Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
-
Samuel K. Oppong
 und
Gabriel Kwabena Buabeng
und
Gabriel Kwabena Buabeng

Abstract
Aquatic macro-invertebrates play a vital role in the food chain of river ecosystem at several trophic guilds and consumer levels, and are used as biomonitoring tools for aquatic ecosystem health. However, hydrologic conditions of these ecosystems have been severely altered because of the increase in urban development and agricultural expansion. This study examined benthic invertebrate response to processes that structure their community in the Wewe River, segmented into intact, medium, and severe condition zones. We sampled in 100 stations in a period of 4 months in the wet (June–September, 2019) and 3 months in the dry (January–March, 2020) seasons. Geometric series, rarefaction, and Hill numbers models were used to quantify invertebrate assemblages, while ordination technique, canonical correspondence analysis, was used to evaluate the influence of predictive factors on their assemblages. A total of 2,075 individuals belonging to 20 family taxa were registered. There was no significant difference in benthic assemblages between the dry and wet seasons. Predictive factors accounted for 47.04 and 50.84% variances, respectively. Taxa distribution patterns differed significantly only in the severely disturbed zone during the wet season. Neptidae, Libellulidae, and Chironomidae were the most abundant taxa, indicating their broad range habitat preference and their ability to adapt to seasonal changes. Asellidae and Perlidae were the least detected, suggesting their sensitivity to elevated levels of some water quality parameters. The findings highlight the threats to the benthic community and overall functional state of the Wewe River, with the need to consider the proposed conservation interventions indicated in this study.
1 Introduction
Aquatic macro-invertebrates play an important role in the food chain of an ecosystem at several trophic guilds and consumer levels, and thus reflect ecosystem health [1]. Within the aquatic environment, macro-invertebrates range across a diverse range of microhabitats, with their diversity increasing in areas that provide abundant and diverse resources [2]. Their diversity and abundance are significant community attributes that are controlled by a variety of mechanisms at different spatial scales [3]. These environmental variables, which tend to influence their distribution and abundance, have been documented by a number of studies [3,4]. A good association among macro-invertebrate assemblages [3], which include chemical variables [5,6], organic energy base [7], and habitat-related physical factors such as substrate composition [8], elevation and stream size [9,10], vegetation, geology, and human land use [11,12], and temperature [10,13,14], has been documented as the factors influencing macro-invertebrate community assemblages. Other studies point to hydrologic conditions as the key driving forces affecting distribution and abundance patterns of benthic invertebrates [15,16,17]. For instance, studies on the hydro-climatic trends and variability over the Black Volta in Ghana suggest an increase in warming trends [18], and this phenomenon equally has the potential to impact on benthic invertebrate assemblages.
Changes in benthic invertebrate distributions caused by river regulation may occur because of altered habitat, flow patterns, sediment input, water quality, and thermal regimes [2,6]. Apart from these aforementioned drivers, yearly variations in seasons can be a factor that significantly affect the hydrologic regime and geomorphology in stream environments, by determining the distribution and abundance [12]. For example, a wet season low and a dry season high are expected for periodic seasonal patterns in abundance, depending on the frequency and intensity of summer monsoon rainfall [3].
Streams and rivers in urban centres worldwide have been severely polluted because of increase in urban development [19,20]. In Ghana, rivers draining through urban centres have underwent a significant transformation because of agricultural expansion and infrastructural development [21,22,23,24]. These threats could potentially impact on benthic invertebrate habitat quality, with a probable effect on their abundance, diversity, and spatial distribution. Scientific studies on macrobenthic invertebrates among urban rivers of Ghana remain poor, compared with extensive studies in similar areas in North Africa [25,26,27] and South Africa [28,29]. The few studies in Ghana’s freshwater systems have focused on hydroclimatic trends [18] and safe use of ground water [30]. The Wewe River is one of the few urban systems that drains through patches of the urban forest reserve in the Kumasi Metropolitan Area of Ghana. However, there have been concerns in recent times on the increasing level of human-led disturbances, namely farming activities, sewage disposal, tree felling, and bushfire [24].
Given the lack of scientific information about benthic invertebrates’ status in the Kumasi Metropolitan Area, it is not clear how these disturbance-related drivers have directly influenced physicochemical parameters and the consequent effect on benthic invertebrates. Furthermore, because of their sensitivity to aquatic environments, benthic invertebrates are widely considered as good indicators of water quality [14], by aiding in the identification of anthropogenic disturbances [1]. Thus, understanding how current water physicochemical parameters impact on benthic invertebrate assemblages is critical in choosing the appropriate conservation measures that will help restore the ecological integrity of Wewe River health. Secondly, the study on diversity and distribution patterns of benthic invertebrates and how they are influenced by physicochemical drivers are vital, because these organisms are used to track changes in the biological integrity of ecosystems [2].
In this paper, we assessed seasonal response of benthic invertebrate to physicochemical parameters in the Wewe River. To achieve this broad aim, we sought to answer the following objectives: (1) Are there variations in the abundance and distribution patterns along the three condition zones of the Wewe River? (2) Are there differences in benthic diversity among the three condition zones of the Wewe River? (3) What processes structured benthic assemblages among the three condition zones of the Wewe River? We hypothesized that (a) benthic invertebrate assemblages will differ between seasons, in terms of both taxonomic diversity and number of individuals, because benthic invertebrates have the capacity to recover rapidly from extreme drought periods [31]; (b) physicochemical drivers like dissolved oxygen (DO), total dissolved solids (TDS), electrical conductivity (EC), surface water temperature, salinity, mercury content, water depth, stream flow (slow, medium, and fast), and substrate composition that structure benthic invertebrate communities will vary in the wet and dry seasons, because seasonality tends to directly influence physicochemical drivers.
2 Materials and methods
2.1 Study area
The study area is a suburban forest reserve in the Kumasi Metropolis of Ghana and surrounded by communities like Ayigya, Weweso, Bomso, Gyenyasi, Kentikrono, and Ahinsan. Farming along the fringes of the river occurs year round. Waste water from nearby settlements and farm waste equally drains into the river course through direct channel connectivity. Wewe River is a typical unregulated system and takes its source from mountains near Aboabo Nkwanta and flows for about 8.125 km southwest towards Abirem and Weweso [32]. The river is located between N 06°41′30.1″, W 001°33′74.4″ and N 06°40′32.9″, W 001°34′20.9″ (Figures 1 and 2). Soil is typically heavy clay to loamy, characterized by cobbles and boulders. The rock type is igneous and metamorphic rocks, with undulating topography. The average temperature is 24–34°C p.a. and generally humid. Rainfall pattern is typically bimodal, with annual average of 2,000 mm p.a. [33]. The study was conducted in a period of 4 months in the wet (June–September, 2019) and 3 months in the dry (January–March, 2020) seasons, spanning a total of 7 months.
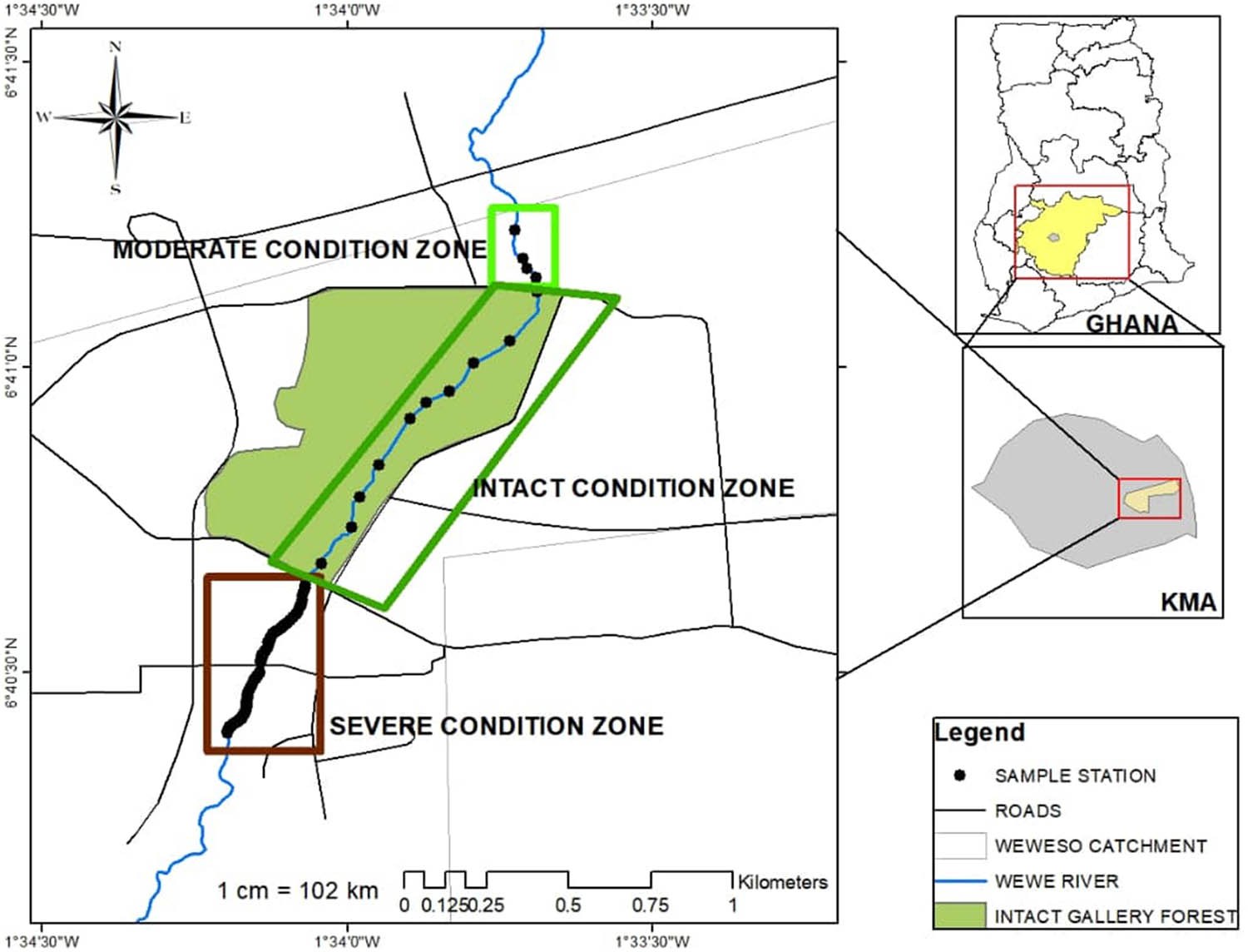
Map of Ghana, showing the study area in the Kumasi Metropolitan Area (Ashanti Region).

Photographs of some segments of the unregulated Wewe River, where the study was conducted during the wet (June–September, 2019) and dry (January–March, 2020) seasons. a1 & a2 represent the severely disturbed zone (downstream of the river), b = intact zone (river midstream) and c = moderately disturbed zone (river upstream).
2.2 Classification of the sampling sites on the riparian zone
We initially demarcated the upstream, midstream, and downstream of the Wewe River into three condition zones (i.e. intact, moderate, and severe condition zones, following the Riparian Quality Index (RQI) methods). The river course was classified into three condition zones, because it is surrounded by a forest reserve, with some disturbance along the riparian zone, which could have a direct influence on the water quality and consequently on benthic community structure. The index ranges from 1 and 15. Thus, intact condition class (10–15): areas dominated by different vegetation strata that cover the full length of the segment, which is linked to natural fluvial forms and slightly fragmented; moderately disturbed condition class (7–9): areas with vegetation cover nearly half of the study zone being disturbed; 1–3 m active channel width and about 10–30% exotic and ruderal species present; severely disturbed condition class (1–6): areas where 60% of the riparian corridor is reduced by human-led activities; vegetation covering <30% (mainly grasses/herbs and isolated woody species), with channel banks connected to agricultural fields. The end-to-end distance within each condition zone included the following: 180 m (upstream, moderately disturbed condition zone), 937 m (midstream, intact condition zone), and 533 m (downstream, severely disturbed condition zone, with an accuracy of ±5 m) (Figure 1, Plate 1). The RQI methods represent a useful tool for monitoring and evaluating the structure of riparian zones, an element of the river morphological conditions.
2.3 Benthic macro-invertebrate sampling procedure
Benthic invertebrates were collected in biweekly intervals for a 7-month period – 4 months in the wet season (June–September, 2019) and 3 months in the dry season (January–March, 2020). There is no rainfall in the dry season, and this reflects in the low flow rate. A total of 100 samples were randomly collected across the three segments (i.e. upstream [moderate condition zone] = 30 samples, midstream [intact condition zone] = 40 samples, and downstream [severe condition zone] = 40 samples), with six replicates per sample point. We sampled each condition zone once per week. Invertebrates were collected using a D-frame sweep net (800 μm mesh size, 690 cm2 mouth area, and 1 m length dimensions). The D-frame sweep net was driven deep into the sediment and against the flow direction, to trap all organisms inhabiting the sediment column beneath surface water. The D-frame sweep nets are best used as qualitative or semi-quantitative analyses where a diversity of specimens (e.g. IBI or other indices) is more important than density estimates sampling.
Sediments were collected during low tide to avoid pressure waves displacing the finest surface sediments, and subsequently placed in a petri dish filled with soda water and labelled according to the segment they were sampled. They were subsequently transported to the laboratory, where all organisms were sorted from detritus and inorganic materials by sieving on a mesh hardware cloth basket and stored in 95% ethanol [34]. Organisms were then identified up to the family level with the aid of a stereomicroscope (LEICA MZ6) [35] and taxonomic keys provided by Guide to Freshwater invertebrates [36] and Asian stream guide for identification [37]. Voucher specimens that we could not identify were sent to the Faculty of Biological Sciences for identification.
2.4 Measurement of physicochemical parameters
Water quality variables, DO, TDS, EC, surface water temperature, salinity, mercury content (mm Hg), and pH, were measured in situ with a multi-probe portable meter (Hanna instrument model H19828). Physicochemical drivers were log transformed where appropriate to achieve normality.
2.5 Statistical analysis
Objective 1: Are there variations in the abundance and distribution patterns of benthic taxa along the three condition zones of the Wewe River?
2.5.1 Benthic invertebrate abundance distribution along the three condition zones
Both benthic invertebrates and physicochemical drivers’ dataset were initially subjected to square root transformation to homogenize variances and achieve normality [38]. Invertebrate abundance as a measure of diversity was quantified by applying the rank abundance distribution model [39,40]. Geometric series (GS) model was then fitted to the benthic invertebrate data using regression model approach [41], to determine their distribution patterns along the condition zones in the riverine continuum. This model approach was used to test against the null hypothesis (H o) that invertebrate abundance distribution and richness did not differ in each of the three segments or condition zones classes of the Wewe River.
All registered invertebrate order in each of the three segments of the river were ranked from the most to the least abundant on the rank abundant curve [42], with each species rank plotted on the x-axis and the abundance plotted on the y-axis. Analysis of covariance was used to test for the significant difference of the equality of the slope of invertebrate abundance distributions among the three segments of the Wewe River.
Objective 2: Are there differences in benthic richness and diversity among the three condition zones of the Wewe River?
2.5.2 Analyses of benthic richness and diversity
Individual-based rarefaction model [43] was performed to determine invertebrate richness. The rarefaction curve
where
Finally, invertebrate diversity was quantified using Hill numbers [48,49,50,51,52]. We used Hill numbers because they incorporate relative abundance and species richness in diversity analysis [49] and defined as follows:
where S is the number of species in the assemblage, and the i th species has relative abundance p i , i = 1, 2,..., S.
Objective 3: What processes structured benthic assemblages among the three condition zones of the Wewe River?
2.5.3 Analysis of benthic taxa-physicochemical driver relationship
Canonical correspondence analysis (CCA) was used to evaluate the influence of predictive factors on benthic assemblages [53]. CCA is a direct ordination method, with the resulting product being the variability of the physicochemical drivers and benthic invertebrate data [54]. A ridge regression was performed to remove multicollinearity (i.e. perfect correlation with other predictive factors, which tend to inflate variances of the parameter estimates) [55,56]. Mixed ANOVA test (a parametric technique) or split-plot ANOVA was used to test for a significant difference in invertebrate assemblages (abundance, richness, and diversity) across the three condition zones and the physicochemical drivers. Student’s t-test was performed to determine seasonal variability among invertebrates’ families and physicochemical drivers. Where significant difference was detected, we further used Tukey’s HSD post hoc test to determine the habitats that differed. Spearman’s rank correlation test was performed to evaluate the significant relationship among physicochemical drivers. All the analyses were performed using PAST ver. 3.18 Package [57].
3 Results
3.1 Seasonal trends in benthic invertebrate composition and individual abundance across the three condition zones
A total of 2,075 individuals belonging to 20 benthic invertebrate families were registered in the dry (n = 693) and wet (n = 1,382) seasons and among the habitat condition zones (Figure 3). Variations among mean individuals were substantial in the dry (F 2,33 = 63.56, P < 0.0007) and wet (F 2,48 = 73.86, P < 0.0001) seasons (Figure 4a and b). Considerable variations among mean number of individuals in the dry season were observed between the moderate and severe (P < 0.0003, Tukey’s post hoc test) and intact and severe condition zones (P < 0.0001), whereas during the wet season, it was between the moderate and severe condition zones (P < 0.0007) and intact and severe condition zones (P < 0.0006). Although we detected more family taxa in the wet season (n = 17) than the dry season (n = 12), mean seasonal variations were not substantially significant (Student’s t-test = −0.329, P = 0.75) (Figure 5). From individual condition zones, the upstream of the river which constitutes the moderate condition zone recorded the highest mean number of individuals per family taxa in the dry (3.25 ± S.E. 0.85) and wet seasons (2.25 ± S.E. 1.3), while the lowest number was registered in the severe condition zone (downstream) during the dry season (0.25 ± S.E. 0.05) and intact condition zone in the wet season (0.22 ± S.E. 0.22) (Figure 4a and b).
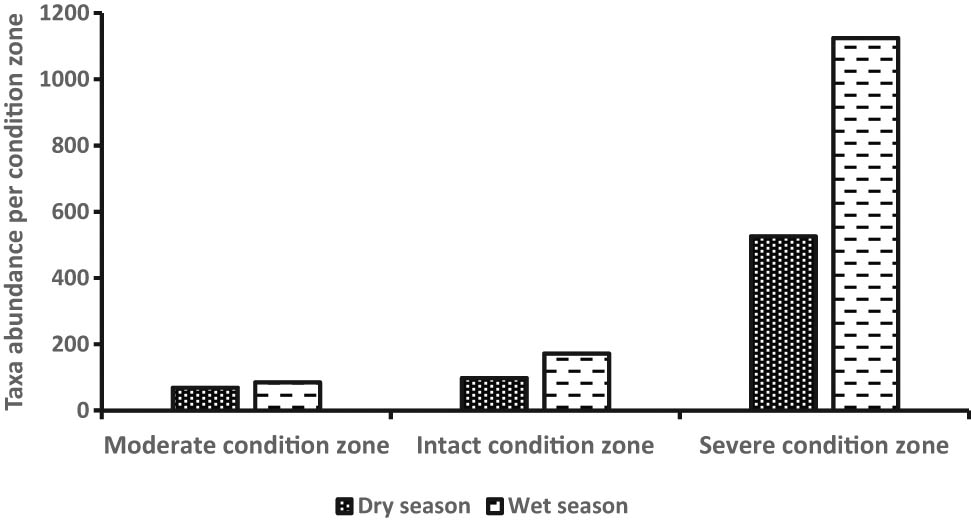
Comparison of seasonal abundance among the three conditions in Wewe River.
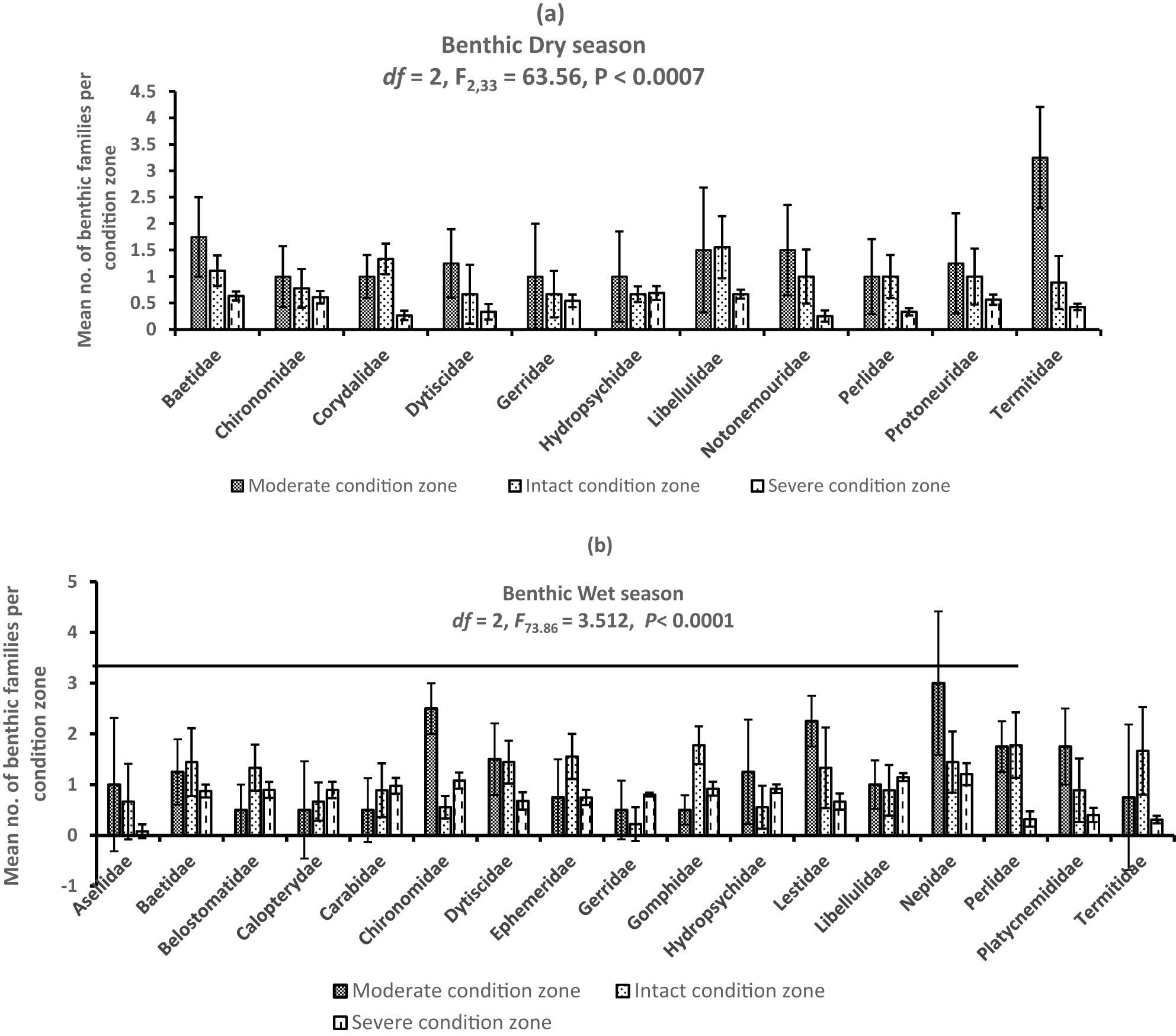
(a and b) Variations in mean composition of benthic families in the Wewe River in the dry and wet seasons. Notice that Chironomidae was the most dominant family across the three habitat condition zones of the river in the wet season.
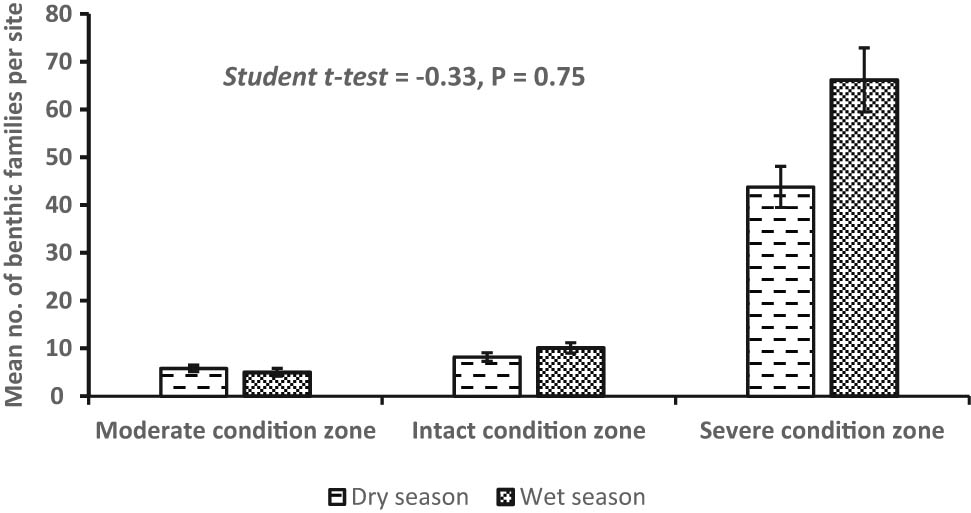
Variations in the seasonal composition of benthic invertebrate along the riverine continuum. Notice that benthic invertebrates were generally higher in the wet season than the dry season. Means are not significantly different.
3.2 Benthic family distribution patterns in the three condition zones
We found in all cases that invertebrate family abundance distribution among the three habitat condition zones fitted well in the GS model (Figure 6). However, comparison between the dry (C.V. = 52.38%) and wet (C.V. = 35.84%) seasons revealed that invertebrates were not significantly distributed (Z = −0.517, P = 0.61, Fligner Kileen test for equal C.V.) (Figure 5). Individual segments of the Wewe River showed that taxa abundance distribution along the curves was not significantly different in the moderate (χ²P = 2.69, P < 0.98), intact (χ²P = 2.49, P < 0.99), and severely disturbed (χ²P = 5.15, P < 0.88) condition zones in the dry season (Figure 6, Table 1). Similarly, in the wet season, there was no substantial difference in the moderate (P < 0.901) and intact (P < 0.99), with the exception of the severely disturbed condition zone (χ²P = 75.81, P < 0.0004) (Figure 6, Table 1).
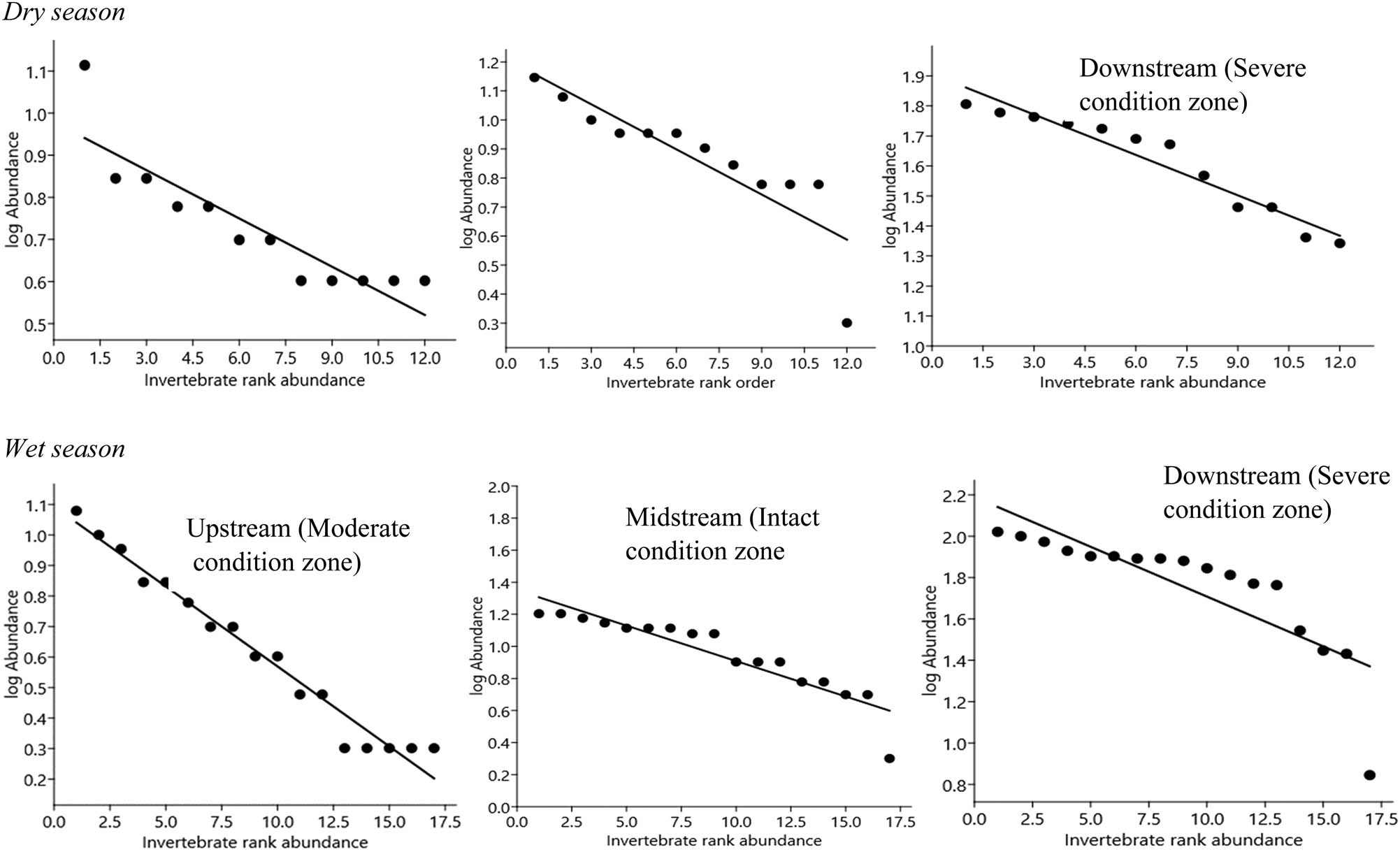
GS model for benthic families rank abundance distribution across the three condition zones in the dry and wet seasons, on the Wewe River. Abundance is based on cumulative count values per sample zone. Notice that benthic families are ordered in decreasing magnitude and plotted against their corresponding rank.
χ 2 p goodness of fit for the GS model for the abundance rank distribution of benthic invertebrate, calculated for all three segments of the Wewe River in the dry and wet seasons. The variable α determines the shape of the distribution
| River segments | α | χ²P-value for GS | P-level |
|---|---|---|---|
| Dry season | |||
| Moderate | 0.084 | 2.69 | 0.98 |
| Intact | 0.112 | 2.49 | 0.99 |
| Severe | 0.098 | 5.15 | 0.88 |
| Wet season | |||
| Moderate | 0.114 | 0.43 | 0.90 |
| Intact | 0.096 | 4.76 | 0.99 |
| Severe | 0.105 | 75.81 | 0.0004*** |
Significance of P-values: *P < 0.05, **P < 0.01, ***P < 0.0001.
Of the 20 benthic families registered, Libellulidae (n = 78), Odontopygidae (n = 73), and Baetidae (n = 72) were the highest ranked on the abundance distribution curve and were widely distributed across the three condition zones, during the dry season (Figure 6, Table 2). Rarer orders such as Notonemouridae (n = 37, 5.3%), Dytiscidae (n = 40, 5.7%), and Perlidae (n = 42, 6.0%) were the least ranked in the moderate and intact condition zone, reflecting their sensitivity to habitat perturbation. Overall, the severely disturbed condition zone supported the most abundant family taxa.
Summary of individual benthic invertebrates recorded in the dry and wet seasons, along the three condition zones, in the Wewe River. N = 100 sample plots
| Macro-invertebrate families | Total number of individuals | Total | ||
|---|---|---|---|---|
| Moderate condition zone | Intact condition zone | Severe condition zone | ||
| Dry season | ||||
| Baetidae | 7 | 10 | 55 | 72 |
| Chironomidae | 4 | 7 | 53 | 64 |
| Corydalidae | 4 | 12 | 23 | 39 |
| Dytiscidae | 5 | 6 | 29 | 40 |
| Gerridae | 4 | 6 | 47 | 57 |
| Hydropsychidae | 4 | 6 | 60 | 70 |
| Libellulidae | 6 | 14 | 58 | 78 |
| Notonemouridae | 6 | 9 | 22 | 37 |
| Odontopygidae | 7 | 2 | 64 | 73 |
| Perlidae | 4 | 9 | 29 | 42 |
| Protoneuridae | 5 | 9 | 49 | 63 |
| Termitidae | 13 | 8 | 37 | 58 |
| Total | 69 | 98 | 526 | 693 |
| Wet season | ||||
| Asellidae | 4 | 6 | 7 | 17 |
| Baetidae | 5 | 13 | 76 | 94 |
| Belostomatidae | 2 | 12 | 78 | 92 |
| Calopterydae | 2 | 6 | 78 | 86 |
| Carabidae | 2 | 8 | 85 | 95 |
| Chironomidae | 10 | 5 | 94 | 109 |
| Dytiscidae | 6 | 13 | 59 | 78 |
| Ephemeridae | 3 | 14 | 65 | 82 |
| Gerridae | 2 | 2 | 70 | 74 |
| Gomphidae | 2 | 16 | 80 | 98 |
| Hydropsychidae | 5 | 5 | 80 | 90 |
| Lestidae | 9 | 12 | 58 | 79 |
| Libellulidae | 4 | 8 | 100 | 112 |
| Neptidae | 12 | 13 | 105 | 130 |
| Perlidae | 7 | 16 | 28 | 51 |
| Platycnemididae | 7 | 8 | 35 | 50 |
| Termitidae | 3 | 15 | 27 | 45 |
| Total | 85 | 172 | 1,125 | 1,382 |
In the wet season, Neptidae (n = 130, 9.4%), Libellulidae (n = 112, 8.1%), and Chironomidae (n = 109, 7.9%) were the highest ranked invertebrate families in all three condition zones, while Asellidae (n = 17) and termitidae (n = 45) were the least ranked and infrequent taxa, whose distribution occurred in narrow ranges in the three condition zones (Figure 6). These benthic families constituted 1.2 and 3.2%, respectively, of the total number of invertebrates sampled.
Observation from individual condition zones revealed that the abundance and widespread distribution of benthic taxa occurred in the severely disturbed condition zone during the dry and wet seasons, thus reflecting their broad range tolerance to varying physicochemical concentrated levels. Comparison of invertebrate abundant distribution for the three condition zones distinguishes them in relation to the influence of predictive factors. Thus, the shape of the rank abundance curves generally showed differences in invertebrate relative dominance and spatial distribution from individual sample stations.
3.3 Benthic invertebrate richness and diversity along the Wewe Riverine system
Seasonal variations in taxa richness were not significant (Student’s t-test = 4.335, P < 0.62), although the number of benthic families appeared more in the wet season (n = 17) than in the dry season (n = 12) (Figure 7). Generally, the severe condition zone was the richest in family taxa, while the moderate condition zone was the poorest. Libellulidae was more dominant in the intact (midstream) and severe (downstream) condition zones during the dry season, while Neptidae was dominant in all three condition zones in the wet season (Figure 7). Observed variability in taxa richness and abundance distribution patterns across the three segments of the Wewe River reflected in their diversity profile and ranked from higher to lower diversity indices along the alpha (a) scale values (Figure 7). Condition zone with shallow curve is the most diverse and highest ranked, while those with steep curves are the least diverse and found at the bottom of the Hill number diversity profile.
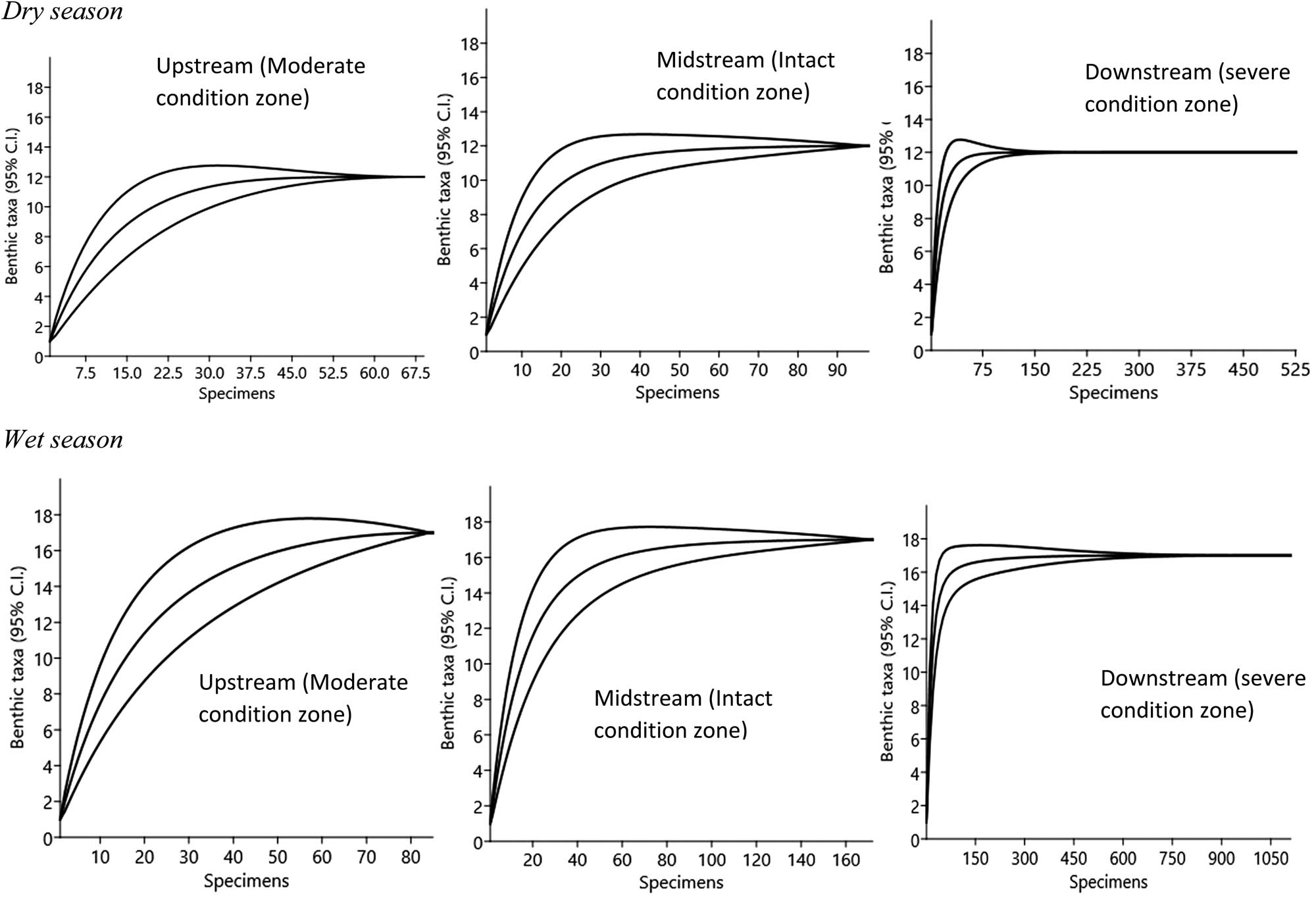
Standardized comparison of benthic invertebrate richness for individual-based rarefaction curves. The data represent summary counts of benthic invertebrates that were recorded from the three condition zones of the Wewe River. The rarefaction curves were calculated from equation (3) (Gotelli and Colwell, 2001), with a 95% confidence interval. The dotted vertical lines illustrate family richness comparison standardized to 69 (dry season) and 85 (wet season) individuals, which was the observed abundance in the upstream of the three segments benthic data set. The average of these individual curves represents the statistical expectation of the species accumulation curve for that particular sample drawn on re-orderings, and the variability among the different orderings is reflected in the specific variance (conditional) in the number of families recorded for any given number of individuals.
Diversity generally did not differ significantly between the dry (F 2,6 = 0.0461, P = 0.65) and wet (F 2,6 = 1.06, P = 0.40) seasons, and ranged as q D = 2.974–2.996 in the dry and wet seasons (Figure 8, Table 3). However, from individual condition zones, we found the intact condition zone (midstream of the river in red colour) ( q D = 2.996) and the severe condition zone (downstream of the river, in blue colour) ( q D = 2.991) to be the most diverse in the dry and wet seasons, respectively (Figure 8, Table 3). Invertebrate diversity appeared similar in the different condition zones, suggesting similar patterns in spatial evenness distribution. The moderate condition zone (upstream of the river) was consistently least diverse in the dry ( q D = 2.974) and wet ( q D = 2.854) seasons. The low diversity in the moderate condition zone appears to reflect in its lowest abundance of individuals in both seasons as well (Figure 8, Table 2).
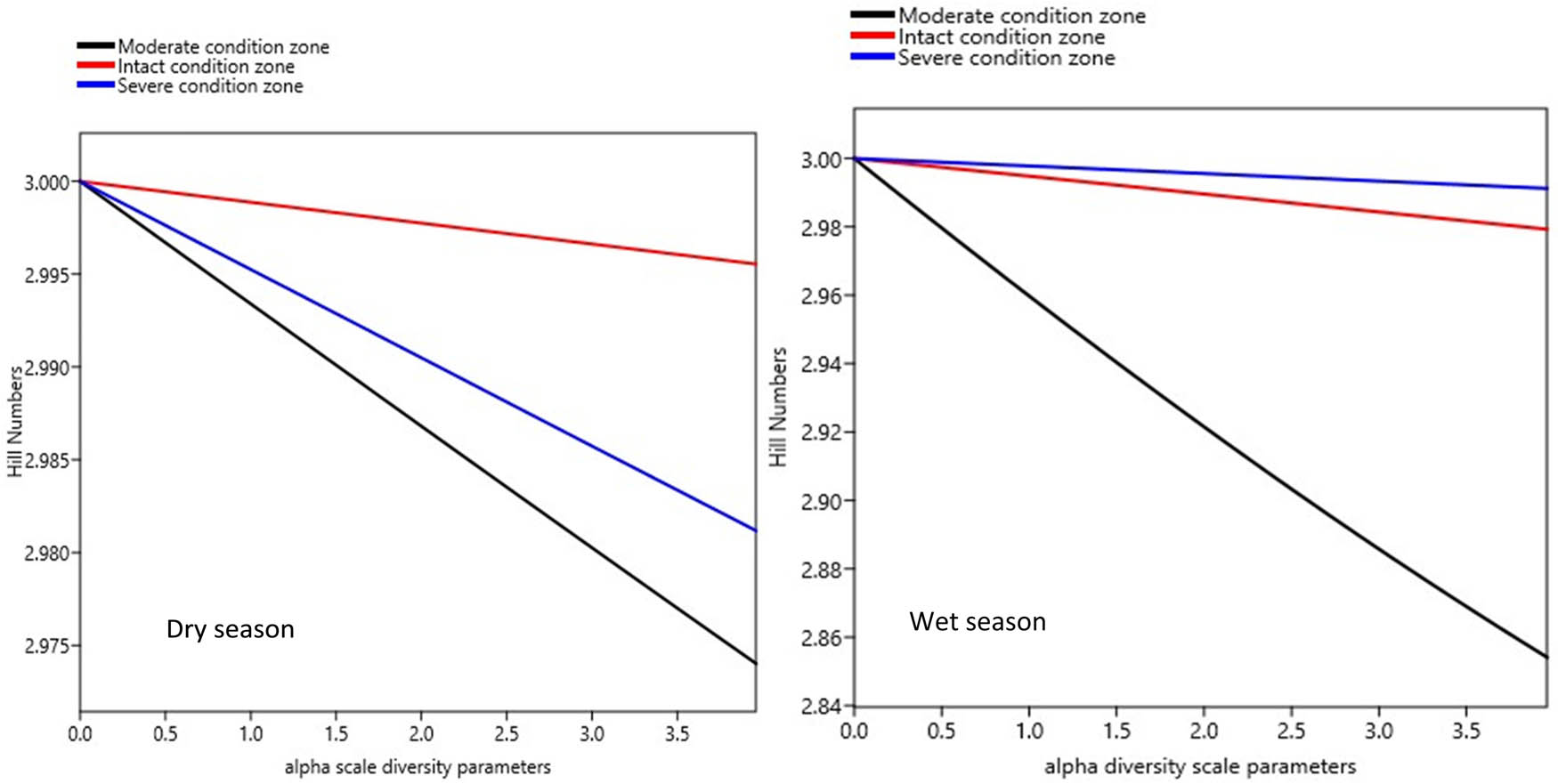
Seasonal trends in benthic invertebrate diversity among the three condition zones, in the Wewe River. Shallower curves reflect high diversity (top of the curves), while steeper curves indicate less diversity (bottom of the curves). Notice that the midstream (intact zone and top red curve) is the most diverse in the dry season, while the downstream (severe condition zone and top blue curve) is the most diverse during the wet season.
Summary of Hill’s number diversity order (at q = 0, 1, and 2), along the three condition zones, in the Wewe River. The order q is mathematically unified family of diversity indices differing among themselves only by an exponent q and indicates their likelihood to include or ignore the relatively rarer species (Hill, 1973). Thus, q = 0 represents the number of species in the sample (richness index), q = 1 the exponential of the Shannon–Weiner index, and q = 2 the reciprocal of the Simpson’s index (i.e. equivalent number of species)
| Alpha values | Moderate condition zone | Intact condition zone | Severe condition zone |
|---|---|---|---|
| Dry season | |||
| 0 (Number of sp.) | 12 | 12 | 12 |
| 1 (exp[Shannon]) | 10.082 | 10.627 | 10.169 |
| 2 (Inv.[Gini_Simpson]) | 9.352 | 10.286 | 9.591 |
| Shannon_Evenness | 0.916 | 0.966 | 0.924 |
| Hill diversity numbers | 2.974 | 2.2996 | 2.981 |
| Wet season | |||
| Alpha values | |||
| 0 (Number of sp.) | 17 | 17 | 17 |
| 1 (exp[Shannon]) | 11.608 | 12.771 | 13.249 |
| 2 (Inv.[Gini_Simpson) | 9.325 | 12.159 | 12.775 |
| Shannon_Evenness | 0.829 | 0.912 | 0.946 |
| Hill diversity numbers | 2.854 | 2.979 | 2.991 |
3.4 Seasonal environmental influence on benthic community structure across the three condition zones in the Wewe River
The matrices of the invertebrate-site biplot generated by CCA showed DO (r = 0.54, P < 0.01) and surface water temperature (r = 0.31, P < 0.05) on Axis I, and total dissolved solids (r = 0.32, P < 0.05) and EC (r = 0.32, P < 0.05) on Axis II, as the major physicochemical drivers of benthic invertebrate assemblages among the three condition zones in the dry season (Figure 9a and b; Tables 4 and 5). Whereas in the wet season, EC (r = −0.63, P < 0.01)), TDS (r = 0.43, P < 0.05), and SS (r = 0.31, P < 0.05) on Axis I and Mercury (r = 0.42, P < 0.05) and pH (r = 0.38, P < 0.05) on Axis II were found to influence the community structure of benthic invertebrate (Figure 10; Tables 4 and 5). The first two axes in the dry season (Axes I = 31.28%, II = 19.56%) accounted for 50.84%, while the wet season totaled 47.04% (Axes I = 38.63%, II = 15.41%) of variations in the weighted averages of the 14 invertebrate families in relation to six physicochemical drivers (Table 5). Physicochemical drivers did not differ among the three condition zones of the Wewe River in the dry (F 2,18 = 0.0026, P < 0.99) and wet (F 2,18 = 0.044, P < 0.95, one-way ANOVA test) seasons. Furthermore, seasonal variability in physicochemical drivers was equally not significant (Student’s t-test = −0.033, P < 0.97).
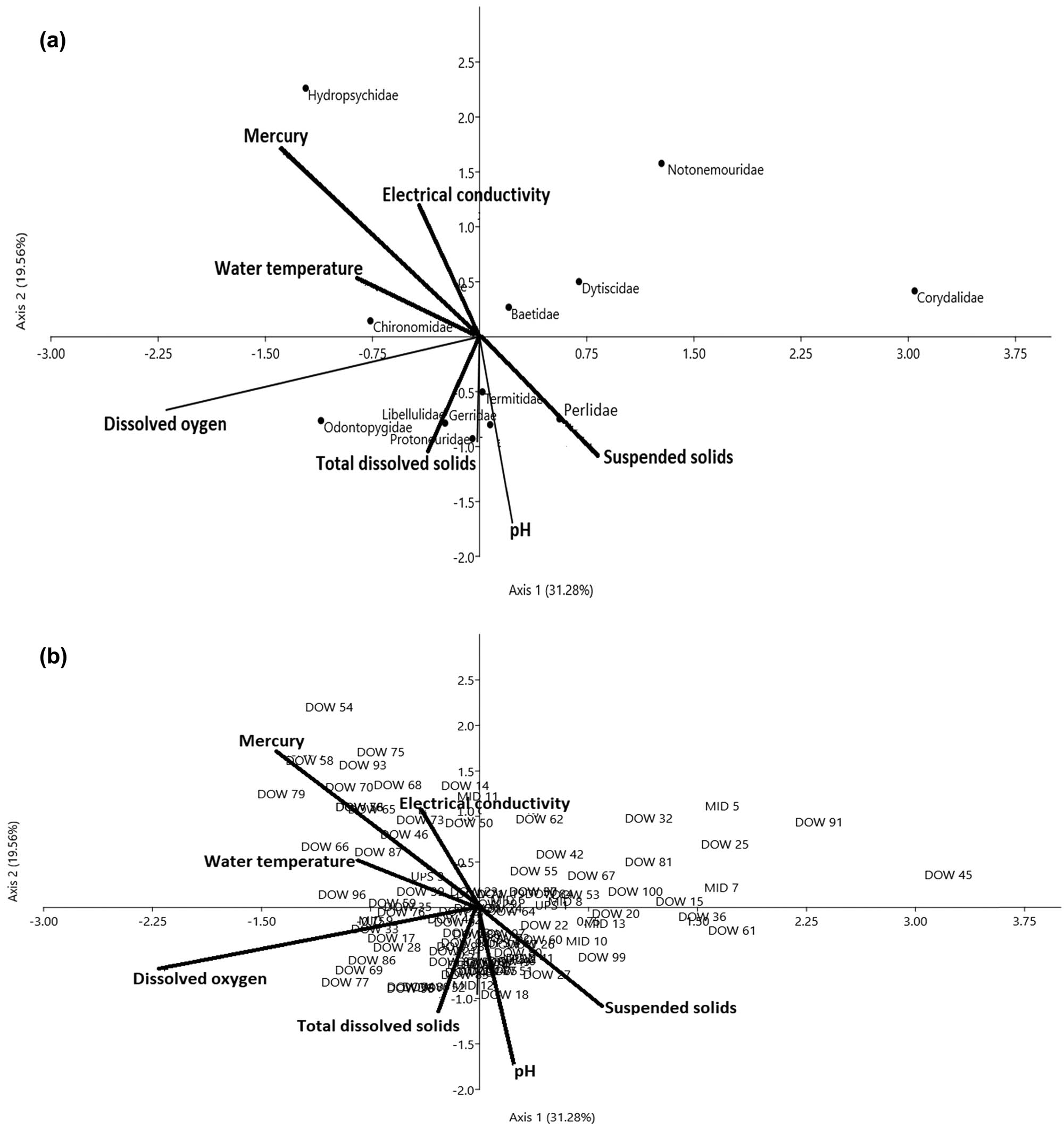
(a and b) CCA diagram showing the influence of physicochemical drivers on benthic invertebrate assemblages in the dry season. The first two axes (Axes I = 31.28 and II = 19.56) explained 50.9% of variations across the three condition zones in the dry season. The arrows represent each of the physicochemical drivers plotted pointing in the direction of maximum change of explanatory variables among the three habitats. Sample plot codes represent each of the three condition zones along the river course (i.e. DOW = downstream of the severe condition zone, MID = midstream of the intact condition zone, and UPS = upstream of the moderate condition zone).
Summary of water quality parameters of the 100 sample stations, across the three condition zones of the Wewe River, in the dry and wet seasons
| Moderate condition zone | Intact condition zone | Severe condition zone | |
|---|---|---|---|
| Dry season | |||
| Dissolved oxygen (mg/L) | 4.6 ± 0.3 | 3.0 ± 0.2 | 5.0 ± 0.1 |
| Total dissolved solids (ppm) | 88.8 ± 0.6 | 90 ± 0.5 | 99 ± 1.2 |
| Electrical conductivity (μS/cm) | 329.5 ± 3.8 | 397.6 ± 4.8 | 357.6 ± 1.9 |
| Water temperature (°C) | 26.4 ± 0.7 | 27.9 ± 0.3 | 28.1 ± 0.1 |
| Mercury (mm Hg) | 738.1 ± 0.0 | 738.3 ± 3.3 | 738.7 ± 0.1 |
| Suspended solids (mg/L) | 177.8 ± 1.3 | 179.7 ± 0.9 | 185.5 ± 1.04 |
| pH | 6.2 ± 0.6 | 6.0 ± 0.4 | 5.9 ± 0.1 |
| Wet season | |||
| Dissolved oxygen (mg/L) | 4.5 ± 0.3 | 4.9 ± 0.3 | 5.0 ± 0.1 |
| Total dissolved solids (ppm) | 71.3 ± 2.2 | 57.7 ± 6.4 | 84.5 ± 1.1 |
| Electrical conductivity (μS/cm) | 499.4 ± 17.9 | 483.4 ± 44.9 | 169 ± 2.2 |
| Water temperature (°C) | 25.9 ± 0.4 | 26.2 ± 0.2 | 26 ± 0.1 |
| Mercury (mm Hg) | 739.6 ± 0.1 | 738.8 ± 1.4 | 739.4 ± 0.1 |
| Suspended solids (mg/L) | 199.5 ± 25.6 | 205.8 ± 8.9 | 238.3 ± 3.5 |
| pH | 6.6 ± 0.3 | 6.2 ± 0.4 | 5.3 ± 0.1 |
Summary of CCA axis lengths for ground cover, showing the levels of correlation between axes and physicochemical driver gradients, percentage variance of benthic taxa and benthic taxa–physicochemical driver relationship
| Dry season | Wet season | |||
|---|---|---|---|---|
| Axis | I | II | I | II |
| Canonical eigenvalue | 0.184 | 0.041 | 0.109 | 0.035 |
| % variance explained | 31.28 | 19.56 | 38.63 | 15.41 |
| No. of variables = 6 | ||||
| Correlations | ||||
| DO | 0.538* | −0.053 | 0.033 | −0.013 |
| TDS | 0.071 | −0.317* | 0.425* | −0.173 |
| EC | −0.026 | 0.324* | −0.631** | 0.031 |
| Temp. | 0.314* | 0.145 | 0.026 | 0.176 |
| mm Hg | 0.013 | −0.004 | 0.133 | −0.417* |
| SS | 0.082 | −0.109 | 0.313* | −0.003 |
| pH | 0.162 | 0.076 | −0.075 | 0.377* |
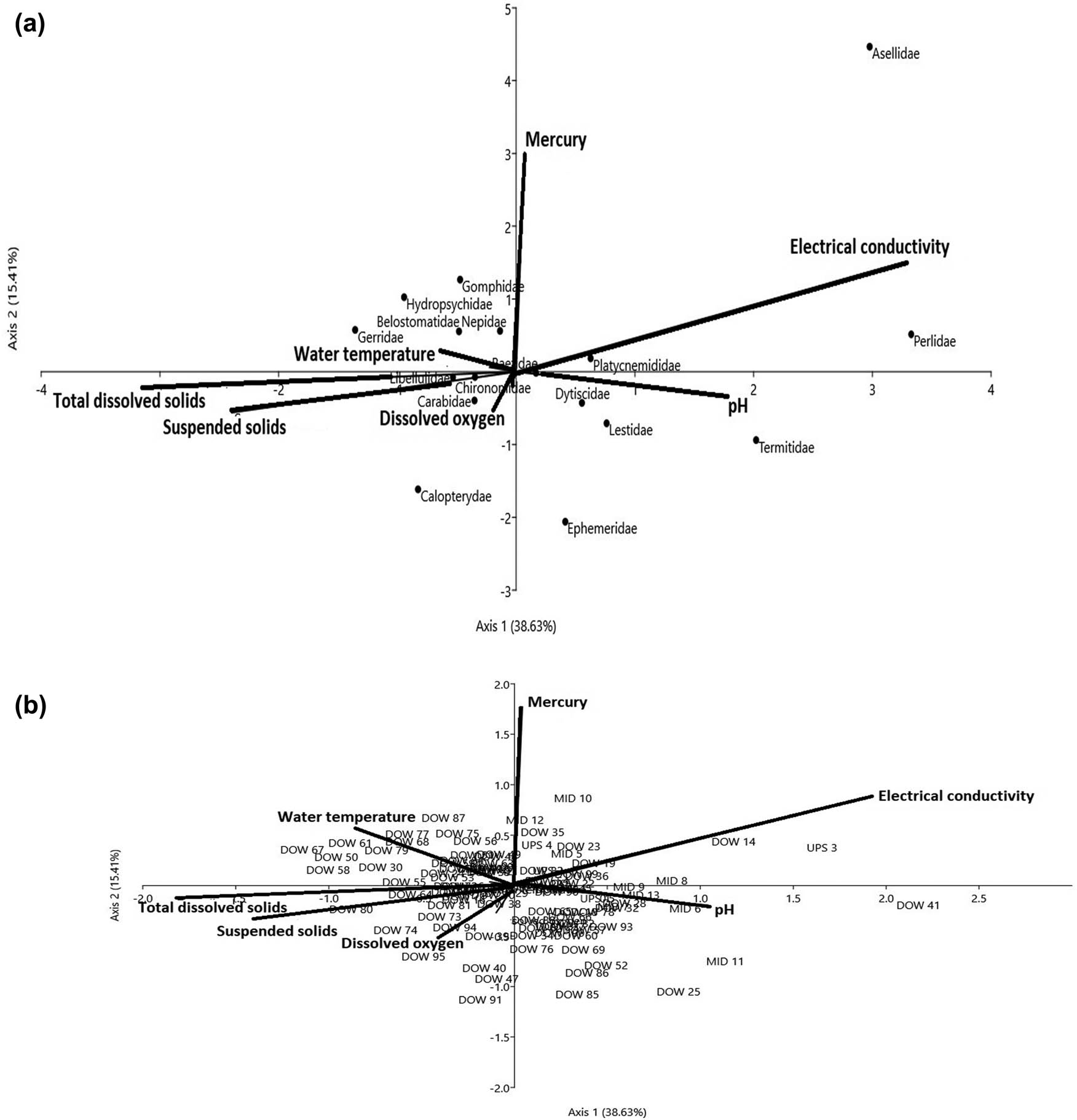
(a and b) CCA diagram showing the influence of physicochemical drivers on benthic invertebrate assemblages in the wet season. The first two axes (Axes I = 31.63 and II = 15.41) explained 47.04% of variations across the three condition zones in the dry season. The arrows represent each of the physicochemical drivers plotted pointing in the direction of maximum change of explanatory variables among the three habitats. Sample plot codes represent each of the three condition zones along the river course (i.e. DOW = downstream of the severe condition zone, MID = midstream of the intact condition zone, and UPS = upstream of the moderate condition zone).
In the dry season, Hydropsychidae and Chironomidae correlated positively with optimal levels of surface water temperature (r = 0.31, P < 0.05) and EC (r = 0.32, P < 0.05) in the severe condition zone (DOW = downstream), along Axes I and II (upper left of the CCA diagram). On the lower left of the ordination diagram, Libellulidae, Odontopygidae, and Gerridae were negatively associated with total dissolved solids (r = −0.32, P < 0.05) and DO (r = 0.54, P < 0.05) on Axes I and II (Figure 9a and b, Table 5). These sections of the river bed were characterized by cobble-gravel, boulders, and a high flow rate. Intercorrelation between DO and suspended solids (SS) (r s = 0.76, P < 0.01) may be an effect of the high flow rate, causing oxygen to dissolve in the water, and simultaneously carrying floating debris (Figure 9a and b, Table 6). On the lower right of the CCA diagram, we found that weak acidic pH levels appeared to affect the abundance of Perlidae (n = 4, 5.7%) and Termitidae (n = 8, 11.6%) in the moderate (MID – midstream) and severely disturbed (DOW – downstream) condition zones. The least abundance of Notonemouridae (n = 9) and Corydalidae (n = 9) in these two zones was largely linked to the influence of EC on Axis II (Figure 9a and b). Taxa found at the middle of the CCA diagram (i.e. Baetidae and Dytiscidae) appeared to exhibit broad tolerance to average levels of all physicochemical drivers assessed in the three condition zones of the Wewe River.
Summary of Spearman rank (r s) correlation matrix between the physicochemical drivers across the three habitats on the riparian zone. A correlation above/below ±0.61 is significant at *** P = 0.001; ±0.45 at ** P = 0.01, and ±0.33 at * P = 0.05
| Total dissolved solids | Electrical conductivity | Temperature | Mercury | Suspended solids | pH | |
|---|---|---|---|---|---|---|
| Dry season | ||||||
| Dissolved oxygen | 0.04 | 0.135 | 0.232 | 0.723*** | 0.765*** | 0.854*** |
| Total dissolved solids | 0.011 | 0.595** | 0.585** | 0.051 | 0.486** | |
| Electrical conductivity | 0.061 | 0.034 | 0.746*** | 0.679** | ||
| Temperature | 0.026 | 0.094 | 0.146 | |||
| Mercury | 0.012 | 0.006 | ||||
| Suspended solids | 0.738*** | |||||
| pH | ||||||
| Wet season | ||||||
| Dissolved oxygen | 0.0003 | 0.031 | 0.033 | 0.848*** | 0.001 | 0.090 |
| Total dissolved solids | 0.0004 | 0.005 | 0.056 | 0.014 | 0.344* | |
| Electrical conductivity | 0.003 | 0.087 | 0.255 | 0.038 | ||
| Temperature | 0.706*** | 0.057 | 0.073 | |||
| Mercury | 0.430* | 0.735*** | ||||
| Suspended solids | 0.660** | |||||
| pH | ||||||
In the wet season, we observed that increased level of mercury (r = −0.42, P < 0.05) and EC (r = −0.63, P < 0.01) on Axes I and II contributed to the low abundance of Asellidae, Perlidae, and Platycnemididae, located in the upper right hand of the ordination diagram. Similarly, the influence of weak to near-neutral pH levels (r = −0.38, P < 0.05) partly contributed to high abundance of Dytiscidae, Ephemeridae, and Lestidae. However, abundance of Termitidae (n = 3) was affected by the near-neutral levels of pH, especially in the moderately disturbed MID – midstream, condition zones (Figure 10a and b, Table 5). The use of organophosphate pesticides to control pest invasion on the nearby vegetable farms and liquid waste discharge from human settlement may have contributed to a decrease in DO concentration (lower left of CCA diagram), and this impacted on Carabidae dominance. This was evidenced in the strong intercorrelation between mercury and DO (r s = 0.85, P < 0.01) (Figure 10a and b, Table 6). Widespread distribution of Libellulidae (n = 112) and Chironomidae (n = 109) was found to strongly correlate with total dissolved solids (r = 0.43, P < 0.05) and SS (r = 0.31, P < 0.05), on Axis 1. Other taxa, namely Gerridae, Gomphidae, and Hydropsychidae, on the upper left of the CCA diagram, were within the tolerable range of surface water temperature in the severe condition zone (downstream).
4 Discussion
4.1 Seasonal trends in benthic invertebrate composition and abundance distribution pattern in the Wewe River
Studies of seasonal dynamics in benthic invertebrate communities among smaller lotic systems have been reported by a number of ecologists [58,59,60,61]. Recently, attempts have been made to examine seasonal or short-term temporal variations in invertebrates among a number of large rivers [62]. The findings in this study showed that spatiotemporal changes in water quality in the severe condition might have contributed to highest invertebrate diversity in the severe and intact condition zones, during the wet and dry seasons, respectively, and intact zone during the dry season. For instance, the high amount of DO concentration (4.9–5.0 mg/L) and the optimal water temperature (26.1–28.1°C) in these two zones appeared to favour invertebrate abundance, richness, and evenness distribution (Table 4). Secondly, the presence of rarer taxa (e.g. Chironomidae, Dytiscidae, Gerrida, and Odontopygidae), particularly in the intact zone, probably contributed to highest diversity during the dry season (Table 2). Diversity is considered as a composite index that combines proportional number of individuals, richness, and evenness distribution [49,63] and have been widely used to measure the ecological integrity of ecosystems worldwide. Apart from this composite index for measuring diversity, the concept of species rarity has widely been used to determine a species’ contribution to the diversity (represented by species’ number, abundance, and range area) [64].
Studies in an intermittent river in North Africa found similar variability in diversity of benthic invertebrate community during the dry and wet seasons [25]. However, the high taxa richness in the wet season compared to the dry season contrast the findings of Grohs [61], who reported of higher total richness in macro-invertebrate in summer and fall, compared with winter and spring in the Missouri River. These differences may be because of variations in seasonal habitat conditions in the two biogeographical zones.
Higher taxa abundance in the wet season was thought to coincide with the larval developmental stages and differing responses of taxa to changes in water quality in the dry and wet seasons’ shift, which is largely driven by seasonal-specific disturbances (i.e. intensity of farming practices mostly in the wet season) and changes in the physical environment of the Wewe River. Norris and Thoms [65] argued that some points of the life cycle of benthic invertebrate communities are inextricably linked to biotic and abiotic stream characteristics. However, Ramírez et al. [60] found increase in insect abundance in the dry season compared to the wet season, in tropical low land streams. The authors attributed this to annual temporal changes in stream physicochemistry related to rainfall, with subsequent changes in discharge.
Low abundance of taxa, especially in the moderate and intact condition zones during the dry season, may occur due to the low levels of concentrated DO, related to low water volume and flow rate (Figure 2, Tables 2 and 4). Extreme dryness is a common phenomenon observed in Ghana, during the peak of the dry season (i.e. January–March). This seasonal phenomenon tends to increase evapotranspiration in many rivers and streams including the Wewe River where this study was conducted. Studies in North Africa have found low benthic taxa abundance to be common in many intermittent streams [25,66,67]. These streams are largely characterized by frequent absence of flow and insufficient water levels during peak flows [25,66,67]. These unstable environmental conditions have the tendency to influence water temperature variability and which may in turn affect smaller instars of insects, ionic concentration, and pH [25]. With air temperature projected to increase (2.5°C) over most parts of Africa by the end of the twenty-first century [68], water temperature is likely to increase. This condition could further affect DO levels in most African freshwater systems, including the Wewe River, where this study was conducted. Thus, benthic taxa noted for drought tolerance condition (e.g. Chironomidae and Libellulidae) [69,70,71] may go extinct.
Changes in stream conditions as a result of seasonality and other direct anthropogenic drivers such as farming, dumping of solid waste in streams, and burning can affect some benthic community structure that are sensitive extreme habitat perturbation, and this probably explained why Corydalidae, Notonemouridae, and Protoneuridae were only found during the dry season, whereas Belostomatidae, Gomphidae, and Neptidae thrived only in wet season (Figures 9a and b and 10a and b). The dominance of Nepidae (water scorpion) in the wet season was probably because of their broad range habitat preferences (i.e. ditches, muddy sections of streams, and water area with dead leaves and twigs) (Figure 10a and b). Finally, the presence of the second most dominant taxa like Chironomidae and Libellulidae equally suggests their ability to inhabit or adapt to different hydrologic conditions. Chironomidae and Libellulidae are known to exist in all freshwater wetland types and, are microhabitat selective and play a vital role in food webs [72]. Thus, their large populations are facilitated by the high productivity of freshwater wetlands [72]. Other studies also revealed that Chironomids are also known to be tolerant to disturbances [70,71]. In coastal rivers of southeast Ivory Coast (a neighbouring country on the western border of Ghana), Edia et al. [73] found Chironomidae among the richest taxon diversity in slightly disturbed environment. Thus, Chironomids could be used as an indicator of severe freshwater habitat transformation, giving its resilience to disturbances.
4.2 Seasonal physicochemical influence on benthic invertebrate across the three condition zones
Benthic habitats are complex, and a variety of environmental variables acting at multiple spatial scales regulate the composition and distribution patterns of stream macro-invertebrate assemblages in a synergistic fashion [74,75]. For example, latitude, longitude, pH, and stream characteristics like water velocity, width and depth, substrate composition, and concentrations of nutrients and dissolved organic carbon are often important determinants of macro-invertebrate community structure and may also drive patterns in benthic community composition [63,76,77,78,79]. In this study, DO concentration, surface water temperature, TDS, SS, EC, and pH were the physicochemical drivers that influenced higher abundance of benthic invertebrates in the wet season than in the dry season (Figures 9a and b and 10a and b). These physicochemical drivers were probably at optimal levels and within tolerable limit for benthic invertebrates.
In a similar study on seasonal variations in benthic invertebrates, Dudgeon [12] found hydrologic regime and geomorphology in stream environments, as the key determinants of their distribution and abundance. Using multivariate approach, CCA, Jonsson et al. [80] listed pH, water velocity, organic matter, and low canopy openness as the principal predictors of benthic invertebrate community structure in boreal streams. The impact of these predictive factors that cause changes in aquatic environmental conditions may lead to the loss of species, altered community composition, and homogenization of communities [81]. The aftermath of these effects according to Meyer et al. [81] may result in reduced biodiversity and consequently impair the functioning of these habitats [82].
Impact of higher levels of mercury concentration and SS on the low abundance and distribution of Asellidae and Perlidae especially in the moderate condition zone was probably a result of intensive use of agrochemicals on nearby farmlands, deposition of solid waste on the riparian zone, and the flow of liquid waste from surrounding human settlements, during the wet season. An increase in mercury concentration might have contributed to oxygen depletion, which consequently reduced Asellidae population. Such negative impacts on the functional role of invertebrates as litter decomposers could be impaired and consequently have long-term effects on the ecosystem health of the Wewe River, leading to loss of biodiversity. Hellawell [83] revealed that Perlidae and Ephemeridae are among the intolerant groups, and this tends to reflect in their low numbers usually in the agricultural streams. Getwongsa et al. [84] also found Perlidae to decrease in agricultural streams. Thus, with the current rate of farming expansion along the fringes of the Wewe River, there is the likelihood of increase in mercury concentration in the water column (Table 4), through surface run-off during the wet season. Thus, sensitive benthic taxa such as Assellidae and Perlidae [85] could be threatened by this disturbance-related driver. Expansion of cultivated lands along agroecological zones in the humid highlands of Ethiopia was found to have an impact on ecosystem like freshwater systems [86]. This suggests that aquatic life along these agroecological zones may as well be at risk of extinction as a result of these human-led disturbances.
5 Conclusion
This study assessed the seasonal response of benthic invertebrate to physicochemical drivers in the Wewe River. Our findings showed that benthic invertebrate assemblages did not differ in the dry and wet seasons. Nonetheless, there was a slight increase among individuals, taxa richness, and diversity in the wet season compared with the dry season. The most severely disturbed segment (downstream) of the Wewe River registered the highest number of individuals, suggesting the role of habitat perturbation and sample size in influencing macro-invertebrate heterogeneity and abundance. Dominance of Chironomidae in the dry and wet seasons was attributed to their broad range habitat preferences and their ability to adapt to seasonal changes in physicochemical driver conditions, while the lowest abundance of Asellidae and Perlidae was probably due to their sensitivity to elevated levels of some water quality parameters, namely mercury and low dissolve oxygen concentration, which were characteristics of the moderately disturbed zone (upstream) of the Wewe River. Thus, for effective management of the Wewe River, we recommend that Asellidae and Perlidae be considered as a suit of indicator benthic orders to monitor the water quality health. Other conservation measures that should be considered in protecting the overall ecosystem of the Wewe River include the following:
farming activities within the riparian zone should be banned and farmers relocated in places further away from the riparian zones;
diversion of liquid sewage spillway emptying into the river course; and
dumping of solid refuse along the riparian zone must be banned by authorities and managers of the Wewe River.
In terms of the setbacks that arose from this study, we noted the difficulty in sampling with less than four persons per segment of the river per day. The extreme stress or tiredness observed among the researchers may have resulted in the lack of detection or loss of some benthic invertebrate samples, as we could not sample every section of the different condition zones of the riverine continuum. We suggest that future studies should involve not less than seven persons, sampling in each segment of a riverine continuum.
Acknowledgements
The authors wish to thank Madam Gloria (Lab Technician in the Faculty of Renewable Natural Resources) for permitting the research team to use the laboratory for invertebrate identification.
-
Funding information: This research was funded by the KreF Fund of Kwame Nkrumah University of Science & Technology.
-
Author contributions: C.A.N. and S.K.O. conceived the ideas and designed the methodology; C.A.N., S.K.O., and G.K.B. carried out field data collection, identification of benthic taxa, and data entry in excel sheet; C.A.N. and G.K.B. analysed the data and led the writing of the manuscript. S.K.O. undertook in-house review of the drafts and gave final approval for submission.
-
Conflict of interest: The authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
[1] Balachandran C, Ramachandra T. Aquatic macroinvertebrate diversity and water quality of Bangalore lakes. Wetlands: Biodiversity and Climate Change. Bangalore: Energy and Wetland Research Group; 2010.Suche in Google Scholar
[2] Andersen AN, Fisher A, Hoffmann BD, Read JL, Richards R. Use of terrestrial invertebrates for biodiversity monitoring in Australian rangelands, with particular reference to ants. J Aust Ecol. 2004;20429(1):87–92.10.1111/j.1442-9993.2004.01362.xSuche in Google Scholar
[3] Jun Y-C, Kim N-Y, Kim S-H, Park Y-S, Kong D-S, Hwang S-J. Spatial distribution of Benthic macroinvertebrate assemblages in relation to environmental variables in Korean nationwide streams. Water. 2016;8(1):27. http://www.mdpi.com/2073-4441/8/1/2710.3390/w8010027Suche in Google Scholar
[4] Mereta ST, Boets P, Bayih A, Malu A, Ephrem Z, Sisay A, et al. Analysis of environmental factors determining the abundance and diversity of macroinvertebrate taxa in natural wetlands of Southwest Ethiopia. Ecol Inform. 2012;7(1):52–61. 10.1016/j.ecoinf.2011.11.005.Suche in Google Scholar
[5] Buss DF, Baptista DF, Silveira MP, Nessimian JL, Dorvillé LF. Influence of water chemistry and environmental degradation on macroinvertebrate assemblages in a river basin in south-east Brazil. Hydrobiol. 2002;481(1–3):125–36.10.1023/A:1021281508709Suche in Google Scholar
[6] Al-Shami SA, Rawi CSM, Ahmad AH, Hamid SA, Nor SAM. Influence of agricultural, industrial, and anthropogenic stresses on the distribution and diversity of macroinvertebrates in Juru River Basin, Penang, Malaysia. Ecotoxicol Environ Saf. 2011;74(5):1195–202.10.1016/j.ecoenv.2011.02.022Suche in Google Scholar
[7] Bott TL, Brock JT, Dunn CS, Naiman RJ, Ovink RW, Petersen RC. Benthic community metabolism in four temperate stream systems: an inter-biome comparison and evaluation of the river continuum concept. Hydrobiol. 1985;123(1):3–45.10.1007/BF00006613Suche in Google Scholar
[8] Merz JE, Chan LKO. Effects of gravel augmentation on macroinvertebrate assemblages in a regulated California river. River Res Appl. 2005;21(1):61–74. 10.1002/rra.819.Suche in Google Scholar
[9] Covich AP, Crowl TA, Heartsill-Scalley T. Effects of drought and hurricane disturbances on headwater distributions of palaemonid river shrimp (Macrobrachium spp.) in the Luquillo Mountains, Puerto Rico. North Amer Benthol Soc. 2006;25(1):99–107.10.1899/0887-3593(2006)25[99:EODAHD]2.0.CO;2Suche in Google Scholar
[10] Henriques-Oliveira AL, Nessimian JL. Aquatic macroinvertebrate diversity and composition in streams along an altitudinal gradient in Southeastern Brazil. Biota Neotrop. 2010;10(3):115–28.10.1590/S1676-06032010000300012Suche in Google Scholar
[11] Haro RR, Johnson L, Host G. Catchment and reach-scale properties as indicators of macroinvertebrate species traits. Freshw Biol. 1997;37(1):219–30.10.1046/j.1365-2427.1997.d01-540.xSuche in Google Scholar
[12] Dudgeon D. The ecology of tropical Asian rivers and streams in relation to biodiversity conservation. Ann Rev Ecol Syst. 2000;31(1):239–63.10.1146/annurev.ecolsys.31.1.239Suche in Google Scholar
[13] Graça MA, Pinto P, Cortes R, Coimbra N, Oliveira S, Morais M, et al. Factors affecting macroinvertebrate richness and diversity in Portuguese streams: a two-scale analysis. Int Rev Hydrobiol. 2004;89(2):151–64.10.1002/iroh.200310705Suche in Google Scholar
[14] Sharma RC, Rawat JS. Monitoring of aquatic macroinvertebrates as bioindicator for assessing the health of wetlands: a case study in the central Himalayas, India. Ecol Indic. 2009;9(1):118–28.10.1016/j.ecolind.2008.02.004Suche in Google Scholar
[15] Gore JA, Layzer JB, Mead JIM. Mead, Macroinvertebrate instream flow studies after 20 years: a role in stream management and restoration. Reg Rivers Res Manag Int J Devot River Res Manag. 2001;17(4–5):527–42.10.1002/rrr.650Suche in Google Scholar
[16] Brown LE, Céréghino R, Compin A. Endemic freshwater invertebrates from southern France: diversity, distribution and conservation implications. Biol Conserv. 2009;142(11):2613–9.10.1016/j.biocon.2009.06.009Suche in Google Scholar
[17] Mabidi A, Mathews SB, Perissinotto R. Distribution and diversity of aquatic macroinvertebrate assemblages in a semi-arid region earmarked for shale gas exploration (Eastern Cape Karoo, South Africa). PLoS One 2012. 2017;12(6):e0178559.10.1371/journal.pone.0178559Suche in Google Scholar PubMed PubMed Central
[18] Abungba JA, Khare D, Pingale SM, Adjei KA, Gyamfi C, Odai SN. Assessment of hydro-climatic trends and variability over the Black Volta Basin in Ghana. J Earth Syst Environ. 2020;4:739–55. 10.1007/s41748-020-00171-9.Suche in Google Scholar
[19] Fu BJ, Zhuang XL, Jiang GB, Shi JB, Lü YH. Environmental problems and challenges in China. Environ Sci Technol. 2007;41:7597–602.10.1021/es072643lSuche in Google Scholar PubMed
[20] Qin B, Xu P, Wu Q, Luo L, Zhang Y. Environmental issues of Lake Taihu. China Hydrobiol. 2007;581(1):3–14. 10.1007/978-1-4020-6158-5_2.Suche in Google Scholar
[21] Mensah-Bonsu IF, Owusu-Ansah JK. State of the environment in Kumasi. In: Adarkwa KK, editor. The future of the tree: towards growth and development of Kumasi. Upk, Kumasi; 2011. p. 174–94.Suche in Google Scholar
[22] Forkuor D, Callistus M, Kyei PK, Forkuor G. Changes in land use in the Kumasi metropolis of Ghana – whose fault? Local Gov Stud. 2012;4(1):58–77.Suche in Google Scholar
[23] Agyapong EB, Ashiagbor G, Nsor CA, van Leeuwen LM. Urban land transformations and its implication on tree abundance distribution and richness in Kumasi. Ghana Urban Ecol. 2018;4(1):1–11. 10.1093/jue/juy019.Suche in Google Scholar
[24] Forkuor D, Otieku E. Environmental consciousness and practices of land users along urban water bodies in Ghana: the case of Kumasi Metropolis. Ethiopian Environ Stud Manag. 2015;8(Suppl. 2):966–75. 10.4314/ejesm.v8i2.10S.Suche in Google Scholar
[25] Arab A, Lec S, Lounaci A, Park YS. Spatial and temporal patterns of benthic invertebrate communities in an intermittent river (North Africa). Int J Limnol. 2004;40(4):317–27.10.1051/limn/2004029Suche in Google Scholar
[26] Lounaci A, Brosse S, Thomas A, Lek S. Abundance, diversity and community structure of macroinvertebrates in an Algerian stream: the Sébaou wadi. Ann Limnol Int J Limnol. 2000;36:123–33.10.1051/limn/2000008Suche in Google Scholar
[27] Gasith A, Resh VH. Streams in Mediterranean climate regions: abiotic influences and biotic responses to predictable seasonal events. Ann Rev Ecol Syst. 1999;30:51–81.10.1146/annurev.ecolsys.30.1.51Suche in Google Scholar
[28] Kalejta B, Hockey PAR. Distribution, abundance and productivity of benthic invertebrates at the Berg River estuary, South Africa. Estuar Coast Shelf Sci. 1991;33(2):175–91. 10.1016/0272-7714(91)90005-V.Suche in Google Scholar
[29] Niba A, Sakwe S. Turnover of benthic macroinvertebrates along the Mthatha River, Eastern Cape, South Africa: implications for water quality bio-monitoring using indicator species. Freshw Ecol. 2018;33(1):157–1717. 10.1080/02705060.2018.14319692018.Suche in Google Scholar
[30] Nwankwo CB, Hoque MA, Islam MA, Dewan A. Groundwater constituents and trace elements in the basement aquifers of Africa and sedimentary aquifers of Asia: medical hydrogeology of drinking water minerals and toxicants. J Earth Syst Env. 2020;4(2):369–84. 10.1007/s41748-020-00151-z.Suche in Google Scholar
[31] Pires AM, Cowx IG, Coelho MM. Benthic macroinvertebrate communities of intermittent streams in the middle reaches of the Guadiana Basin (Portugal). Hydrobiol. 2000;435(1–3):167–75.10.1023/A:1004003726283Suche in Google Scholar
[32] Tay SC, Kye-Duodu G, Gbedema SY. River Wiwi: a source of Schistosoma haematobium infection in school children in Kumasi, an urban African setting. African. J Macrobiol Res. 2013;7(20):2213–20.10.5897/AJMR12.489Suche in Google Scholar
[33] Ghana Meteorological Agency (GMet). The head Office, P. O. Box LG 87, Legon, Accra, GR, Ghana; 2008.Suche in Google Scholar
[34] Stein ED, White BP, Mazor RD, Miller PE, Erik M, Pilgrim EM. Evaluating ethanol-based sample preservation to facilitate use of DNA barcoding in routine freshwater biomonitoring programs using benthic macroinvertebrates. PLoS One. 2013;8(1):e51273. 10.1371/journal.pone.0051273.Suche in Google Scholar PubMed PubMed Central
[35] Brinkman MA, Duffy WG. Evaluation of four wetland aquatic invertebrate samplers and four sample sorting methods. Freshw Ecol. 1996;11:193–200.10.1080/02705060.1996.9663478Suche in Google Scholar
[36] Bobson M, Pawley S, Fletcher M, Powell A. Guide to freshwater invertebrates. Cumbria, UK: Freshwater Biological Association Scientific Publication; 2012.Suche in Google Scholar
[37] Dudgeon D. Tropical Asian streams: zoobenthos, ecology and conservation. Hong Kong: Hong Kong University Press; 1999.Suche in Google Scholar
[38] Hair JF, Black WC, Babin BJ, Anderson RE, Tatham RL. Análise multivariada de dados. Bookman Editora; 2009.Suche in Google Scholar
[39] Magurran AE. Measuring biological diversity. Oxford: Blackwell Science; 2004.Suche in Google Scholar
[40] Fattorini S. A simple method to fit geometric series and broken stick models in community ecology and island biogeography. Acta Oecol. 2005;28(3):199–205.10.1016/j.actao.2005.04.003Suche in Google Scholar
[41] Fattorini S. Species ecological preferences predict extinction risk in urban tenebrionid beetle guilds. Anim Biol. 2013;63:93–106. 10.1163/15707563-00002396.Suche in Google Scholar
[42] Fattorini S, Rigal F, Cardoso P, Borges PAV. Using species abundance distribution models and diversity indices for biogeographical analyses. Acta Oecol. 2016;70:21–8. 10.1016/j.actao.2015.11.003.Suche in Google Scholar
[43] Colwell RK, Gotelli NJ. Quantifying biodiversity: procedures and pitfalls in the measurement and comparison of species richness. Ecol Lett. 2001;4(4):379–91.10.1046/j.1461-0248.2001.00230.xSuche in Google Scholar
[44] Gray JS, Mirza FBA. Possible method for the detection of pollution-induced disturbance on marine benthic communities. Mar Pollut Bull. 1979;10:142–6.10.1016/0025-326X(79)90082-1Suche in Google Scholar
[45] Colwell RK, Coddington JA. Estimating terrestrial biodiversity through extrapolation. Philos Trans R Soc Lond B Biol Sci. 1994;345:101–18. 10.1098/rstb.1994.0091.Suche in Google Scholar PubMed
[46] Colwell RK. EstimateS: statistical estimation of species richness and shared species from samples. Version 60; 2000. http://viceroy.eeb.uconn.edu/estimatesSuche in Google Scholar
[47] King JR, Porter S. Evaluation of sampling methods and species richness estimators for ants in upland ecosystems in Florida. Environ Entomol. 2005;34(6):1566–78. 10.1603/0046-225X-34.6.1566.Suche in Google Scholar
[48] McArthur RH. Patterns of species diversity. Biol Rev. 1965;40(4):510–33. 10.1111/j.1469185X.1965.tb00815.x Suche in Google Scholar
[49] Hill MO. Diversity and evenness: a unifying notation and its consequences. Ecol. 1973;54:427–32. 10.2307/1934352 Suche in Google Scholar
[50] Jost L. Entropy and diversity. Oikos. 2006;113:363–75.10.1111/j.2006.0030-1299.14714.xSuche in Google Scholar
[51] Jost L. Partitioning diversity into independent alpha and beta components. Ecol. 2007;88:2427–39.10.1890/06-1736.1Suche in Google Scholar
[52] Chao A, Gotelli NJ, Hsieh TC, Sande EL, Ma KH, Colwell RK, et al. Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. J Ecol Monogr. 2014;84(1):45–67.10.1890/13-0133.1Suche in Google Scholar
[53] Ter Braak CJF. Canonical correspondence analysis: a new eigenvector technique for multivariate direct gradient analysis. Ecol. 1986;67(5):1167–79.10.2307/1938672Suche in Google Scholar
[54] Kent M, Coker P. Vegetation description and analysis: a practical approach. New York, NY, USA: John Wiley and Sons Ltd; 2002.Suche in Google Scholar
[55] Dorugade AV, Kashid DN. Alternative method for choosing ridge parameter for regression. Appl Math Sci. 2010;4(9):447–56.Suche in Google Scholar
[56] Schreiber-Gregory DN, Jackson HM. Multicollinearity: what is it, why should we care, and how can it be controlled?. Env’tal Sc. 2017. p. 1–12.Suche in Google Scholar
[57] Hammer Ø, Harper DAT, Ryan PD. PAST: paleontological statistics software package for education and data analysis. Palaeontol Electron. 2001;4(1):9.Suche in Google Scholar
[58] Modde TC, Schmulbach JC. Seasonal changes in the drift and benthic macroinvertebrates in the unchannelized Missouri River in South Dakota. Proc South Dak Acad Sci. 1973;52:118–26.Suche in Google Scholar
[59] Barnum J, Bachmann RW. Benthic macroinvertebrate habitat associations of the channelized middle Missouri River. Iowa Acad Sci: JIAS. 1988;95(2):60–5.Suche in Google Scholar
[60] Ramírez A, Pringle CM, Douglas M. Temporal and spatial patterns in stream physicochemistry and insect assemblages in tropical lowland streams. J North Am Benthol Soc. 2006;25(1):108–25. 10.1899/0887-3593(2006)25[108:TASPIS]2.0.CO;2.Suche in Google Scholar
[61] Grohs KL. Macroinvertebrate composition and patterns of prey use by juvenile pallid sturgeon (Scaphirhynchus albus) in the Missouri River, South Dakota and Nebraska. Master’s thesis. Brookings (SD): South Dakota State University; 2008.Suche in Google Scholar
[62] McCord SB, Kuhl BA. Macroinvertebrate community structure and its seasonal variation in the Upper Mississippi River, USA: case study. Freshw Ecol. 2013;28(1):63–78. 10.1080/02705060.2012.693458.Suche in Google Scholar
[63] Maturo F, Di Battista T. A functional approach to Hill’s numbers for assessing changes in species variety of ecological communities over time. Ecol Indic. 2018;84:70–81. 10.1016/j.ecolind.2017.08.016.Suche in Google Scholar
[64] Kondratyeva A, Grandcolas P, Pavoin S. Reconciling the concepts and measures of diversity, rarity and originality in ecology and evolution. Biol Rev. 2019;94:1317–37. 10.1111/brv.12504.Suche in Google Scholar
[65] Norris RH, Thoms MC. What is river health? Freshw Biol. 1999;41:197–209.10.1046/j.1365-2427.1999.00425.xSuche in Google Scholar
[66] Belaidi N, Taleb J, Gagneur J. Composition and dynamics of hyporheic and surface fauna in a semi-arid stream in relation to the management of a polluted reservoir. Ann Limnol Int J Limnol. 2004;40:237–48.10.1051/limn/2004020Suche in Google Scholar
[67] Pires AM, Cowx IG, Coelho MM. Benthic macroinvertebrate communities of intermittent streams in the middle reaches of the Guadiana Basin (Portugal). Hydrobiol. 2000;435:167–75.10.1023/A:1004003726283Suche in Google Scholar
[68] Almazroui M, Saeed F, Islam MN, Ismail M, Klutse NAB, et al. Projected change in temperature and precipitation over Africa from CMIP6. Earth Syst Env. 2020;4:455–75. 10.1007/s41748-020-00161-x.Suche in Google Scholar
[69] Hadjoudj S, Khelifa R, Guebailia A, Amari H, Hadjadji S, Zebsa R, et al. Emergence ecology of Orthetrum cancellatum: temporal pattern and microhabitat selection (Odonata: Libellulidae). Ann de la Soc Entomol de Fr (NS). 2014;50(3–4):343–9. 10.1080/00379271.2014.938941.Suche in Google Scholar
[70] Collier KJ, Bowman EJ. Role of wood in pumice-bed streams - I: Impacts of post-harvest management on water quality, habitat and benthic invertebrates. For Ecol Mgt. 2003;177:243–59.10.1016/S0378-1127(02)00447-4Suche in Google Scholar
[71] Martel N, Rodriguez MA, Berube P. Multi-scale analysis of responses of stream macrobenthos to forestry activities and environmental context. Freshw Biol. 2007;52:85–97.10.1111/j.1365-2427.2006.01674.xSuche in Google Scholar
[72] Keiper JB, Walton WE, Foote BA. Biology and ecology of higher diptera from freshwater wetlands. Ann Rev Entomol. 2002;47:207–32. 10.1146/annurev.ento.47.091201.145159.Suche in Google Scholar PubMed
[73] Edia EO, Brosse S, Ouattara A, Gourene G, Winterton P, Lek-Ang S. Aquatic insects’ assemblage patterns in four West-African coastal rivers. Biol Sci. 2007;7(7):1130–8.10.3923/jbs.2007.1130.1138Suche in Google Scholar
[74] Bae MJ, Kwon Y, Hwang SJ, Con TS, Yang HJ, Kwak I-S, et al. Relationships between three major stream assemblages and their environmental factors in multiple spatial scales. Ann Limnol Int J Limnol. 2011;47:S91–105.10.1051/limn/2011022Suche in Google Scholar
[75] Braccia A, Voshell JR. Environmental factors accounting for benthic macroinvertebrate assemblage structure at the sample scale in streams subjected to a gradient of cattle grazing. Hydrobiol. 2006;573:55–73.10.1007/s10750-006-0257-2Suche in Google Scholar
[76] Heino J, Muotka T, Paavola R. Determinants of macroinvertebrate diversity in headwater streams: regional and local influences. Anim Ecol. 2003;72:425–34.10.1046/j.1365-2656.2003.00711.xSuche in Google Scholar
[77] Heino J, Ilmonen J, Paasivirta L. Continuous variation of macroinvertebrate communities along environmental gradients in northern streams. Boreal Environ Res. 2014;201419:21–38.Suche in Google Scholar
[78] Schmera D, Erős T, Heino J. Habitat filtering determines spatial variation of macroinvertebrate community traits in northern headwater streams. Comm Ecol. 2013;14(1):77–88.10.1556/ComEc.14.2013.1.9Suche in Google Scholar
[79] Göthe E, Friberg N, Kahlert M, Temnerud J, Sandin L. Headwater biodiversity among different levels of stream habitat hierarchy. Biodivers Conserv. 2014;23(1):63–80.10.1007/s10531-013-0584-3Suche in Google Scholar
[80] Jonsson M, Burrows RM, Lidman J, Fàltstróm E, Laudon H, Sponseller RA. Land use influences macroinvertebrate community composition in boreal headwaters through altered stream conditions. Ambio. 2017;46:311–23. 10.1007/s13280-016-0837-y.Suche in Google Scholar PubMed PubMed Central
[81] Meyer JL, Strayer DL, Wallace JB, Eggert SL, Helfman GS, Leonard NL. The contribution of headwater streams to biodiversity in river networks. Amer Water Res Assoc. 2007;43:86–103.10.1111/j.1752-1688.2007.00008.xSuche in Google Scholar
[82] Vaughn CC. Biodiversity losses and ecosystem functioning in freshwaters: Emerging conclusions and research directions. BioScience. 2010;60:25–35.10.1525/bio.2010.60.1.7Suche in Google Scholar
[83] Hellawell JM, editor. Biological indicators of freshwater pollution and environmental management. In: Melanby K, editor. Pollution monitoring series; 2012. p. 546. 10.1007/978-94-009-4315-5.Suche in Google Scholar
[84] Getwongsa P, Hanjavanit C, Sangpradub N. Impacts of agricultural land use on stream benthic macroinvertebrates in tributaries of the Mekong River, northeast Thailand. Adv Environ Sci Int J Bioflux Soc AES Bioflux. 2010;2(2):97–112. http://www.aes.bioflux.com.ro.Suche in Google Scholar
[85] Kaller MD, Hartman KJ. Evidence of a threshold level of fine sediment accumulation for altering benthic macroinvertebrate communities. Hydrobiol. 2004;518:95–104.10.1023/B:HYDR.0000025059.82197.35Suche in Google Scholar
[86] Negash E, Getachew T, Birhane E, Gebrewahed H. Ecosystem Service value distribution along the agroecological gradient in north-central Ethiopia. Earth Syst Environ. 2020;4:107–16. 10.1007/s41748-020-00149-7.Suche in Google Scholar
© 2021 Samuel K. Oppong et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells
Artikel in diesem Heft
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells