed femoral vein ligation reduces operative time and blood loss during hip disarticulation in patients with extremity tumors
- The expression of ASAP3 and NOTCH3 and the clinicopathological characteristics of adult glioma patients
- Longitudinal analysis of factors related to Helicobacter pylori infection in Chinese adults
- HOXA10 enhances cell proliferation and suppresses apoptosis in esophageal cancer via activating p38/ERK signaling pathway
- Meta-analysis of early-life antibiotic use and allergic rhinitis
- Marital status and its correlation with age, race, and gender in prognosis of tonsil squamous cell carcinomas
- HPV16 E6E7 up-regulates KIF2A expression by activating JNK/c-Jun signal, is beneficial to migration and invasion of cervical cancer cells
- Amino acid profiles in the tissue and serum of patients with liver cancer
- Pain in critically ill COVID-19 patients: An Italian retrospective study
- Immunohistochemical distribution of Bcl-2 and p53 apoptotic markers in acetamiprid-induced nephrotoxicity
- Estradiol pretreatment in GnRH antagonist protocol for IVF/ICSI treatment
- Long non-coding RNAs LINC00689 inhibits the apoptosis of human nucleus pulposus cells via miR-3127-5p/ATG7 axis-mediated autophagy
- The relationship between oxygen therapy, drug therapy, and COVID-19 mortality
- Monitoring hypertensive disorders in pregnancy to prevent preeclampsia in pregnant women of advanced maternal age: Trial mimicking with retrospective data
- SETD1A promotes the proliferation and glycolysis of nasopharyngeal carcinoma cells by activating the PI3K/Akt pathway
- The role of Shunaoxin pills in the treatment of chronic cerebral hypoperfusion and its main pharmacodynamic components
- TET3 governs malignant behaviors and unfavorable prognosis of esophageal squamous cell carcinoma by activating the PI3K/AKT/GSK3β/β-catenin pathway
- Associations between morphokinetic parameters of temporary-arrest embryos and the clinical prognosis in FET cycles
- Long noncoding RNA WT1-AS regulates trophoblast proliferation, migration, and invasion via the microRNA-186-5p/CADM2 axis
- The incidence of bronchiectasis in chronic obstructive pulmonary disease
- Integrated bioinformatics analysis shows integrin alpha 3 is a prognostic biomarker for pancreatic cancer
- Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway
- Comparison of hospitalized patients with severe pneumonia caused by COVID-19 and influenza A (H7N9 and H1N1): A retrospective study from a designated hospital
- lncRNA ZFAS1 promotes intervertebral disc degeneration by upregulating AAK1
- Pathological characteristics of liver injury induced by N,N-dimethylformamide: From humans to animal models
- lncRNA ELFN1-AS1 enhances the progression of colon cancer by targeting miR-4270 to upregulate AURKB
- DARS-AS1 modulates cell proliferation and migration of gastric cancer cells by regulating miR-330-3p/NAT10 axis
- Dezocine inhibits cell proliferation, migration, and invasion by targeting CRABP2 in ovarian cancer
- MGST1 alleviates the oxidative stress of trophoblast cells induced by hypoxia/reoxygenation and promotes cell proliferation, migration, and invasion by activating the PI3K/AKT/mTOR pathway
- Bifidobacterium lactis Probio-M8 ameliorated the symptoms of type 2 diabetes mellitus mice by changing ileum FXR-CYP7A1
- circRNA DENND1B inhibits tumorigenicity of clear cell renal cell carcinoma via miR-122-5p/TIMP2 axis
- EphA3 targeted by miR-3666 contributes to melanoma malignancy via activating ERK1/2 and p38 MAPK pathways
- Pacemakers and methylprednisolone pulse therapy in immune-related myocarditis concomitant with complete heart block
- miRNA-130a-3p targets sphingosine-1-phosphate receptor 1 to activate the microglial and astrocytes and to promote neural injury under the high glucose condition
- Review Articles
- Current management of cancer pain in Italy: Expert opinion paper
- Hearing loss and brain disorders: A review of multiple pathologies
- The rationale for using low-molecular weight heparin in the therapy of symptomatic COVID-19 patients
- Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2
- Interleukin-35 in autoimmune dermatoses: Current concepts
- Recent discoveries in microbiota dysbiosis, cholangiocytic factors, and models for studying the pathogenesis of primary sclerosing cholangitis
- Advantages of ketamine in pediatric anesthesia
- Congenital adrenal hyperplasia. Role of dentist in early diagnosis
- Migraine management: Non-pharmacological points for patients and health care professionals
- Atherogenic index of plasma and coronary artery disease: A systematic review
- Physiological and modulatory role of thioredoxins in the cellular function
- Case Reports
- Intrauterine Bakri balloon tamponade plus cervical cerclage for the prevention and treatment of postpartum haemorrhage in late pregnancy complicated with acute aortic dissection: Case series
- A case of successful pembrolizumab monotherapy in a patient with advanced lung adenocarcinoma: Use of multiple biomarkers in combination for clinical practice
- Unusual neurological manifestations of bilateral medial medullary infarction: A case report
- Atypical symptoms of malignant hyperthermia: A rare causative mutation in the RYR1 gene
- A case report of dermatomyositis with the missed diagnosis of non-small cell lung cancer and concurrence of pulmonary tuberculosis
- A rare case of endometrial polyp complicated with uterine inversion: A case report and clinical management
- Spontaneous rupturing of splenic artery aneurysm: Another reason for fatal syncope and shock (Case report and literature review)
- Fungal infection mimicking COVID-19 infection – A case report
- Concurrent aspergillosis and cystic pulmonary metastases in a patient with tongue squamous cell carcinoma
- Paraganglioma-induced inverted takotsubo-like cardiomyopathy leading to cardiogenic shock successfully treated with extracorporeal membrane oxygenation
- Lineage switch from lymphoma to myeloid neoplasms: First case series from a single institution
- Trismus during tracheal extubation as a complication of general anaesthesia – A case report
- Simultaneous treatment of a pubovesical fistula and lymph node metastasis secondary to multimodal treatment for prostate cancer: Case report and review of the literature
- Two case reports of skin vasculitis following the COVID-19 immunization
- Ureteroiliac fistula after oncological surgery: Case report and review of the literature
- Synchronous triple primary malignant tumours in the bladder, prostate, and lung harbouring TP53 and MEK1 mutations accompanied with severe cardiovascular diseases: A case report
- Huge mucinous cystic neoplasms with adhesion to the left colon: A case report and literature review
- Commentary
- Commentary on “Clinicopathological features of programmed cell death-ligand 1 expression in patients with oral squamous cell carcinoma”
- Rapid Communication
- COVID-19 fear, post-traumatic stress, growth, and the role of resilience
- Erratum
- Erratum to “Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway”
- Erratum to “Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study”
- Erratum to “lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2”
- Retraction
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Retraction to “miR-519d downregulates LEP expression to inhibit preeclampsia development”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part II
- Usefulness of close surveillance for rectal cancer patients after neoadjuvant chemoradiotherapy
Artikel in diesem Heft
- Research Articles
- AMBRA1 attenuates the proliferation of uveal melanoma cells
- A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma
- Differences in complications between hepatitis B-related cirrhosis and alcohol-related cirrhosis
- Effect of gestational diabetes mellitus on lipid profile: A systematic review and meta-analysis
- Long noncoding RNA NR2F1-AS1 stimulates the tumorigenic behavior of non-small cell lung cancer cells by sponging miR-363-3p to increase SOX4
- Promising novel biomarkers and candidate small-molecule drugs for lung adenocarcinoma: Evidence from bioinformatics analysis of high-throughput data
- Plasmapheresis: Is it a potential alternative treatment for chronic urticaria?
- The biomarkers of key miRNAs and gene targets associated with extranodal NK/T-cell lymphoma
- Gene signature to predict prognostic survival of hepatocellular carcinoma
- Effects of miRNA-199a-5p on cell proliferation and apoptosis of uterine leiomyoma by targeting MED12
- Does diabetes affect paraneoplastic thrombocytosis in colorectal cancer?
- Is there any effect on imprinted genes H19, PEG3, and SNRPN during AOA?
- Leptin and PCSK9 concentrations are associated with vascular endothelial cytokines in patients with stable coronary heart disease
- Pericentric inversion of chromosome 6 and male fertility problems
- Staple line reinforcement with nebulized cyanoacrylate glue in laparoscopic sleeve gastrectomy: A propensity score-matched study
- Retrospective analysis of crescent score in clinical prognosis of IgA nephropathy
- Expression of DNM3 is associated with good outcome in colorectal cancer
- Activation of SphK2 contributes to adipocyte-induced EOC cell proliferation
- CRRT influences PICCO measurements in febrile critically ill patients
- SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis
- lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells
- circ_AKT3 knockdown suppresses cisplatin resistance in gastric cancer
- Prognostic value of nicotinamide N-methyltransferase in human cancers: Evidence from a meta-analysis and database validation
- GPC2 deficiency inhibits cell growth and metastasis in colon adenocarcinoma
- A pan-cancer analysis of the oncogenic role of Holliday junction recognition protein in human tumors
- Radiation increases COL1A1, COL3A1, and COL1A2 expression in breast cancer
- Association between preventable risk factors and metabolic syndrome
- miR-29c-5p knockdown reduces inflammation and blood–brain barrier disruption by upregulating LRP6
- Cardiac contractility modulation ameliorates myocardial metabolic remodeling in a rabbit model of chronic heart failure through activation of AMPK and PPAR-α pathway
- Quercitrin protects human bronchial epithelial cells from oxidative damage
- Smurf2 suppresses the metastasis of hepatocellular carcinoma via ubiquitin degradation of Smad2
- circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury
- Sonoclot’s usefulness in prediction of cardiopulmonary arrest prognosis: A proof of concept study
- Four drug metabolism-related subgroups of pancreatic adenocarcinoma in prognosis, immune infiltration, and gene mutation
- Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma
- LMO3 promotes proliferation and metastasis of papillary thyroid carcinoma cells by regulating LIMK1-mediated cofilin and the β-catenin pathway
- Cx43 upregulation in HUVECs under stretch via TGF-β1 and cytoskeletal network
- Evaluation of menstrual irregularities after COVID-19 vaccination: Results of the MECOVAC survey
- Histopathologic findings on removed stomach after sleeve gastrectomy. Do they influence the outcome?
- Analysis of the expression and prognostic value of MT1-MMP, β1-integrin and YAP1 in glioma
- Optimal diagnosis of the skin cancer using a hybrid deep neural network and grasshopper optimization algorithm
- miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells
- Clinical value of SIRT1 as a prognostic biomarker in esophageal squamous cell carcinoma, a systematic meta-analysis
- circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8
- miR-22-5p regulates the self-renewal of spermatogonial stem cells by targeting EZH2
- hsa-miR-340-5p inhibits epithelial–mesenchymal transition in endometriosis by targeting MAP3K2 and inactivating MAPK/ERK signaling
- circ_0085296 inhibits the biological functions of trophoblast cells to promote the progression of preeclampsia via the miR-942-5p/THBS2 network
- TCD hemodynamics findings in the subacute phase of anterior circulation stroke patients treated with mechanical thrombectomy
- Development of a risk-stratification scoring system for predicting risk of breast cancer based on non-alcoholic fatty liver disease, non-alcoholic fatty pancreas disease, and uric acid
- Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway
- circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis
- Human amniotic fluid as a source of stem cells
- lncRNA NONRATT013819.2 promotes transforming growth factor-β1-induced myofibroblastic transition of hepatic stellate cells by miR24-3p/lox
- NORAD modulates miR-30c-5p-LDHA to protect lung endothelial cells damage
- Idiopathic pulmonary fibrosis telemedicine management during COVID-19 outbreak
- Risk factors for adverse drug reactions associated with clopidogrel therapy
- Serum zinc associated with immunity and inflammatory markers in Covid-19
- The relationship between night shift work and breast cancer incidence: A systematic review and meta-analysis of observational studies
- LncRNA expression in idiopathic achalasia: New insight and preliminary exploration into pathogenesis
- Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway
- Moscatilin suppresses the inflammation from macrophages and T cells
- Zoledronate promotes ECM degradation and apoptosis via Wnt/β-catenin
- Epithelial-mesenchymal transition-related genes in coronary artery disease
- The effect evaluation of traditional vaginal surgery and transvaginal mesh surgery for severe pelvic organ prolapse: 5 years follow-up
- Repeated partial splenic artery embolization for hypersplenism improves platelet count
- Low expression of miR-27b in serum exosomes of non-small cell lung cancer facilitates its progression by affecting EGFR
- Exosomal hsa_circ_0000519 modulates the NSCLC cell growth and metastasis via miR-1258/RHOV axis
- miR-455-5p enhances 5-fluorouracil sensitivity in colorectal cancer cells by targeting PIK3R1 and DEPDC1
- The effect of tranexamic acid on the reduction of intraoperative and postoperative blood loss and thromboembolic risk in patients with hip fracture
- Isocitrate dehydrogenase 1 mutation in cholangiocarcinoma impairs tumor progression by sensitizing cells to ferroptosis
- Artemisinin protects against cerebral ischemia and reperfusion injury via inhibiting the NF-κB pathway
- A 16-gene signature associated with homologous recombination deficiency for prognosis prediction in patients with triple-negative breast cancer
- Lidocaine ameliorates chronic constriction injury-induced neuropathic pain through regulating M1/M2 microglia polarization
- MicroRNA 322-5p reduced neuronal inflammation via the TLR4/TRAF6/NF-κB axis in a rat epilepsy model
- miR-1273h-5p suppresses CXCL12 expression and inhibits gastric cancer cell invasion and metastasis
- Clinical characteristics of pneumonia patients of long course of illness infected with SARS-CoV-2
- circRNF20 aggravates the malignancy of retinoblastoma depending on the regulation of miR-132-3p/PAX6 axis
- Linezolid for resistant Gram-positive bacterial infections in children under 12 years: A meta-analysis
- Rack1 regulates pro-inflammatory cytokines by NF-κB in diabetic nephropathy
- Comprehensive analysis of molecular mechanism and a novel prognostic signature based on small nuclear RNA biomarkers in gastric cancer patients
- Smog and risk of maternal and fetal birth outcomes: A retrospective study in Baoding, China
- Let-7i-3p inhibits the cell cycle, proliferation, invasion, and migration of colorectal cancer cells via downregulating CCND1
- β2-Adrenergic receptor expression in subchondral bone of patients with varus knee osteoarthritis
- Possible impact of COVID-19 pandemic and lockdown on suicide behavior among patients in Southeast Serbia
- In vitro antimicrobial activity of ozonated oil in liposome eyedrop against multidrug-resistant bacteria
- Potential biomarkers for inflammatory response in acute lung injury
- A low serum uric acid concentration predicts a poor prognosis in adult patients with candidemia
- Antitumor activity of recombinant oncolytic vaccinia virus with human IL2
- ALKBH5 inhibits TNF-α-induced apoptosis of HUVECs through Bcl-2 pathway
- Risk prediction of cardiovascular disease using machine learning classifiers
- Value of ultrasonography parameters in diagnosing polycystic ovary syndrome
- Bioinformatics analysis reveals three key genes and four survival genes associated with youth-onset NSCLC
- Identification of autophagy-related biomarkers in patients with pulmonary arterial hypertension based on bioinformatics analysis
- Protective effects of glaucocalyxin A on the airway of asthmatic mice
- Overexpression of miR-100-5p inhibits papillary thyroid cancer progression via targeting FZD8
- Bioinformatics-based analysis of SUMOylation-related genes in hepatocellular carcinoma reveals a role of upregulated SAE1 in promoting cell proliferation
- Effectiveness and clinical benefits of new anti-diabetic drugs: A real life experience
- Identification of osteoporosis based on gene biomarkers using support vector machine
- Tanshinone IIA reverses oxaliplatin resistance in colorectal cancer through microRNA-30b-5p/AVEN axis
- miR-212-5p inhibits nasopharyngeal carcinoma progression by targeting METTL3
- Association of ST-T changes with all-cause mortality among patients with peripheral T-cell lymphomas
- LINC00665/miRNAs axis-mediated collagen type XI alpha 1 correlates with immune infiltration and malignant phenotypes in lung adenocarcinoma
- The perinatal factors that influence the excretion of fecal calprotectin in premature-born children
- Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study
- Does the use of 3D-printed cones give a chance to postpone the use of megaprostheses in patients with large bone defects in the knee joint?
- lncRNA HAGLR modulates myocardial ischemia–reperfusion injury in mice through regulating miR-133a-3p/MAPK1 axis
- Protective effect of ghrelin on intestinal I/R injury in rats
- In vivo knee kinematics of an innovative prosthesis design
- Relationship between the height of fibular head and the incidence and severity of knee osteoarthritis
- lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury during cerebral ischemic stroke via miR-186-5p/XIAP axis
- Correlation of cardiac troponin T and APACHE III score with all-cause in-hospital mortality in critically ill patients with acute pulmonary embolism
- LncRNA LINC01857 reduces metastasis and angiogenesis in breast cancer cells via regulating miR-2052/CENPQ axis
- Endothelial cell-specific molecule 1 (ESM1) promoted by transcription factor SPI1 acts as an oncogene to modulate the malignant phenotype of endometrial cancer
- SELENBP1 inhibits progression of colorectal cancer by suppressing epithelial–mesenchymal transition
- Visfatin is negatively associated with coronary artery lesions in subjects with impaired fasting glucose
- Treatment and outcomes of mechanical complications of acute myocardial infarction during the Covid-19 era: A comparison with the pre-Covid-19 period. A systematic review and meta-analysis
- Neonatal stroke surveillance study protocol in the United Kingdom and Republic of Ireland
- Oncogenic role of TWF2 in human tumors: A pan-cancer analysis
- Mean corpuscular hemoglobin predicts the length of hospital stay independent of severity classification in patients with acute pancreatitis
- Association of gallstone and polymorphisms of UGT1A1*27 and UGT1A1*28 in patients with hepatitis B virus-related liver failure
- TGF-β1 upregulates Sar1a expression and induces procollagen-I secretion in hypertrophic scarring fibroblasts
- Antisense lncRNA PCNA-AS1 promotes esophageal squamous cell carcinoma progression through the miR-2467-3p/PCNA axis
- NK-cell dysfunction of acute myeloid leukemia in relation to the renin–angiotensin system and neurotransmitter genes
- The effect of dilution with glucose and prolonged injection time on dexamethasone-induced perineal irritation – A randomized controlled trial
- miR-146-5p restrains calcification of vascular smooth muscle cells by suppressing TRAF6
- Role of lncRNA MIAT/miR-361-3p/CCAR2 in prostate cancer cells
- lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2
- Noninvasive diagnosis of AIH/PBC overlap syndrome based on prediction models
- lncRNA FAM230B is highly expressed in colorectal cancer and suppresses the maturation of miR-1182 to increase cell proliferation
- circ-LIMK1 regulates cisplatin resistance in lung adenocarcinoma by targeting miR-512-5p/HMGA1 axis
- LncRNA SNHG3 promoted cell proliferation, migration, and metastasis of esophageal squamous cell carcinoma via regulating miR-151a-3p/PFN2 axis
- Risk perception and affective state on work exhaustion in obstetrics during the COVID-19 pandemic
- lncRNA-AC130710/miR-129-5p/mGluR1 axis promote migration and invasion by activating PKCα-MAPK signal pathway in melanoma
- SNRPB promotes cell cycle progression in thyroid carcinoma via inhibiting p53
- Xylooligosaccharides and aerobic training regulate metabolism and behavior in rats with streptozotocin-induced type 1 diabetes
- Serpin family A member 1 is an oncogene in glioma and its translation is enhanced by NAD(P)H quinone dehydrogenase 1 through RNA-binding activity
- Silencing of CPSF7 inhibits the proliferation, migration, and invasion of lung adenocarcinoma cells by blocking the AKT/mTOR signaling pathway
- Ultrasound-guided lumbar plexus block versus transversus abdominis plane block for analgesia in children with hip dislocation: A double-blind, randomized trial
- Relationship of plasma MBP and 8-oxo-dG with brain damage in preterm
- Identification of a novel necroptosis-associated miRNA signature for predicting the prognosis in head and neck squamous cell carcinoma
- Delayed femoral vein ligation reduces operative time and blood loss during hip disarticulation in patients with extremity tumors
- The expression of ASAP3 and NOTCH3 and the clinicopathological characteristics of adult glioma patients
- Longitudinal analysis of factors related to Helicobacter pylori infection in Chinese adults
- HOXA10 enhances cell proliferation and suppresses apoptosis in esophageal cancer via activating p38/ERK signaling pathway
- Meta-analysis of early-life antibiotic use and allergic rhinitis
- Marital status and its correlation with age, race, and gender in prognosis of tonsil squamous cell carcinomas
- HPV16 E6E7 up-regulates KIF2A expression by activating JNK/c-Jun signal, is beneficial to migration and invasion of cervical cancer cells
- Amino acid profiles in the tissue and serum of patients with liver cancer
- Pain in critically ill COVID-19 patients: An Italian retrospective study
- Immunohistochemical distribution of Bcl-2 and p53 apoptotic markers in acetamiprid-induced nephrotoxicity
- Estradiol pretreatment in GnRH antagonist protocol for IVF/ICSI treatment
- Long non-coding RNAs LINC00689 inhibits the apoptosis of human nucleus pulposus cells via miR-3127-5p/ATG7 axis-mediated autophagy
- The relationship between oxygen therapy, drug therapy, and COVID-19 mortality
- Monitoring hypertensive disorders in pregnancy to prevent preeclampsia in pregnant women of advanced maternal age: Trial mimicking with retrospective data
- SETD1A promotes the proliferation and glycolysis of nasopharyngeal carcinoma cells by activating the PI3K/Akt pathway
- The role of Shunaoxin pills in the treatment of chronic cerebral hypoperfusion and its main pharmacodynamic components
- TET3 governs malignant behaviors and unfavorable prognosis of esophageal squamous cell carcinoma by activating the PI3K/AKT/GSK3β/β-catenin pathway
- Associations between morphokinetic parameters of temporary-arrest embryos and the clinical prognosis in FET cycles
- Long noncoding RNA WT1-AS regulates trophoblast proliferation, migration, and invasion via the microRNA-186-5p/CADM2 axis
- The incidence of bronchiectasis in chronic obstructive pulmonary disease
- Integrated bioinformatics analysis shows integrin alpha 3 is a prognostic biomarker for pancreatic cancer
- Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway
- Comparison of hospitalized patients with severe pneumonia caused by COVID-19 and influenza A (H7N9 and H1N1): A retrospective study from a designated hospital
- lncRNA ZFAS1 promotes intervertebral disc degeneration by upregulating AAK1
- Pathological characteristics of liver injury induced by N,N-dimethylformamide: From humans to animal models
- lncRNA ELFN1-AS1 enhances the progression of colon cancer by targeting miR-4270 to upregulate AURKB
- DARS-AS1 modulates cell proliferation and migration of gastric cancer cells by regulating miR-330-3p/NAT10 axis
- Dezocine inhibits cell proliferation, migration, and invasion by targeting CRABP2 in ovarian cancer
- MGST1 alleviates the oxidative stress of trophoblast cells induced by hypoxia/reoxygenation and promotes cell proliferation, migration, and invasion by activating the PI3K/AKT/mTOR pathway
- Bifidobacterium lactis Probio-M8 ameliorated the symptoms of type 2 diabetes mellitus mice by changing ileum FXR-CYP7A1
- circRNA DENND1B inhibits tumorigenicity of clear cell renal cell carcinoma via miR-122-5p/TIMP2 axis
- EphA3 targeted by miR-3666 contributes to melanoma malignancy via activating ERK1/2 and p38 MAPK pathways
- Pacemakers and methylprednisolone pulse therapy in immune-related myocarditis concomitant with complete heart block
- miRNA-130a-3p targets sphingosine-1-phosphate receptor 1 to activate the microglial and astrocytes and to promote neural injury under the high glucose condition
- Review Articles
- Current management of cancer pain in Italy: Expert opinion paper
- Hearing loss and brain disorders: A review of multiple pathologies
- The rationale for using low-molecular weight heparin in the therapy of symptomatic COVID-19 patients
- Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2
- Interleukin-35 in autoimmune dermatoses: Current concepts
- Recent discoveries in microbiota dysbiosis, cholangiocytic factors, and models for studying the pathogenesis of primary sclerosing cholangitis
- Advantages of ketamine in pediatric anesthesia
- Congenital adrenal hyperplasia. Role of dentist in early diagnosis
- Migraine management: Non-pharmacological points for patients and health care professionals
- Atherogenic index of plasma and coronary artery disease: A systematic review
- Physiological and modulatory role of thioredoxins in the cellular function
- Case Reports
- Intrauterine Bakri balloon tamponade plus cervical cerclage for the prevention and treatment of postpartum haemorrhage in late pregnancy complicated with acute aortic dissection: Case series
- A case of successful pembrolizumab monotherapy in a patient with advanced lung adenocarcinoma: Use of multiple biomarkers in combination for clinical practice
- Unusual neurological manifestations of bilateral medial medullary infarction: A case report
- Atypical symptoms of malignant hyperthermia: A rare causative mutation in the RYR1 gene
- A case report of dermatomyositis with the missed diagnosis of non-small cell lung cancer and concurrence of pulmonary tuberculosis
- A rare case of endometrial polyp complicated with uterine inversion: A case report and clinical management
- Spontaneous rupturing of splenic artery aneurysm: Another reason for fatal syncope and shock (Case report and literature review)
- Fungal infection mimicking COVID-19 infection – A case report
- Concurrent aspergillosis and cystic pulmonary metastases in a patient with tongue squamous cell carcinoma
- Paraganglioma-induced inverted takotsubo-like cardiomyopathy leading to cardiogenic shock successfully treated with extracorporeal membrane oxygenation
- Lineage switch from lymphoma to myeloid neoplasms: First case series from a single institution
- Trismus during tracheal extubation as a complication of general anaesthesia – A case report
- Simultaneous treatment of a pubovesical fistula and lymph node metastasis secondary to multimodal treatment for prostate cancer: Case report and review of the literature
- Two case reports of skin vasculitis following the COVID-19 immunization
- Ureteroiliac fistula after oncological surgery: Case report and review of the literature
- Synchronous triple primary malignant tumours in the bladder, prostate, and lung harbouring TP53 and MEK1 mutations accompanied with severe cardiovascular diseases: A case report
- Huge mucinous cystic neoplasms with adhesion to the left colon: A case report and literature review
- Commentary
- Commentary on “Clinicopathological features of programmed cell death-ligand 1 expression in patients with oral squamous cell carcinoma”
- Rapid Communication
- COVID-19 fear, post-traumatic stress, growth, and the role of resilience
- Erratum
- Erratum to “Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway”
- Erratum to “Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study”
- Erratum to “lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2”
- Retraction
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Retraction to “miR-519d downregulates LEP expression to inhibit preeclampsia development”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part II
- Usefulness of close surveillance for rectal cancer patients after neoadjuvant chemoradiotherapy