ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
-
Xinyu Tan
 and
Xinping Xu
and
Xinping Xu

Abstract
Cell proliferation and epithelial–mesenchymal transition (EMT) are two common tumor phenotypes closely linked to malignant tumor progression. Genes regulating tumor progression often exhibit consistent regulatory trends in these phenotypes; however, certain genes may display inconsistent regulatory patterns in tumor proliferation and EMT. In this investigation, initial transcriptomic and tumor database analyses revealed that alcohol dehydrogenase 5 (ADH5) is downregulated in non-small cell lung cancer (NSCLC) tissues and correlates negatively with NSCLC prognosis. Subsequent experimental manipulation of ADH5 levels in tumor cells demonstrated that ADH5 overexpression decreased proliferation while enhancing migration and invasion capacities in NSCLC cells. Moreover, ADH5 overexpression hindered xenograft tumor growth in nude mice. However, ADH5 knockdown yielded contrasting outcomes by stimulating NSCLC cell proliferation while impeding migration and invasion abilities. Notably, ADH5 overexpression triggered EMT through Smad2/Smad3 activation, leading to the upregulation of SRY-Box Transcription Factor 9. TGFbetaR1/ALK5 inhibitor SB431542 was able to alleviate the effects of ADH5 overexpression on NSCLC cells. This study indicates a critical role of ADH5 in tumors associated with cancer cell growth inhibition but EMT activation. These findings underscore ADH5 as a potential regulator of NSCLC cell plasticity, emphasizing its promise as a therapeutic target for NSCLC management.
1 Introduction
Non-small cell lung cancer (NSCLC), the predominant histological subtype of lung cancer, ranks as the second most common and deadly cancer globally [1]. Although significant advances have been made in the treatments of NSCLC, including targeted therapy and immune therapy, the prognosis still falls short due to the ongoing evolution of cancer cells. Despite the positive response to treatment at the beginning, the eventual emergence of drug resistance and immune evasion leads to the malignancy of cancer cells. Unravelling the mechanisms through which cancer cells modulate their phenotypes to gain advantages in motility and proliferation during cancer progression remains a challenge.
Alcohol dehydrogenase 5 (ADH5) is a member of the alcohol dehydrogenase family and responsible for metabolizing intracellular S-nitrosoglutathione, which indirectly regulates protein S-nitrosylation [2]. By reducing protein S-nitrosylation, ADH5 functions as an antagonist of nitric oxide, a molecule is known for its dual roles in biological processes and tumorigenesis [3,4]. Formaldehyde dehydrogenase and ADH5 were discovered to be one enzyme in 1989 [5]. Aldehyde-driven DNA damage leading to AMeD syndrome and Cockayne syndrome, has been found to be the consequence of digenic mutations in ADH5 and aldehyde dehydrogenase 2 or Cockayne syndrome protein B [6,7]. The modulation of S-nitrosylation and formaldehyde by ADH5 has been implicated in various fields including hepatocellular carcinoma, cardiovascular diseases, and senescence [8–17], and mainly functions as a protector of the cell homeostasis. However, the integral role of ADH5 in the progression of NCSLC still remains to be elucidated.
Here, this study revealed that ADH5 is downregulated in NSCLC tissues and cell lines. ADH5 overexpression resulted in reduced proliferation but increased migration and invasion of NSCLC cells; whereas, ADH5 knockdown led to enhanced proliferation and reduced migration and invasion. Further investigations demonstrated that overexpression of ADH5 activated Smad2/Smad3 signaling and induced epithelial–mesenchymal transition (EMT) in NSCLC cells. We then applied TGFbetaR1/ALK5 inhibitor SB431542 to rescue the consequence of ADH5 overexpression confirming the regulation of Smad2/3 by ADH5. Moreover, a positive correlation was observed between ADH5 and SRY-Box Transcription Factor 9 (SOX9), with ADH5 upregulating SOX9 expression in NSCLC cells. These findings suggest that ADH5 may serve as a novel regulator of cancer cell plasticity in NSCLC.
2 Materials and methods
2.1 Tissue samples and mice
NSCLC tissues and paired adjacent tissues were collected from patients who were histologically diagnosed at First Affiliated Hospital of Nanchang University between 2020 and 2022. All patients signed informed consent for the research. This study has been approved by the Human Ethics Committee of First Affiliated Hospital of Nanchang University (CDYFYYLK(10-008)). Balb/c nude mice were housed in a pathogen-free laboratory animal center at First Affiliated Hospital of Nanchang University. Animal experiments were scrutinized and approved (CDYFY-IACUC-202210QR002).
-
Informed consent: Informed consent has been obtained from all individuals included in this study.
-
Ethical approval: The research related to human use has been complied with all the relevant national regulations, institutional policies and in accordance with the tenets of the Helsinki Declaration, and has been approved by the Human Ethics Committee of First Affiliated Hospital of Nanchang University (CDYFYYLK(10-008)).
-
Ethical approval: The research related to animal use has been complied with all the relevant national regulations and institutional policies for the care and use of animals, and has been approved by the Ethical Committee of The First Affiliated Hospital of Nanchang University (CDYFY-IACUC-202210QR002).
2.2 Tumor formation in nude mice
PC9 cells overexpressing ADH5 and PC9 cells containing the relative empty vector were prepared in 100 µL with 4 × 106 cells and injected subcutaneously into male BALB/c nude mice. Tumor volumes were measured and calculated every 3 days according to the formula V = 0.5 × L × W 2. Five weeks later, the nude mice were sacrificed and the tumors were collected for further analysis.
2.3 Cell cultures
The NSCLC cell lines and normal human bronchial epithelial cell line were purchased from ATCC. Cells were cultured in RPMI-1640 (Gibco) or Dulbecco’s modified Eagle’s medium (Gibco) supplemented with 10% fetal bovine serum (FBS), 100 U/mL penicillin, and 100 mg/mL streptomycin in incubators at 37°C with 5% carbon dioxide.
2.4 Construction of stable cell lines
H358 and PC9 cells were transfected with ADH5 expression plasmids (Origene) using lipofectamine 3000 (Invitrogen) according to the manufacturer’s instructions. Two days after the transfection, neomycin was applied for a week. Limited dilution assay was performed to acquire monoclonal ADH5 overexpression cells. Validation of overexpression via western blot confirmed two H358 monoclonal overexpression cell lines and one PC9 monoclonal overexpression cell line. shRNA plasmids for ADH5 and corresponding negative control (Origene) were transfected into the 293 T cell. After 48 h incubation, virus-containing supernatants were collected for transfection into H358 and PC9 cells. Puromycin was applied for 3 days. Knock-down was confirmed by western blot.
2.5 qRT-PCR
TRIzol reagent (TransGen Biotech) was used to extract total RNA from cells and the purity and integrity were assessed by Nanodrop 2000/2000C spectrophotometry (Thermo Fisher Scientific). Total RNA was reversely transcribed to high-quality cDNA using All-in-One First-Strand cDNA Synthesis SuperMix for qPCR (TransGen Biotech). qRT-PCR was performed using SYBR Green Mix (Thermo Fisher Scientific). Results were normalized to the expression of glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (Table 1).
Primers used in qRT-PCR
| Gene | Forward (5′–3′) | Reverse (5′–3′) |
|---|---|---|
| GAPDH | ACCACAGTCCAT GCCATCAC | TCCACCACCCTGTTGCTGTA |
| ADH5 | ATGGCGAACGAGGTTATCAAG | CATGTCCCAAGATCACTGGAAAA |
| c-Myc | TCTCCTTGCAGCTGCTTAG | GTCGTAGTCGAGGTCATAG |
| P15 | CACCGTTGGCCGTAAACTTAAC | TAATGAAGCTGAGCCCAGGTCT |
2.6 Western blotting
Total proteins were collected with ice-cold RIPA lysis buffer and the concentrations were determined by BCA protein reagent kit. Then the proteins were heated at 100°C with protein loading buffer. Samples were subjected to SDS-PAGE and transferred to PVDF membrane. The antibodies used in the study include: β-actin antibody (1/5,000; Proteintech), ADH5 antibody (1/2,000; Abcam), SMAD2/3 antibody (1/5,000; Cell Signaling Technology), phospho-SMAD2/3 (1/2,000; Cell Signaling Technology), E-cadherin antibody (1/2,000; Cell Signaling Technology), N-cadherin antibody (1/2,000; Cell Signaling Technology), Snail antibody (1/2,000; Cell Signaling Technology), Slug antibody (1/2,000; Cell Signaling Technology), AKT antibody (1/5,000; Proteintech), p-AKT antibody (1/2,000; Proteintech), and SOX9 antibody (1/2,000; Proteintech).
2.7 Assays for CCK8 cell proliferation and colony formation
H358 and PC9 cells with knocked down or overexpressed ADH5 were seeded onto 96-well plates (1 × 103 cells per well). CCK8 (Solarbio) was used according to the manufacturer’s instructions. For colony formation, cells were seeded at a density of 1.0 × 103 cells/mL in six-well plates. After 2 weeks’ culture, cells were fixed with 4% paraformaldehyde and stained with crystal violet (Solarbio).
2.8 Transwell assays
Transwell membranes (Corning Costar) were used. For migration assays, 4.0 × 104 cells in 200 μL of RPMI-1640 medium containing 0.1% FBS were added to the upper chambers of transwell plates. Medium containing 10% FBS was added to the bottom chamber. After incubation, the migrated cells were fixed with 4% paraformaldehyde and stained with crystal violet. For invasion assay, 5.0 × 104 cells were seeded to the top chambers coated with Matrigel. After 36 h, the cells were fixed with 4% paraformaldehyde and stained with crystal violet.
2.9 Bioinformatics of public databases
The expression of ADH5 in LUAD was obtained from the Cancer Genome Atlas (TCGA) data visualization web-tool UALCAN (http://ualcan.path.uab.edu/). ADH5 protein expression in normal lung tissues and lung adenocarcinoma was from The Human Protein Atlas (THPA) database (https://www.proteinatlas.org/). Kaplan–Meier analysis of ADH5 in LUAD was acquired from Kaplan–Meier Plotter (http://kmplot.com/). The correlation between ADH5 and SOX9 was obtained from Gepia database (http://gepia.cancer-pku.cn/).
2.10 Statistical analysis
GraphPad Prism 8.0 was used to perform statistical analysis. Numerical data were expressed as mean ± standard deviation. Statistical significance was defined as P < 0.05. Student’s t-test and one-way analysis of variance were used to compare the expression levels of two groups and more, respectively.
3 Results
3.1 ADH5 was downregulated in NSCLC tissues and cell lines
First, we conducted a bioinformatic analysis to examine the expression of ADH5 in NSCLC. Data from the TCGA database indicated a decrease in ADH5 mRNA levels in lung adenocarcinoma (Figure 1a). The protein levels of ADH5 also decreased in the THPA database (Figure 1b), which has been reported in several types of cancers [8,18–20]. Furthermore, the survival probability of LUAD patients with high ADH5 expression was significantly higher than those with low ADH5 expression level (Figure 1c). Subsequently, to validate both the mRNA and protein expression of ADH5, western blot and qRT-PCR were conducted to confirm ADH5 expression in normal cell line and NSCLC cell lines (Figure 1d and e). When compared to the normal human bronchial epithelial cell line BEAS-2B, both the protein and mRNA levels of ADH5 were significantly downregulated in NSCLC cell lines, except H1299. The loss of p53 protein in H1299 may contribute to ADH5 overexpression because p53 directly regulates ADH5 [21]. We further confirmed the mRNA and protein expression of ADH5 in paired NSCLC tissues, revealing lower levels of both mRNA and protein in the NSCLC tissues than that in adjacent normal tissues (Figure 1f and g), given that ADH5 has been reported to be post-transcriptionally regulated [21]. Significant downregulation of mRNA level and protein level of ADH5 were found in both lung cancer cell lines and tissues. These findings imply that ADH5 is downregulated in NSCLC and could potentially contribute to the pathogenesis of the disease.
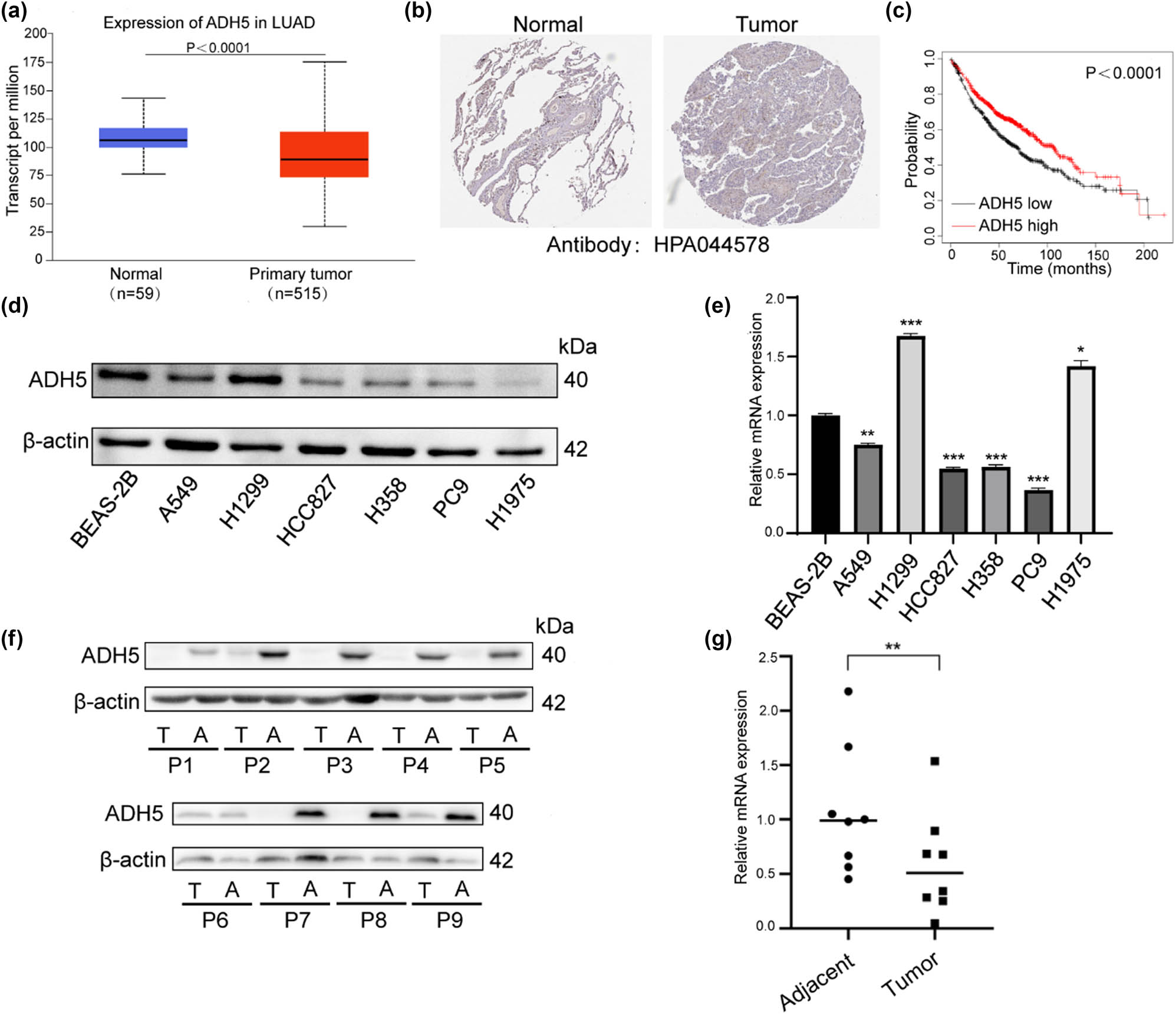
Expression of ADH5 in NSCLC cell lines and tissues. (a) mRNA expression of ADH5 from TCGA database. (b) ADH5 protein expression in normal tissues and lung adenocarcinoma from THPA. (c) Kaplan–Meier analysis indicated that the low expression of ADH5 was significantly correlated with poor overall survival. (d) Western blotting analysis of ADH5 expression in NSCLC cell lines and one normal human bronchial epithelial cell line. (e) Relative mRNA expression of ADH5 in lung cancer cell lines. ADH5 expression was examined using qRT-PCR and was normalized to GAPDH expression. (f) Western blotting analysis of ADH5 expression in NSCLC tissues compared with corresponding adjacent tissues. (g) Relative expression levels of ADH5 in NSCLC tissues compared with corresponding adjacent tissues. Data from cells were generated from at least three biologically independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
3.2 ADH5 overexpression impaired proliferation but enhanced migration and invasion of NSCLC cells
To investigate the function of ADH5 in NSCLC, we performed overexpression of ADH5 in two different NSCLC cell lines (H358 and PC9), validating the expression levels via western blot and qRT-PCR (Figure 2a). Overexpression of ADH5 significantly suppressed the proliferation and colony-formation ability of both H358 and PC9 cells (Figure 2b–d). Interestingly, transwell assay showed that overexpression of ADH5 increased cell migration and invasion in both H358 and PC9 cells (Figure 2e and f). Hence, ADH5 overexpression inhibited proliferation but promoted metastasis in NSCLC. The presence of this phenotype is a typical characteristic of cancer cell plasticity [22,23], indicating that ADH5 plays a role in NSCLC cell plasticity. Subsequently, PC9 cells transfected with either the ADH5 overexpression vector or an empty vector were then subcutaneously injected into male BALB/c nude mice. The group with ADH5 overexpression exhibited a notable decrease in tumor volumes and weights compared to the control group (Figure 2g and h).
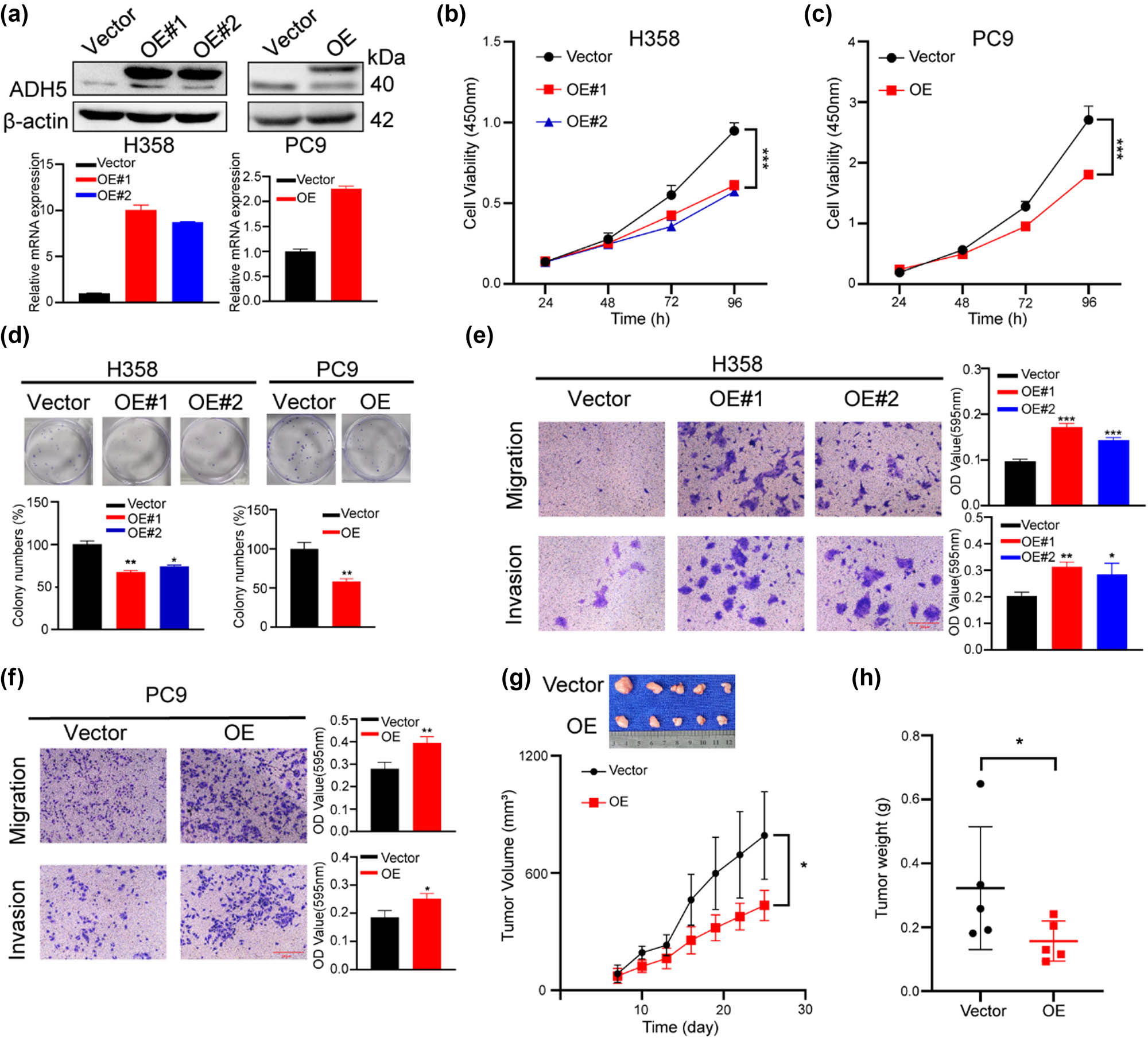
ADH5 overexpression impaired proliferation but enhanced metastasis of NSCLC cells. (a) Western blot and qRT-PCR showed that ADH5 was efficiently overexpressed in H358 and PC9 cells. (b) and (c) Cell-viability assay was performed to measure the growth of ADH5-transfected H358 and PC9 cells. (d) Colony formation was conducted in H358 and PC9 cells transfected with or without ADH5. (e) and (f) Transwell assays were performed to evaluate migration and invasion of ADH5 overexpressed H358 cells and PC9 cells. For migration H358 was incubated for 24 h, PC9 was incubated for 24 h. For invasion H358 was incubated for 48 h, PC9 was incubated for 36 h. (g) and (h) PC9 ADH5-overexpressed cells were subcutaneously injected into BALB/c nude mice. Tumor volumes were evaluated every 3 days. Tumor weights were evaluated 4 weeks later. Data from cells were generated from at least three biologically independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
3.3 ADH5 knockdown impaired migration and invasion but enhanced proliferation of NSCLC cells
To further assess the impact of reduced ADH5 expression in NSCLC cells, shRNAs targeting ADH5 mRNA were employed to silence the expression of ADH5 in H358 and PC9 cells, which were confirmed with western blot and qPCR (Figure 3a). Consistently, downregulation of ADH5 increased the proliferation and colony-forming capacity but hindered the migration of H358 and PC9 cells (Figure 3b–f).
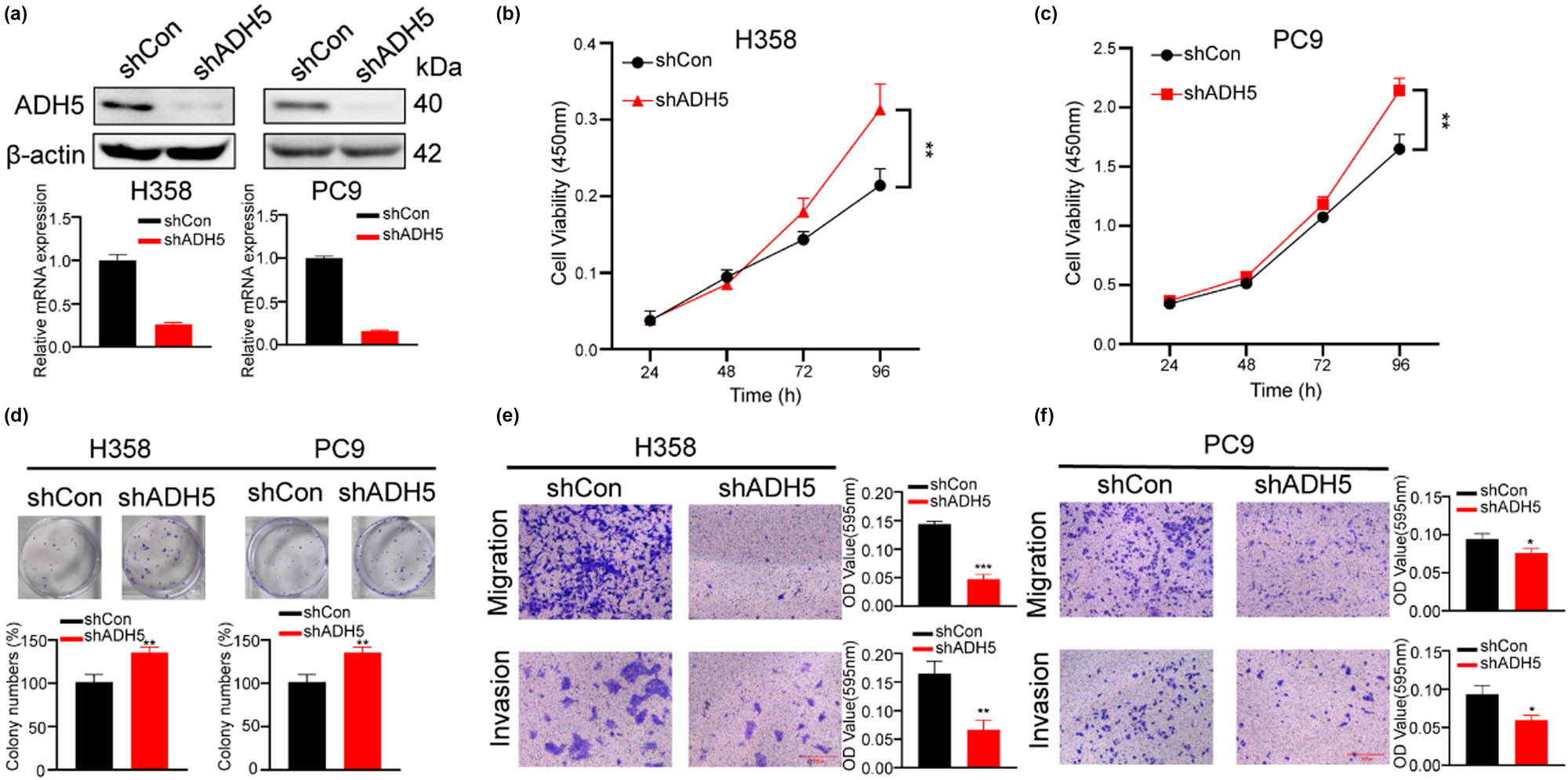
ADH5 knockdown impaired migration but enhanced proliferation of NSCLC cells. (a) Western blot and qRT-PCR showed that ADH5 was efficiently knocked down in H358 and PC9 cells. (b) and (c) CCK8 assay was conducted to measure the proliferation of H358 and PC9 shADH5 cells. The absorbance value was measured at 450 nm. (d) Colony formation was conducted in H358 and PC9 cells lacking ADH5. (e) and (f) Transwell assays were performed to evaluate migration and invasion of H358 and PC9 shADH5 cells. For migration H358 was incubated for 48 h, PC9 was incubated for 24 h. For invasion H358 was incubated for 48 h, PC9 was incubated for 36 h. Data from cells were generated from at least three biologically independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
The advancement of cancer is determined by two key factors: proliferation and metastasis. However, ADH5 plays a “negative” role in cancer proliferation but a “positive” impact on cancer cell migration and invasion which were called metastasis. The discordance in the effects of ADH5 suggests that NSCLC cells are undergoing selective differentiation, emphasizing enhancing metastatic potential while potentially compromising proliferation. This study uncovered the critical involvement of ADH5 in the plasticity of NSCLC cells.
3.4 ADH5 activated Smad2/Smad3
TGF-β plays a crucial role in regulating cancer cell plasticity, stem cell activity, and tumor progression [24–26]. To clarify whether ADH5 modulates the plasticity of NSCLC cell through the regulation of the Smad2/3 pathway, we detected the phosphorylation levels of Smad2/Smad3 in ADH5-overexpressing stable transfected cells. Our results demonstrated that Smad2/Smad3 activation occurred in cells overexpressing ADH5, while it was inhibited in ADH5 knockdown cells (Figure 4a). AKT phosphorylation, downstream of TGF-β, has been proven to induce cancer cell transdifferentiation, as a regulator of cancer cell plasticity [27,28]. Western blot assays showed that ADH5 overexpression activated AKT while ADH5 knockdown inhibited AKT phosphorylation (Figure 4b). Moreover, ADH5 overexpression suppressed the expression of c-Myc, an important factor for cell proliferation, and upregulated P15, a repressor for cell cycle (Figure 4c and d).
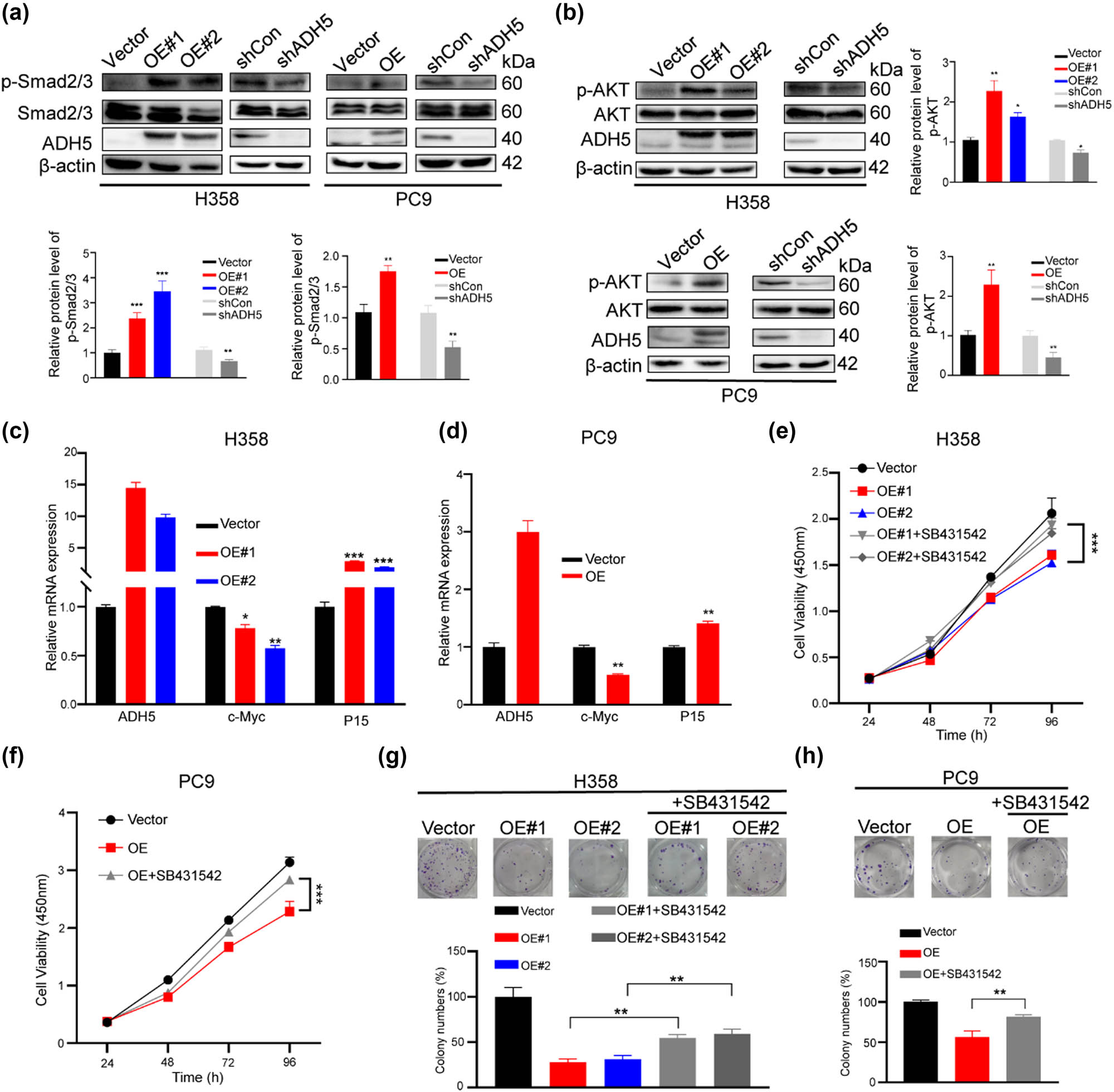
ADH5 activated Smad2/Smad3. (a) Western blot to assess the phosphorylation of Smad2/Smad3 in ADH5 stable transfected H358 and PC9 cells. (b) Western blot to assess the protein expression of ADH5, AKT, and p-AKT in ADH5 transfected stable cell lines. (c) and (d) qRT-PCR was conducted to measure the mRNA levels of c-Myc and P15 in ADH5 stable transfected H358 and PC9 cells. (e) and (f) SB431542 (10 μM) was applied to ADH5 overexpressed cells and cell proliferation was detected by CCK8. (g) and (h) Colony formation was conducted in ADH5 overexpressed cells treated with/without SB431542 (5 μM). Data from cells were generated from at least three biologically independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
To further validate the modulation of Smad2/3 by ADH5 in NSCLC, TGFbetaR1/ALK5 inhibitor SB431542 was applied. When treated with SB431542, the suppressed proliferation by ADH5 was rescued in H358 and PC9 cells (Figure 4e and f), as well as the colony-formation ability (Figure 4g and h). Taken together, the low expression of ADH5 in NSCLC contributes to the progression of cancer cells by impeding the phosphorylation Smad2/3.
3.5 ADH5 upregulated SOX9 and induced EMT
SOX9 regulates lung cancer cell plasticity, promoting invasion and inhibiting proliferation [29]. Given the analogous role of ADH5 to SOX9 in NSCLC, we explored the crosstalk between ADH5 and SOX9. Analysis showed a positive correlation between ADH5 and SOX9 in NSCLC using the GEPIA database (Figure 5a). Moreover, the overexpression of ADH5 led to an elevation in the SOX9 protein expression, whereas knockdown of ADH5 resulted in a reduction in SOX9 expression (Figure 5b). We then discovered that EMT markers like N-cadherin and Snail showed increased levels with ADH5 overexpression and decreased levels with ADH5 knockdown (Figure 5c and d), which is consistent with the observed effects on cell migration and invasion (Figures 2e, f and 3e, f).
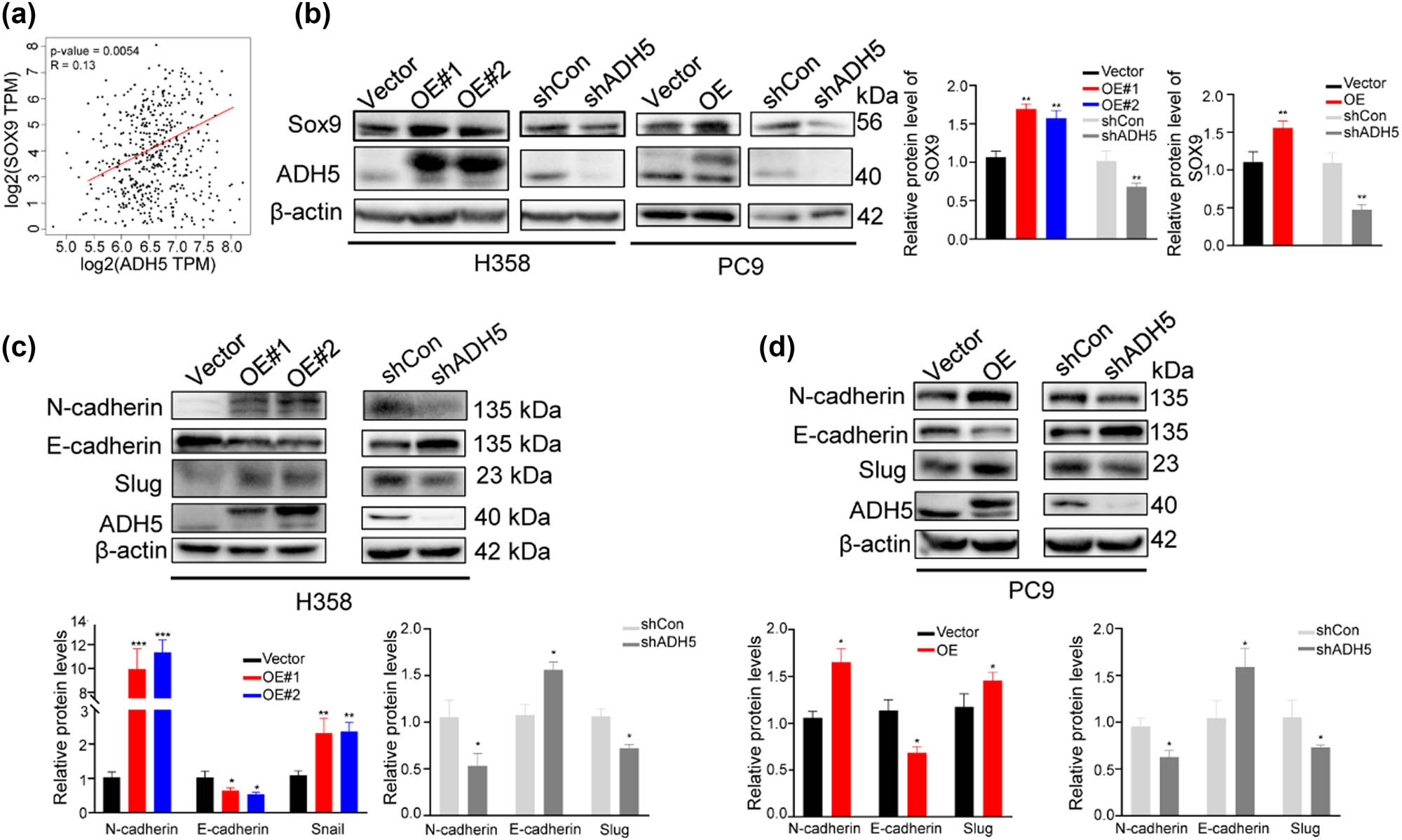
Correlation between ADH5 and SOX9. (a) Scatter plots of correlation between SOX9 and ADH5 expression in lung adenocarcinoma from GEPIA database. (b) Protein level of SOX9 detected by western blot in ADH5 transfected stable cell lines. (c) and (d) Western blot to assess the protein expression of N-cadherin, E-cadherin, Snail, Slug, and ADH5 in ADH5 transfected stable cell lines. Data from cells were generated from at least three biologically independent experiments. *P < 0.05, **P < 0.01, ***P < 0.001.
The findings indicated that ADH5 was involved in activating Smad2/Smad3 and SOX9, thereby ultimately promoting EMT. Furthermore, ADH5 contributed to the suppression of c-Myc, leading to the inhibition of cell proliferation. Consequently, ADH5 regulates the plasticity of NSCLC cells by impeding cancer cell growth while facilitating cancer cell metastasis.
4 Discussion
ADH5 has been linked to various histological and pathological processes. As a regulator of protein S-nitrosylation, ADH5 regulates the bioactivity of nitric oxide [2], which is known for its dual roles in tumorigenesis [30]. Depending on its concentration and spatiotemporal profile, nitric oxide exerts diverse effects on several mechanisms within the tumor microenvironment, such as metabolism, cell cycle, DNA repair, angiogenesis, and apoptosis/necrosis [31]. Mice deficient in ADH5 exhibited increased susceptibility to both spontaneous and carcinogen-induced hepatocellular carcinoma due to the S-nitrosylation of DNA repair enzyme O6-alkylguanine-DNA alkyltransferase [10]. ADH5 deficiency induced S-nitrosylation of focal adhesion kinase 1 and sustained tumorigenicity by providing cancer cells with the ability to evade anoikis [8]. It is evident that ADH5 protects against tumor progression. However, S-nitrosylation involves abundant proteins and the overall effect of ADH5 on tumor requires further investigation. In this study, it was observed that ADH5 has a dual role in NSCLC and mainly exerts its function through the Smad2/Smad3 pathway. Overexpression of ADH5 in H358 and PC9 cells hindered proliferation while prompting migration and invasion, which is a typical characteristic of the TGF-β signaling pathway [32]. The overexpression of ADH5 led to the activation of Smad2/Smad3 and EMT in the cells. And SB431542 successfully counteracted the consequence of ADH5 overexpression in NSCLC cells. It was also observed that SOX9 expression shows a strong positive correlation with ADH5 expression in NSCLC cells. These results suggest that ADH5 induces low proliferation and high invasiveness in NSCLC cells through the activation of Smad2/Smad3 and the modulation of SOX9, a key regulator of NSCLC cell plasticity (Figure 6).
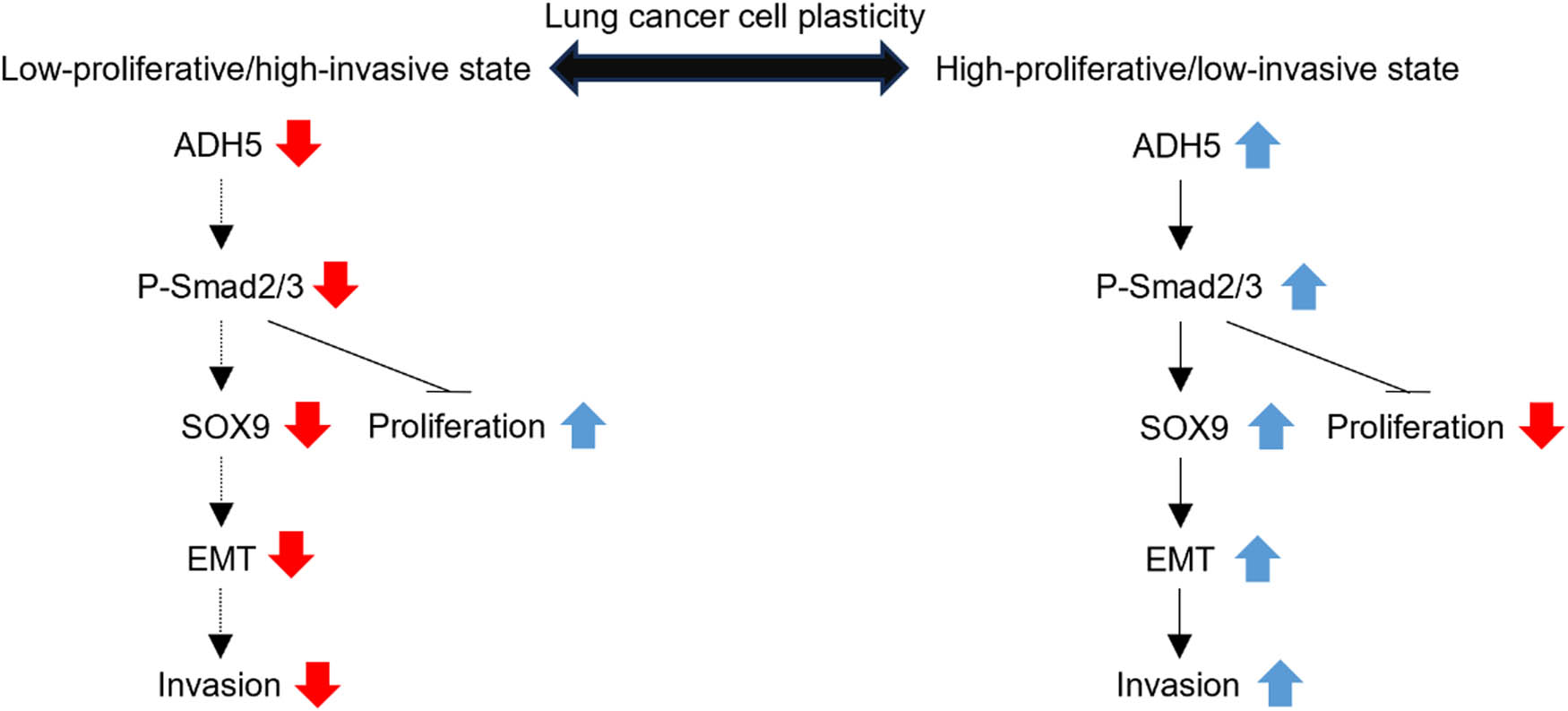
A schematic illustration depicting the regulation of lung cancer cell plasticity by ADH5, where the expression of ADH5 influences the state of lung cancer cells; downregulation of ADH5 suppresses migration but boosts proliferation via Smad2/3, and conversely.
The dual role of TGF-β has been implicated in the progression of many types of cancer, inhibiting tumorigenic inflammation during the early stages of carcinogenesis while promoting various phases of the metastatic process, from invasion, dissemination, immune evasive dormancy, and organ-specific colonization [33,34]. Our research uncovered that ADH5 activated Smad2/Smad3, inducing EMT while suppressing c-Myc, thereby enhancing the migratory and invasive abilities of cells and inhibiting proliferation of NSCLC cells. SOX9, a critical regulator of lung cancer cell plasticity, is upregulated by TGF-β both in embryonic development and tumor progression [24,25]. The control of lung cancer cell plasticity involves a balance between SOX2 and SOX9. SOX2 triggers the EpCAM expression and facilitates proliferation, whereas SOX9 drives SLUG transcription and boosts migration [29]. Our findings demonstrated the positive correlation between ADH5 and SOX9, suggesting that ADH5 plays a role in regulating the plasticity of NSCLC cell.
In conclusion, this study identifies ADH5 as a key regulator of NSCLC cell plasticity through its activation of Smad2/Smad3 and upregulation of SOX9. ADH5 shows promise as a potential biomarker for monitoring the progression of NSCLC, and targeting ADH5 expression could offer a novel therapeutic strategy for treating cancer.
-
Funding information: This work was supported by the Innovative and Entrepreneurial Youth Talents Project of Jiangxi (No. JXSQ2018106040) and Li Ka Shing Foundation LKSF (No. 1210669001).
-
Author contributions: All the authors contributed to the design and implementation of this study. T.X. and H.Y. performed the experiments and acquired the data; H.Y., L.X., X.F., and X.X. performed the database analyses and supervised the study; T.X. and X.X. analyzed the data and wrote the manuscript; L.X. and X.X. supported the research. All the authors reviewed and commented on the final manuscript.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global Cancer Statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49.10.3322/caac.21660Search in Google Scholar
[2] Barnett SD, Buxton ILO. The role of S-nitrosoglutathione reductase (GSNOR) in human disease and therapy. Crit Rev Biochem Mol Biol. 2017;52(3):340–54.10.1080/10409238.2017.1304353Search in Google Scholar
[3] Bentz BG, Simmons RL, Haines GK 3rd, Radosevich JA. The yin and yang of nitric oxide: reflections on the physiology and pathophysiology of NO. Head Neck. 2000;22(1):71–83.10.1002/(SICI)1097-0347(200001)22:1<71::AID-HED11>3.0.CO;2-GSearch in Google Scholar
[4] Sebag SC, Zhang Z, Qian Q, Li M, Zhu Z, Harata M, et al. ADH5-mediated NO bioactivity maintains metabolic homeostasis in brown adipose tissue. Cell Rep. 2021;37(7):110003.10.1016/j.celrep.2021.110003Search in Google Scholar
[5] Koivusalo M, Baumann M, Uotila L. Evidence for the identity of glutathione-dependent formaldehyde dehydrogenase and class III alcohol dehydrogenase. FEBS Lett. 1989;257(1):105–9.10.1016/0014-5793(89)81797-1Search in Google Scholar
[6] Oka Y, Hamada M, Nakazawa Y, Muramatsu H, Okuno Y, Higasa K, et al. Digenic mutations in ALDH2 and ADH5 impair formaldehyde clearance and cause a multisystem disorder, AMeD syndrome. Sci Adv. 2020;6(51):eabd7197.10.1126/sciadv.abd7197Search in Google Scholar
[7] Mulderrig L, Garaycoechea JI, Tuong ZK, Millington CL, Dingler FA, Ferdinand JR, et al. Aldehyde-driven transcriptional stress triggers an anorexic DNA damage response. Nature. 2021;600(7887):158–63.10.1038/s41586-021-04133-7Search in Google Scholar
[8] Rizza S, Di Leo L, Pecorari C, Giglio P, Faienza F, Montagna C, et al. GSNOR deficiency promotes tumor growth via FAK1 S-nitrosylation. Cell Rep. 2023;42(1):111997.10.1016/j.celrep.2023.111997Search in Google Scholar
[9] Rizza S, Filomeni G. Denitrosylate and live longer: how ADH5/GSNOR links mitophagy to aging. Autophagy. 2018;14(7):1285–7.10.1080/15548627.2018.1475818Search in Google Scholar
[10] Wei W, Li B, Hanes MA, Kakar S, Chen X, Liu L. S-nitrosylation from GSNOR deficiency impairs DNA repair and promotes hepatocarcinogenesis. Sci Transl Med. 2010;2(19):19ra3.10.1126/scitranslmed.3000328Search in Google Scholar
[11] Zhang Y, Wu K, Su W, Zhang DF, Wang P, Qiao X, et al. Increased GSNOR expression during aging impairs cognitive function and decreases S-nitrosation of CaMKIIα. J Neurosci. 2017;37(40):9741–58.10.1523/JNEUROSCI.0681-17.2017Search in Google Scholar PubMed PubMed Central
[12] Tang X, Zhao S, Liu J, Liu X, Sha X, Huang C, et al. Mitochondrial GSNOR alleviates cardiac dysfunction via ANT1 denitrosylation. Circ Res. 2023;133(3):220–36.10.1161/CIRCRESAHA.123.322654Search in Google Scholar PubMed
[13] Hayashida K, Bagchi A, Miyazaki Y, Hirai S, Seth D, Silverman MG, et al. Improvement in outcomes after cardiac arrest and resuscitation by inhibition of S-nitrosoglutathione reductase. Circulation. 2019;139(6):815–27.10.1161/CIRCULATIONAHA.117.032488Search in Google Scholar PubMed PubMed Central
[14] Casin KM, Fallica J, Mackowski N, Veenema RJ, Chan A, St Paul A, et al. S-nitrosoglutathione reductase is essential for protecting the female heart from ischemia-reperfusion injury. Circ Res. 2018;123(11):1232–43.10.1161/CIRCRESAHA.118.313956Search in Google Scholar PubMed PubMed Central
[15] Rizza S, Di Leo L, Mandatori S, De Zio D, Filomeni G. Mitophagy contributes to alpha-tocopheryl succinate toxicity in GSNOR-deficient hepatocellular carcinoma. Biochem Pharmacol. 2020;176:113885.10.1016/j.bcp.2020.113885Search in Google Scholar PubMed
[16] Rizza S, Cardaci S, Montagna C, Di Giacomo G, De Zio D, Bordi M, et al. S-nitrosylation drives cell senescence and aging in mammals by controlling mitochondrial dynamics and mitophagy. Proc Natl Acad Sci USA. 2018;115(15):E3388–97.10.1073/pnas.1722452115Search in Google Scholar PubMed PubMed Central
[17] Rizza S, Montagna C, Cardaci S, Maiani E, Di Giacomo G, Sanchez-Quiles V, et al. S-nitrosylation of the mitochondrial chaperone TRAP1 sensitizes hepatocellular carcinoma cells to inhibitors of succinate dehydrogenase. Cancer Res. 2016;76(14):4170–82.10.1158/0008-5472.CAN-15-2637Search in Google Scholar PubMed
[18] Rizza S, Filomeni G. Tumor suppressor roles of the denitrosylase GSNOR. Crit Rev Oncog. 2016;21(5–6):433–45.10.1615/CritRevOncog.2017021074Search in Google Scholar PubMed
[19] Cañas A, López-Sánchez LM, Peñarando J, Valverde A, Conde F, Hernández V, et al. Altered S-nitrosothiol homeostasis provides a survival advantage to breast cancer cells in HER2 tumors and reduces their sensitivity to trastuzumab. Biochim Biophys Acta. 2016;1862(4):601–10.10.1016/j.bbadis.2016.02.005Search in Google Scholar PubMed
[20] Mena-Osuna R, Mantrana A, Guil-Luna S, Sánchez-Montero MT, Navarrete-Sirvent C, Morales-Ruiz T, et al. Metabolic shift underlies tumor progression and immune evasion in S-nitrosoglutathione reductase-deficient cancer. J Pathol. 2023;260(3):261–75.10.1002/path.6080Search in Google Scholar PubMed
[21] Cirotti C, Rizza S, Giglio P, Poerio N, Allega MF, Claps G, et al. Redox activation of ATM enhances GSNOR translation to sustain mitophagy and tolerance to oxidative stress. EMBO Rep. 2021;22(1):e50500.10.15252/embr.202050500Search in Google Scholar PubMed PubMed Central
[22] Miao CC, Hwang W, Chu LY, Yang LH, Ha CT, Chen PY, et al. LC3A-mediated autophagy regulates lung cancer cell plasticity. Autophagy. 2022;18(4):921–34.10.1080/15548627.2021.1964224Search in Google Scholar PubMed PubMed Central
[23] Cheng W, Li HL, Xi SY, Zhang XF, Zhu Y, Le X, et al. Growth differentiation factor 1-induced tumour plasticity provides a therapeutic window for immunotherapy in hepatocellular carcinoma. Nat Commun. 2021;12(1):7142.10.1038/s41467-021-27525-9Search in Google Scholar PubMed PubMed Central
[24] Li T, Huang H, Shi G, Zhao L, Li T, Zhang Z, et al. TGF-β1-SOX9 axis-inducible COL10A1 promotes invasion and metastasis in gastric cancer via epithelial-to-mesenchymal transition. Cell Death Dis. 2018;9(9):849.10.1038/s41419-018-0877-2Search in Google Scholar PubMed PubMed Central
[25] Yasen A, Feng J, Xie XM, Li K, Cai YH, Liao ZH, et al. Exosomes derived from TGF-β1-pretreated mesenchymal stem cells alleviate biliary ischemia-reperfusion injury through Jagged1/Notch1/SOX9 pathway. Int Immunopharmacol. 2023;119:110253.10.1016/j.intimp.2023.110253Search in Google Scholar PubMed
[26] Furumatsu T, Ozaki T, Asahara H. Smad3 activates the Sox9-dependent transcription on chromatin. Int J Biochem Cell Biol. 2009;41(5):1198–204.10.1016/j.biocel.2008.10.032Search in Google Scholar PubMed PubMed Central
[27] Quintanal-Villalonga A, Taniguchi H, Zhan YA, Hasan MM, Chavan SS, Meng F, et al. Comprehensive molecular characterization of lung tumors implicates AKT and MYC signaling in adenocarcinoma to squamous cell transdifferentiation. J Hematol Oncol. 2021;14(1):170.10.1186/s13045-021-01186-zSearch in Google Scholar PubMed PubMed Central
[28] Elghazi L, Weiss AJ, Barker DJ, Callaghan J, Staloch L, Sandgren EP, et al. Regulation of pancreas plasticity and malignant transformation by Akt signaling. Gastroenterology. 2009;136(3):1091–103.10.1053/j.gastro.2008.11.043Search in Google Scholar PubMed PubMed Central
[29] Lin SC, Chou YT, Jiang SS, Chang JL, Chung CH, Kao YR, et al. Epigenetic switch between SOX2 and SOX9 regulates cancer cell plasticity. Cancer Res. 2016;76(23):7036–48.10.1158/0008-5472.CAN-15-3178Search in Google Scholar PubMed
[30] Xu W, Liu LZ, Loizidou M, Ahmed M, Charles IG. The role of nitric oxide in cancer. Cell Res. 2002;12(5–6):311–20.10.1038/sj.cr.7290133Search in Google Scholar PubMed
[31] Panneerselvan P, Vasanthakumar K, Muthuswamy K, Krishnan V, Subramaniam S. Insights on the functional dualism of nitric oxide in the hallmarks of cancer. Biochim Biophys Acta Rev Cancer. 2023;1878(6):189001.10.1016/j.bbcan.2023.189001Search in Google Scholar PubMed
[32] Pickup M, Novitskiy S, Moses HL. The roles of TGFβ in the tumour microenvironment. Nat Rev Cancer. 2013;13(11):788–99.10.1038/nrc3603Search in Google Scholar PubMed PubMed Central
[33] Rastogi S, Mishra SS, Arora MK, Kaithwas G, Banerjee S, Ravichandiran V, et al. Lactate acidosis and simultaneous recruitment of TGF-β leads to alter plasticity of hypoxic cancer cells in tumor microenvironment. Pharmacol Ther. 2023;250:108519.10.1016/j.pharmthera.2023.108519Search in Google Scholar PubMed
[34] Massagué J, Sheppard D. TGF-β signaling in health and disease. Cell. 2023;186(19):4007–37.10.1016/j.cell.2023.07.036Search in Google Scholar PubMed PubMed Central
© 2025 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- 10.1515/biol-2025-1168
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”
Articles in the same Issue
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- 10.1515/biol-2025-1168
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”