Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
-
Xinlu Bai
 ,
Yaping Li
,
Yaping Li

Abstract
Atmospheric nitrogen (N) deposition is recognized as a pivotal nutrient input in forest ecosystems. However, significant gaps persist in our comprehension of the global-scale fate of N in forest ecosystems. In a pioneering effort, this study analyzed the fluxes and determinants of deposited N by 234 observations from 52 published articles. Our findings indicated that plant uptake, soil retention, and N losses, respectively, accounted for 27.4, 57.9, and 14.6% of the total deposited N. The fate of deposited N was significantly influenced by a suite of factors, including forest type, climatic parameters such as mean annual temperature (MAT) and precipitation (MAP), edaphic characteristics such as soil pH and the carbon to nitrogen ratio (C/N), and experimental factors like nitrogen addition rate (NR), nitrogen forms (NF), plot size (PS) for 15N studies, and the duration of study. For the uptake of deposited N, MAP emerged as the predominant positive factors, whereas NR was the dominant negative factors; for deposited N soil retention, NR was the key positive factors, while MAT was the key negative factors; for N losses, MAP was the predominant positive factors, with the C/N ratio serving as a significant negative factor. Thus, for a given forest ecosystem with relatively stable climate and soil conditions, NR, NF, and the soil C/N were the main controlling factors regulating the fate of deposited N. These insights significantly advance our grasp of the N cycle in forest ecosystems. Consecutive monitoring of the impact of deposited N on soil N transformations and carbon sequestration is needed in future studies.
1 Introduction
Human activities, such as fossil fuel consumption, animal husbandry expansion, and agricultural intensification, have strongly accelerated both reactive nitrogen (N) emissions and their deposition in the terrestrial environment globally [1,2]. By the end of the twentieth century, the rate of nitrogen deposition had at least doubled [3], with a substantial amount destined for terrestrial ecosystems. Increased N deposition can alter N cycles, affect ecosystem functions [4,5], and increase N losses via leaching and denitrification, especially in forest ecosystems, because of its limited N availability [6,7,8]. Critically, the effects of N deposition on forest ecosystems depends on the fate of deposited N, including uptake by plants [9,10], retention in the soil [11], and losses [12]. Distinguishing the fate of the deposited N is crucial to understand the effects of N deposition on forest ecosystems better. Many studies have been conducted to assess the deposited N flux by 15N addition experiments at the field scale [13,14]. Xie et al. [15] and Leeson et al. [16] found that N deposition significantly increased ammonia (NH3) volatilization, nitrous oxide (N2O) emissions, and nitrate leaching in forest ecosystems. The accumulation of N in the soil and plant bodies increases with the N deposition [17,18,19]. In addition, determining the fate of deposited N is also essential for an in-depth study of carbon sequestration in forest systems, as the relationship between carbon and nitrogen is very close [20,21,22]. Therefore, it is essential to determine the fate of deposited N in forest systems on a global scale. However, identifying the distribution of deposited N on a global scale remains challenging, due to the challenges of complexity, laboriousness, and the expensive cost of the 15N addition experiments method [23,24].
The fate of deposited N can be influenced by various factors, including forest type (FT), climate (e.g., mean annual air temperature and precipitation), forms of N input (e.g.,
In an effort to bridge the existing gaps in knowledge, this study is designed to quantify the dynamics and fate of N deposits in forest ecosystems. The objectives of the present study are (1) to determine the fate of deposited N in forest ecosystems on a global scale by meta-analysis based on 15N tracer studies and (2) to determine the controlling factors influencing deposited N fate in forest ecosystems.
2 Materials and methods
2.1 Data collection and dataset overview
The dataset was generated by collecting 234 observations from 52 papers containing data from 15N tracer studies across various forest ecosystems. We used the following terms: “the fate of deposited N”; “fates of atmospheric deposited N”; “different fates of deposited N”; “retention of deposited N”; “pathways and dynamics of deposited N” and “recovery of deposited N” to search for publications in Google Scholar and the Web of Science Database up to 31 March, 2024. We used the following criteria for collecting data: (1) conducted the study via field experiments rather than soil column and pot experiments and (2) measured N fluxes using the 15N tracer method. Data were obtained from tables and figures. If the data were presented only in figures, they were digitized using Get-Data v.2.22 (http://gatdata-graph-digitizer.com). The site-specific details were assembled to analyze the patterns and controlling factors of the fate of the deposited N. The experimental location, FT, MAT, mean annual precipitation (MAP), N addition rate (NR), N forms (NF), soil pH, soil carbon to nitrogen ratio (C/N), 15N plot size (PS), and study duration time (SDT) were assembled. We matched these environmental factors with 15N uptake, 15N retention, and 15N losses (Table 1). The experimental locations are shown in Figure 1.
Characteristics of the climatic variables (MAT, MAP), soil properties (pH, C/N), and study variables (PS, SDT, and NR) from the peer-reviewed articles
| Terms | MAT (°C) | MAP (mm) | pH | C/N | PS (m2) | SDT (year) | NR (kg N ha−1) |
|---|---|---|---|---|---|---|---|
| Range | −5.4 to 25.6 | 365–5,500 | 3.0–8.0 | 11.6–64.0 | 0.03–3,000 | 0.04–23 | 0.03–640 |
| Mean value | 8.4 | 1,405 | 4.6 | 24.3 | 136 | 2.8 | 49.1 |
| SD | 5.1 | 1,003 | 1.3 | 10.6 | 125 | 3.2 | 32.9 |
| n | 87 | 92 | 73 | 67 | 121 | 102 | 202 |
MAT, mean annual temperature; MAP, mean annual precipitation; C/N, the ratio of soil carbon to nitrogen; PS, 15N plot size; SDT, study duration time; NR, nitrogen rate; NF, nitrogen forms. “SD” and “n” represent the standard deviation and number of all observations.
Global distribution of study sites included in this work.
2.2 Data analyses
Prior to analysis, all data were checked for normality using the Shapiro–Wilk test. The 95% confidence intervals (CIs) were generated using bootstrapping (4,999 iterations). We calculated the relative importance of the explanatory variables, defined as the magnitude of the increase in the mean squared errors of the multiple linear regression models when one of the selected explanatory variables was randomly permuted, to determine the controlling factors influencing the deposited N fates [29]. Multiple linear regression models were also used [30]: Y = β 0 + β 1 × X 1 + β 2 × X 2 + β 3 × X 3 + β 4 × X 4 + β 5 × X 5 + β 6 × X 6 + β 7 × X 7 + β 8 × X 8 + ε, where β 0, β, and ε are the intercept, slope value, and sampling error, respectively; and X 1, X 2, X 3, X 4, X 5, X 6, X 7, and X 8 refer to MAT, MAP, NR, NF, soil pH, soil carbon to nitrogen ratio, 15N PS, and duration time of the study, respectively. The relative importance of the explanatory variables was obtained using relative weights via the “Car” and “Mass” packages R software [31]. Specifically, this method quantifies the independent contribution of each explanatory variable to the response variable by decomposing the model’s coefficient of determination (R 2). This approach circumvents the bias induced by multicollinearity among predictor variables, which is a common issue in traditional regression analysis. Relative weight analysis provides a more precise reflection of each variable’s importance ranking within the overall model, particularly in scenarios involving highly correlated predictors. The multiple linear regression models and associated predictor variables are presented in Table 2.
Models of multiple linear regression on the fate of deposited nitrogen and associated predictor variables
| Fate | Predictor variables | Estimate | Standard error | p value | Significance level | R 2 | Adjusted R 2 |
|---|---|---|---|---|---|---|---|
| Uptake | MAT | −0.57 | 0.27 | p < 0.01 | ** | 0.603 | 0.581 |
| MAP | 0.01 | 0.01 | 0.03 | * | |||
| PS | 0.01 | 0.01 | 0.13 | ||||
| SDT | 1.01 | 0.31 | 0.21 | ||||
| pH | −0.24 | 0.04 | 0.16 | ||||
| C/N | 0.15 | 0.05 | 0.04 | * | |||
| NR | −0.31 | 0.11 | 0.02 | * | |||
| NF | 0.11 | 0.03 | 0.12 | ||||
| Intercept | 15.74 | 7.84 | 0.04 | * | |||
| Retention | MAT | −0.97 | 0.33 | p < 0.01 | ** | 0.565 | 0.463 |
| MAP | 0.01 | 0.01 | 0.14 | ||||
| PS | 0.01 | 0.01 | 0.15 | ||||
| SDT | −0.16 | 0.06 | 0.23 | ||||
| pH | 2.39 | 0.84 | 0.03 | * | |||
| C/N | 0.05 | 0.01 | 0.03 | * | |||
| NR | 0.03 | 0.01 | 0.02 | * | |||
| NF | −0.09 | 0.02 | p < 0.01 | ** | |||
| Intercept | 66.45 | 8.78 | p < 0.01 | ** | |||
| Losses | MAT | 2.01 | 0.61 | p < 0.01 | ** | 0.732 | 0.654 |
| MAP | 0.03 | 0.01 | 0.12 | ||||
| PS | 0.03 | 0.01 | 0.11 | ||||
| SDT | 0.28 | 0.23 | 0.12 | ||||
| pH | 0.63 | 0.47 | 0.82 | ||||
| C/N | 0.30 | 0.12 | 0.02 | * | |||
| NR | 0.03 | 0.01 | 0.02 | * | |||
| NF | 0.30 | 0.21 | 0.55 | ||||
| Intercept | −7.51 | 2.32 | 0.02 | * |
MAT, mean annual temperature; MAP, mean annual precipitation; C/N, the ratio of soil carbon to nitrogen; PS, 15N plot size; SDT, study duration time; NR, nitrogen rate; NF, nitrogen forms.
3 Results
3.1 Nitrogen uptake
Total N uptake accounted for 27.4% of the deposited N, and the roots, stems, branches, foliage, and understory accounted for 6.0, 3.1, 3.6, 6.3, and 8.5% of the total N uptake, respectively (Figure 2). FT strongly influenced N assimilation, with tropical/subtropical evergreen forests (TSFs) exhibiting the highest uptake capacity, followed by temperate broad-leaved (TBF) and coniferous forests (TCF). Moreover, it had higher N uptake with warm temperatures, low precipitation, and low soil pH and C/N. Warmer temperatures and intermediate-to-high precipitation enhanced N uptake. The amounts of N uptake decreased as follows: 31.6% (MAT > 10°C), 26.3% (6°C ≤ MAT ≤ 10°C), 24.0% (MAT < 6°C); 23.9% (MAP ≤ 1,000 mm), 29.7% (1,000 mm < MAP < 1,500 mm), 32.4% (MAP ≥ 1,500 mm); 34.3% (pH ≤ 4), 28.7% (4 < pH ≤ 6), 24.9% (pH > 6); and 34.2% (C/N ≤ 20), 26.7% (20 < C/N ≤ 30), and 22.1% (C/N > 30).
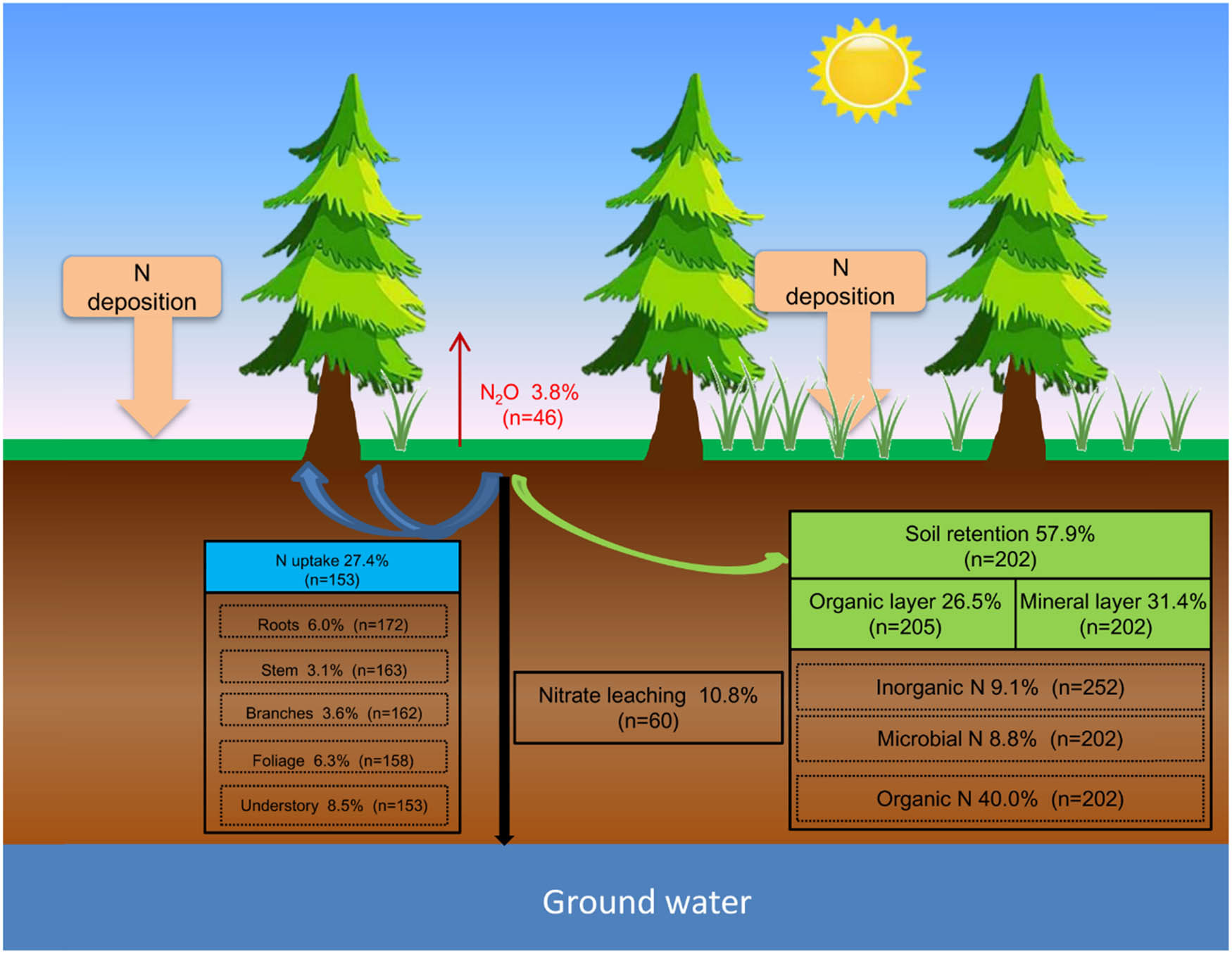
Fate of the deposited N in forest ecosystems. The numbers in the parentheses are the numbers of observations.
In addition to FT, climate, and soil properties, the characteristics of deposited N and experimental duration also contributed to the differences in N uptake. Compared with the low NR (NR ≤ 50 kg N ha−1), the high NR (NR > 50 kg N ha−1) significantly decreased the total N uptakes by 26.4–38.0%. The total N uptake was 20.4, 30.1, and 26.9% when applied as 15
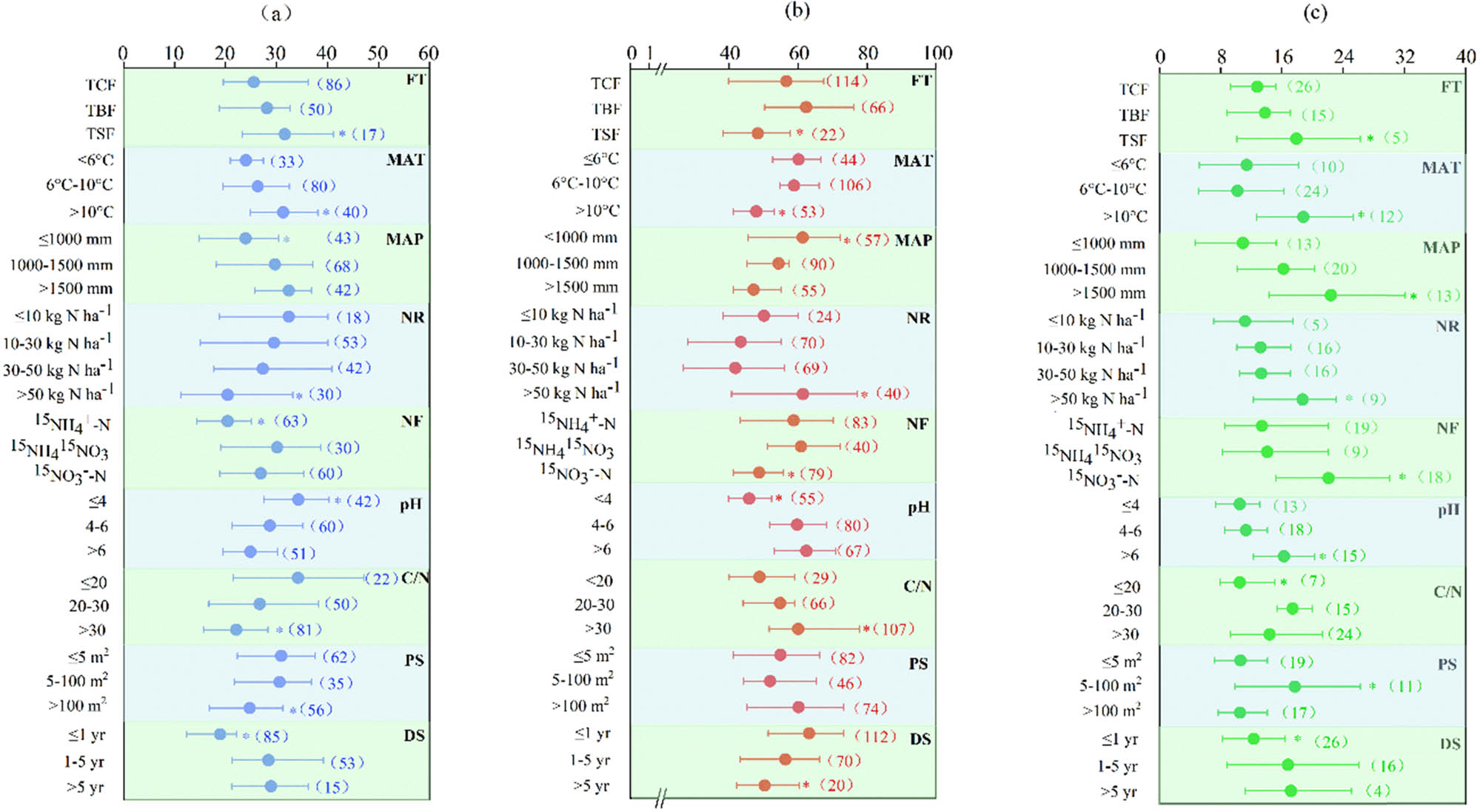
Effect of FT, MAT, MAP, NR, NF, soil pH, soil carbon to nitrogen ratio, 15N PS, and SDT on nitrogen uptake (a), nitrogen retention (b), and nitrogen losses (c), respectively. FT, forest type; TCF, temperate coniferous forest; TBF, temperate broad-leaved forest; TSF, tropic and subtropical evergreen forest; MAT, mean annual temperature; MAP, mean annual precipitation; NR, deposited nitrogen rate; NF, deposited nitrogen forms; C/N, the ratio of soil carbon to nitrogen; PS, 15N plot size; and SDT, study duration time. The numbers in the parentheses are the number of observations. The values are mean values with 95% CIs. *Correlation is significant at p < 0.05.
3.2 Nitrogen retention
N retention in the soil was dominated and accounted for 57.9% of the deposited N, including 26.5% in the organic layer and 31.4% in the mineral layer. Microbial N, inorganic N, and organic N accounted for 8.8, 9.1, and 40.0% of residual N, respectively (Figure 2). The N retention in TCPs and TBFs was 56.6 and 62.3%, respectively, which was significantly higher than that in TSFs (48.3%). The climate also contributed to differences in N retention. The N retention was 58.8–60.1% with MAT ≤ 10°C and 61.3% with MAP ≤ 1,000 mm, which were significantly higher than those of 47.9% MAT > 10°C and 47.1–54.3% with MAP > 1,000 mm, respectively (Figure 3b).
Large amounts of deposited N led to more residual N in the soil. The N retention was 61.4% with NR > 50 kg N ha−1, which was significantly higher than that of 41.8–50.1% with NR < 50 kg N ha−1. For deposited N forms,
3.3 Nitrogen losses
The N loss accounted for 14.6% of the deposited N, including 10.8% for nitrate leaching and 3.8% for nitrous oxide emission, respectively (Figure 2). The total N losses were 12.8 and 13.8% in TCPs and TBFs, respectively, and significantly increased by 29.7–39.8% in TSFs, respectively. Total N losses were also dependent on climate and soil conditions. The highest N losses occurred with MAT > 10°C and MAP > 1,500 mm. Moreover, our meta-analysis showed that soil pH and C/N ratio positively affected total N loss. The total N losses were significantly higher with soil pH > 6 (16.3%), and with soil C/N ≥ 20 (17.4 and 14.4%), respectively.
In addition, the characteristics of deposited N also contributed to the differences in N losses. Total N losses were 9.3, 13.2, 12.3, and 15.7% with deposited N < 10 kg N ha−1, 10–30 kg N ha−1, 30–50 kg N ha−1, and > 50 kg N ha−1, respectively. Total N losses were much more pronounced for 15
3.4 Explanatory factors for the fate of deposited N
Deposited N uptake significantly increased with MAT and MAP, whereas significantly decreased with NR and soil C/N (Figure S1). MAT (38.9% of R 2) and MAP (14.1% of R 2) were the dominant positive factors and the deposited N rate (NR, 15.1% of R 2) and C/N (14.9% of R 2) were the dominant negative factors for deposited N uptake in forest ecosystems. Deposited N retention significantly increased with NR, soil pH, and C/N, whereas significantly decreased with MAT (Figure S2). NR (16.2% of R 2), pH (12.6% of R 2), and C/N (11.6% of R 2) were the dominant positive factors, whereas MAT (32.1% of R 2) and the deposited N form (NF, 20.3% of R 2) were the dominant negative factors for deposited N retention in forest ecosystems. In addition, deposited N losses significantly increased with MAP and NR, whereas significantly decreased with C/N (Figure S3). MAP (51.6% of R 2) and NR (12.3% of R 2) were the dominant positive factors, and C/N (17.4% of R 2) was the dominant negative factor for deposited N losses in forest ecosystems (Figure 4).
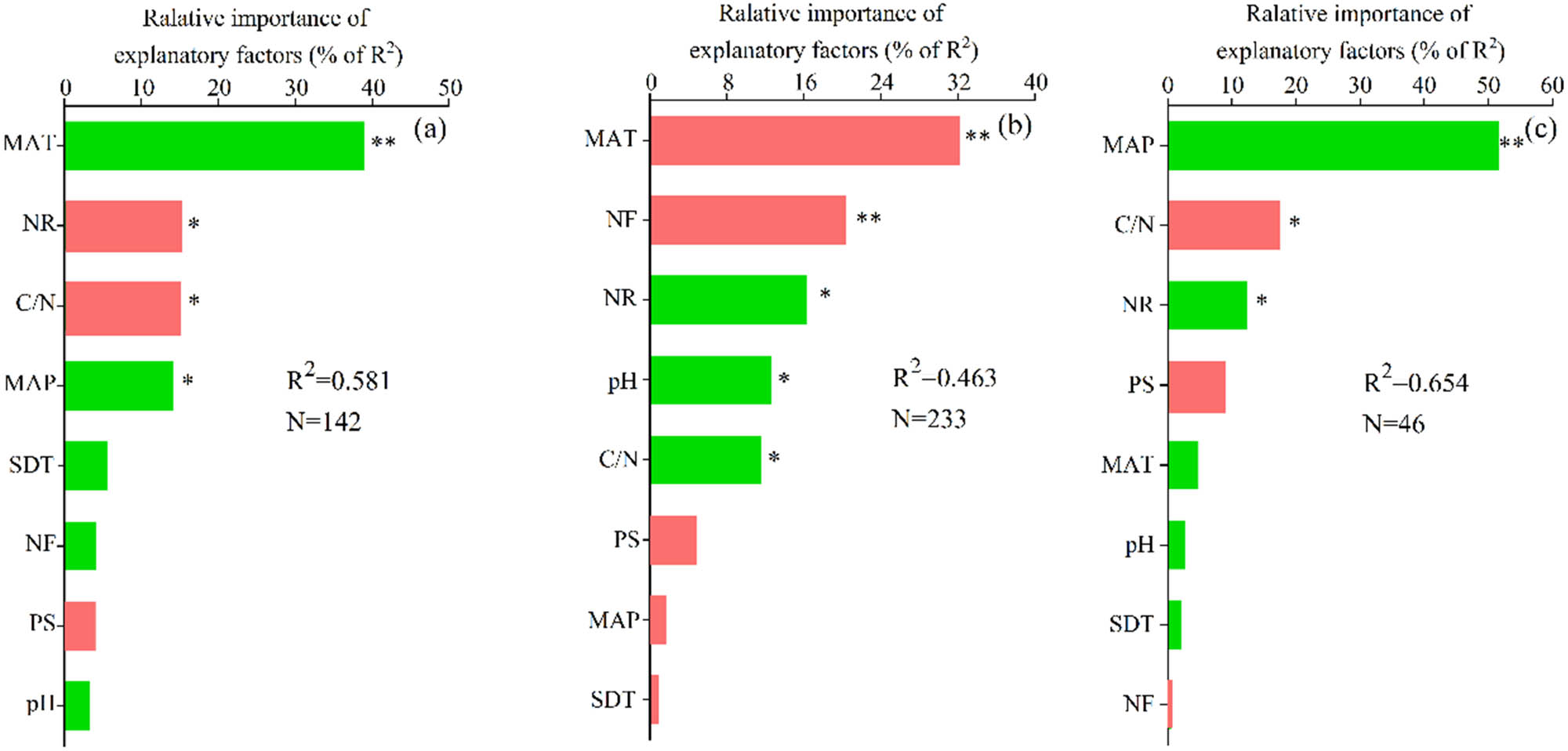
Relative importance of explanatory factors for nitrogen uptake (a), nitrogen retention (b), and nitrogen losses (c) in forest ecosystems. Green columns indicate a positive effect, and red columns indicate a negative effect. ** and *** represent significance at p < 0.05 and p < 0.01, respectively. The predictor variables of multiple linear models are described in Table 1. MAT, mean annual temperature; MAP, mean annual precipitation; NR, nitrogen rate; NF, nitrogen form; C/N, the ratio of soil carbon to nitrogen; PS, 15N plot size; SDT, study duration time.
4 Discussion
4.1 Differential fates of deposited N in forest ecosystems
The total deposited N recoveries for plant and soil pools were 80–88% in forest ecosystems (Figure 3), which is consistent with previous studies. Gurmesa et al. [32] reported that 66–68% of the deposited N was retained in plant and soil pools. Templer et al. [33] reported that more than 70% of the deposited N was retained in plant and soil pools. N retention in the soil dominated and shared 60–69% of the total N recovery. Similarly, a new study by Sun et al. [34] showed that 84% of deposited N was retained in the heavily polluted forests of the Beijing-Tianjin-Hebei region. Considering that forest soil has a high C/N ratio and large amounts of easily available carbon [35], it is a strong sink for N mobilization. In addition, the results revealed that a greater recovery of deposited N was observed in TBF and TCP than in TSF because of higher N retention in soil. These results, indicate that lower N losses were responsible for greater N retention in the soil of TBF and TCP.
Nitrate leaching shared 70–81%, and dominated N losses in forest ecosystems (Figure 3). Similarly, Fang et al. [36] and Liu et al. [37] reported similar nitrate leaching rates in forest ecosystems. Nitrate leaching is also significantly influenced by precipitation [38,39]. Therefore, significantly higher nitrate leaching and total N loss occurred in TSFs. In addition, losses via N2O emissions accounted for only a small proportion of the total deposited N, which is consistent with previous studies. Eickenscheidt et al. [40] reported N2O emission of 0.3–2.6 kg N ha−1 year−1 in beech and spruce forests. In addition, ammonia volatilization and denitrification losses represent major mechanisms for nitrogen loss from deposited N. However, current research on ammonia volatilization and denitrification losses associated with deposited N remains relatively scarce, with incomplete data and unclear mechanisms, making it challenging to effectively incorporate these processes into the analysis. Therefore, we suggest that subsequent research should focus on the dynamic mechanisms, influencing factors, and quantitative analysis of ammonia volatilization and denitrification processes in deposited N. This will enhance our understanding of deposited N loss pathways and provide a scientific basis for relevant environmental management and emission reduction strategies.
4.2 Controls on fates of deposited N in forest ecosystems
The results of our analyses demonstrated that various factors influence the fate of deposited N in forest ecosystems. MAT and MAP were the dominant positive factors (53.1% of R
2) for deposited N uptake. MAT and MAP significantly affect forest plant growth, and suitable temperatures and precipitation increase forest plant biomass and, therefore, deposited N uptake. The results revealed that NR and C/N were the dominant negative factors (30.0% of R
2) for deposited N uptake. Because forest ecosystems are commonly considered N-deficient, more deposited N could be expected to be absorbed with increased deposited N [22,40]. However, lower rates of deposited N were absorbed with a high deposited N load, which was supported by our results showing that deposited N uptake rates significantly decreased with deposited N > 50 kg N ha−1 (Figure 3). Similarly, Templer et al. [33] reported that N uptake rates decrease with deposited N, with an apparent threshold rate of 46 kg N ha−1 year−1. When the deposited N rate in forest ecosystems exceeds 50 kg N ha⁻¹ year⁻¹ (which is far beyond the global critical load threshold of 5–25 kg N ha⁻¹ year⁻¹), the N retention capacity collapses [41]. The core mechanism underlying this phenomenon lies in the imbalance of multi-level biogeochemical processes. The deposited N is predominantly in the form of
4.3 Implications and suggestions
The intergovernmental panel on climate change concluded that climate change was no longer a subject of debate. Climate change manifests itself in many ways, including increased temperatures, changes in precipitation, and precipitation trends. The results of our analyses demonstrated that climate change could significantly affect the fate of deposited N in forest ecosystems. The positive correlation between MAT and deposited N uptake (Figure 4a) was consistent with the result of Templer et al. [33] and Elrys et al. [57]. The pattern of greater deposited N uptake with higher MAT suggested that higher temperatures may have led to higher rates of N uptake by plants [58]. However, greater deposited N uptake could not offset the possible increases in the rates of soil N-cycling processes, such as mineralization and nitrification, and losses via gas emissions or leaching [10]. Therefore, the total ecosystem recovery (N uptake and N retention in the soil) significantly decreased (Figure 3). These results agreed with the watershed mass balances, showing that N exports increase with increasing temperatures [59,60].
Precipitation also controls the fate of deposited N in forest ecosystems. The N retention in the soil was negatively correlated with MAP, whereas the N loss was positively correlated with MAP (Figure 3). These results suggested that N export increased with increasing precipitation. N export from soils may increase soil CO2 efflux into the atmosphere, thereby affecting the global carbon cycle and contributing to global warming [61,62]. Therefore, consecutive monitoring of soil N retention, carbon sequestration, and CO2 emissions is required for future studies.
Global N deposition is likely to increase in the future, increasing N loss (Figure 4). Recently, Tang et al. [63] revealed that increased N deposition stimulated tropical montane forest soil N2 and N2O emissions. Moreover, the increased N deposition causes soil acidification [64], thereby influencing the soil N cycling processes.
The form of deposited N has changed, particularly in China, where the growth rate of deposited nitrate was the highest [65]. Although the total ecosystem N retention was similar for ammonium and nitrate, plants took up more nitrate, and the soil retained more ammonium (Figure 3). This suggested that increased deposited Nitrate could lead to N export from soils, and thereby influencing soil N and C cycles. Furthermore, nitrate is vulnerable to leaching losses, and nitrate leaching deteriorates as more nitrogen is deposited in the form of nitrate. Therefore, the consecutive monitoring of nitrate leaching is required in future studies to better understand the relationships between deposited N and water pollution.
5 Conclusion
Our study provides a comprehensive evaluation of deposited N in forest ecosystems. Our findings revealed that plant uptake, soil retention, and N loss were 27.4, 57.9, and 14.6%, respectively. Deposited N uptake was positively affected by MAT and MAP, but negatively affected by the deposited N rate and C/N. Soil retained deposited N was positively affected by deposited N rate, soil pH, and C/N, but negatively affected by MAT and deposited N form. Deposited N losses were positively affected by MAP and deposited N rate, but negatively affected by C/N. We also found that changes in climate and deposited N could significantly influence soil N export and thereby, soil carbon sequestration. Thus, consecutive monitoring of soil N retention and carbon sequestration dynamics is critically needed, not merely to document biogeochemical fluxes, but to quantify the trade-offs between nitrogen export and ecosystem services.
Acknowledgments
We thank LetPub (www.letpub.com.cn) for its linguistic assistance during the preparation of this manuscript.
-
Funding information: This work was supported by the Natural Science Foundation of Sichuan Province (2023NSFSC1253), the Tarim University President’s Fund “The effect of replacing chemical fertilizers with organic liquid fertilizers on the fate of chemical fertilizers in cotton fields in southern Xinjiang” (TDZKBS202407), and the Tarim University President’s Fund “Innovative research team for efficient utilization of water and fertilizer” (TDZKCX202301).
-
Author contributions: Xinlu Bai and Yaping Li designed the research. Xinlu Bai and Yaping Li analyzed the data. Xinlu Bai wrote the manuscript. Xinlu Bai and Jinhu Zhi acquired fund support. All authors read and approved the final manuscript.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Ackerman D, Millet DB, Chen X. Global estimates of inorganic nitrogen deposition across four decades. Glob Biogeochem Cycles. 2019;33:100–7.10.1029/2018GB005990Suche in Google Scholar
[2] Fowler D, Coyle M, Skiba U, Sutton MA, Cape JN, Reis S, et al. The global nitrogen cycle in the twenty-first century. Philos Trans R Soc B. 2013;368:20130164.10.1098/rstb.2013.0164Suche in Google Scholar PubMed PubMed Central
[3] Reay DS, Dentener F, Smith P, Grace J, Feely RA. Global nitrogen deposition and carbon sinks. Nat Geosci. 2008;1:430–7.10.1038/ngeo230Suche in Google Scholar
[4] Gundersen P, Schmidt IK, Raulund-Rasmussen K. Leaching of nitrate from temperate forests–effects of air pollution and forest management. Environ Rev. 2006;14:1–57.10.1139/a05-015Suche in Google Scholar
[5] Ngaba MJ, Uwiragiye Y, Bol R, de Vries W, Zhou JB. Low-level nitrogen and short-term addition increase soil carbon sequestration in Chinese forest ecosystems. Catena. 2022;215:106333.10.1016/j.catena.2022.106333Suche in Google Scholar
[6] Lebauer DS, Treseder KK. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology. 2008;89:371–9.10.1890/06-2057.1Suche in Google Scholar PubMed
[7] Ning YL, Chen ZD, Gao HD, Yang CB, Zhang X, Wang ZJ, et al. Introducing native tree species alter the soil organic carbon, nitrogen, phosphorus, and fine roots in Moso bamboo plantations. Forests. 2024;15:971.10.3390/f15060971Suche in Google Scholar
[8] Ullah S, Liu WF, Shah JA, Shen F, Liao Y, Duan H, et al. Effects of canopy nitrogen addition and understory vegetation removal on nitrogen transformations in a subtropical forest. Forests. 2024;15:962.10.3390/f15060962Suche in Google Scholar
[9] Liu XY, Koba K, Koyama LA, Liu C. Nitrate is an important nitrogen source for Arctic tundra plants. Proc Natl Acad Sci USA. 2018;115(13):3398–403.10.1073/pnas.1715382115Suche in Google Scholar PubMed PubMed Central
[10] Hu CC, Liu XY, Driscoll AW, Kuang YW, Jack Brookshire EN, Lu XT, et al. Global distribution and drivers of relative contributions among soil nitrogen sources to terrestrial plants. Nat Commun. 2024;15:6407.10.1038/s41467-024-50674-6Suche in Google Scholar PubMed PubMed Central
[11] Xu SQ, Liu XY, Sun ZC, Hu CC, Wanek W, Koba K. Isotopic elucidation of microbial nitrogen transformations in forest soils. Glob Biogeochem Cycles. 2021:35(12):e2021GB007070.10.1029/2021GB007070Suche in Google Scholar
[12] Veerman L, Kalbitz K, Gundersen P, Kjønaas J, Moldan F, Schleppi P, et al. The long-term fate of deposited nitrogen in temperate forest soils. Biogeochemistry. 2020;150:1–15.10.1007/s10533-020-00683-6Suche in Google Scholar
[13] Wang YJ, Cao WC, Zhang XM, Guo JH. Abiotic nitrate loss and nitrogenous trace gas emission from Chinese acidic forest soils. Environ Sci Pollut Res. 2017;24:22679–87.10.1007/s11356-017-9797-4Suche in Google Scholar PubMed
[14] Wessel WW, Boxman AW, Cerli C, van Loon EE, Tietema A. Long-term stabilization of 15N-labeled experimental NH4+ deposition in a temperate forest under high N deposition. Sci Total Environ. 2021;768:144356.10.1016/j.scitotenv.2020.144356Suche in Google Scholar PubMed
[15] Xie DN, Si GY, Zhang T, Mulder J, Duan L. Nitrogen deposition increases N2O emission from an N-saturated subtropical forest in southwest China. Environ Pollut. 2018;243:1818–24.10.1016/j.envpol.2018.09.113Suche in Google Scholar PubMed
[16] Leeson SN, Levy PE, Dijk N, Drewer J, Robinson S, Jones MR, et al. Nitrous oxide emissions from a peatbog after 13 years of experimental nitrogen deposition. Biogeosciences. 2017;14:5753–64.10.5194/bg-14-5753-2017Suche in Google Scholar
[17] Meunier CL, Gundale MJ, Sánchez IS, Liess A. Impact of nitrogen deposition on forest and lake food webs in nitrogen-limited environments. Glob Change Biol. 2016;22:164–79.10.1111/gcb.12967Suche in Google Scholar PubMed
[18] Rennenberg H, Dannenmann M, Gessler A, Kreuzwieser J, Simon J, Papen H. Nitrogen balance in forest soils: nutritional limitation of plants under climate change stresses. Plant Biol. 2009;11:4–23.10.1111/j.1438-8677.2009.00241.xSuche in Google Scholar PubMed
[19] Wang S, Zhuang QL, Jin XX, Yang ZJ, Liu HB. Predicting soil organic carbon and soil nitrogen stocks in topsoil of forest ecosystems in northeastern China using remote sensing data. Remote Sens. 2020;12:1115.10.3390/rs12071115Suche in Google Scholar
[20] Balestrini R, Delconte CA, Buffagni A, Fumagalli A, Freppaz M, Buzzetti I, et al. Dynamic of nitrogen and dissolved organic carbon in an alpine forested catchment: atmospheric deposition and soil solution trends. Nat Conserv. 2019;34:41–66.10.3897/natureconservation.34.30738Suche in Google Scholar
[21] Lu X, Vitousek PM, Mao QG, Gilliam FS, Luo YQ, Mo JM, et al. Nitrogen deposition accelerates soil carbon sequestration in tropical forests. Proc Natl Acad Sci USA. 2021;118:e2020790118.10.1073/pnas.2020790118Suche in Google Scholar PubMed PubMed Central
[22] Zhang J, Zhou J, Sayer EJ, Lambers H, Liu Z, Lu X, et al. Nitrogen deposition enhances soil organic carbon and microbial residual carbon in a tropical forest. Plant Soil. 2023;484:217–35.10.1007/s11104-022-05787-6Suche in Google Scholar
[23] De-Vires FT, Bardgett RD. Plant community control on short-term ecosystem nitrogen retention. N Phytol. 2016;210:861–74.10.1111/nph.13832Suche in Google Scholar PubMed PubMed Central
[24] Li ZL, Zeng ZQ, Tian DH, Wang JS, Fu Z, Zhang FY, et al. Global patterns and controlling factors of soil nitrification rate. Glob Change Biol. 2020;26:4147–57.10.1111/gcb.15119Suche in Google Scholar PubMed
[25] Stevens CJ, Manning P, van den Berg LJL, de Graaf MCC, Wamelink GWW, Boxman AW, et al. Ecosystem responses to reduced and oxidised nitrogen inputs in European terrestrial habitats. Environ Pollut. 2011;159(3):665–76.10.1016/j.envpol.2010.12.008Suche in Google Scholar PubMed
[26] Wang A, Zhu WX, Gundersen P, Phillips OL, Chen DX, Fang YT. Fates of atmospheric deposited nitrogen in an Asian tropical primary forest. For Ecol Manag. 2018;411:213–22.10.1016/j.foreco.2018.01.029Suche in Google Scholar
[27] Liu J, Peng B, Xia ZW, Sun JF, Gao DC, Dai WW, et al. Different fates of deposited NH4+ and NO3− in a temperate forest in northeast China: a 15N tracer study. Glob Change Biol. 2017;23:2441–9.10.1111/gcb.13533Suche in Google Scholar PubMed
[28] Liu XY, Liu MN, Qin WX, Song W. Isotope constraints on nitrate exchanges between precipitation and forest canopy. Glob Biogeochem Cycles. 2023;;37(12):e2023GB007920.10.1029/2023GB007920Suche in Google Scholar
[29] Ding WC, He P, Zhang JJ, Liu YX, Xu XP, Ullah S, et al. Optimizing rates and sources of nutrient input to mitigate nitrogen, phosphorus, and carbon losses from rice paddies. J Clean Prod. 2020;256:120603.10.1016/j.jclepro.2020.120603Suche in Google Scholar
[30] Yu ZP, Chen HYH, Searle EB, Sardans J, Ciais P, Peñuelas J, et al. Whole soil acidification and base cation reduction across subtropical China. Geoderma. 2020;361:114107.10.1016/j.geoderma.2019.114107Suche in Google Scholar
[31] R Core Team. R: a language and environment for statistical computing [Internet]. Vienna, Austria: R Foundation for Statistical Computing; 2018. http://www.r-project.org.Suche in Google Scholar
[32] Gurmesa GA, Wang A, Li SL, Peng SS, de Vries W, Gundersen P, et al. Retention of deposited ammonium and nitrate and its impact on the global forest carbon sink. Nat Commun. 2022;13:1–9.10.1038/s41467-022-28345-1Suche in Google Scholar PubMed PubMed Central
[33] Templer PH, Mack MC, Chapin FS, Christenson LM, Compton JE, Crook HD, et al. Sinks for nitrogen inputs in terrestrial ecosystems: a meta-analysis of 15N tracer field studies. Ecology. 2012;93:1816–29.10.1890/11-1146.1Suche in Google Scholar PubMed
[34] Sun ZC, Ma TY, Xu SQ, Guo HR, Hu CC, Chen CJ, et al. Levels and variations of soil bioavailable nitrogen among forests under high atmospheric nitrogen deposition. Sci Total Environ. 2022;838:156405.10.1016/j.scitotenv.2022.156405Suche in Google Scholar PubMed
[35] Kuzyakov Y, Xu XL. Competition between roots and microorganisms for nitrogen: mechanisms and ecological relevance. N Phytol. 2013;198:656–69.10.1111/nph.12235Suche in Google Scholar PubMed
[36] Fang YT, Koba K, Makabe A, Takahashi C, Zhu WX, Hayashi T, et al. Microbial denitrification dominates nitrate losses from forest ecosystems. Proc Natl Acad Sci USA. 2015;112:1470–4.10.1073/pnas.1416776112Suche in Google Scholar PubMed PubMed Central
[37] Liu XJ, Zhang Y, Han W, Tang A, Shen J, Cui Z, et al. Enhanced nitrogen deposition over China. Nature. 2013;494:459–62.10.1038/nature11917Suche in Google Scholar PubMed
[38] Bai XL, Jiang Y, Miao YZ, Xue SQ, Chen ZJ, Zhou JB. Intensive vegetable production results in high nitrate accumulation in deep soil profiles in China. Environ Pollut. 2021;287:117598.10.1016/j.envpol.2021.117598Suche in Google Scholar PubMed
[39] Weitzman JN, Renée Brooks J, Compton JE, Faulkner BR, Mayer PM, Peachey RE, et al. Deep soil nitrogen storage slows nitrate leaching through the Vadose zone. Agr Ecosyst Environ. 2022;332:107949.10.1016/j.agee.2022.107949Suche in Google Scholar PubMed PubMed Central
[40] Eickenscheidt N, Brumme R, Veldkamp E. Direct contribution of nitrogen deposition to nitrous oxide emissions in a temperate beech and spruce forest - a 15N tracer study. Biogeosciences. 2011;8:621–35.10.5194/bg-8-621-2011Suche in Google Scholar
[41] Cai ZC, Yan XY, Gu BJ. Applying C:N ratio to assess the rationality of estimates of carbon sequestration in terrestrial ecosystems and nitrogen budgets. Carbon Res. 2022;1:2.10.1007/s44246-022-00004-6Suche in Google Scholar
[42] Janssens IA, Dieleman W, Luyssaert J, Subke JA, Reichstein M, Ceulemans R, et al. Reduction of forest soil respiration in response to nitrogen deposition. Nat Geosci. 2010;3:315–22.10.1038/ngeo844Suche in Google Scholar
[43] Vitousek PM, Porder S, Houlton BZ, Chadwick OA. Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen–phosphorus interactions. Ecol Appl. 2010;20(1):5–15.10.1890/08-0127.1Suche in Google Scholar PubMed
[44] Lu X, Mao QG, Gilliam FS, Luo YQ, Mo JM. Nitrogen deposition contributes to soil acidification in tropical ecosystems. Glob Change Biol. 2021;20(12):3790–801.10.1111/gcb.12665Suche in Google Scholar PubMed
[45] Elrys AS, Wang J, Metwally MAS, Chen Y, Zhang JB, Cai ZC, et al. Global gross nitrification rates are dominantly driven by soil carbon-to-nitrogen stoichiometry and total nitrogen. Glob Change Biol. 2021;27:6512–24.10.1111/gcb.15883Suche in Google Scholar PubMed
[46] Elrys AS, Desoky ES, El-Maati MF, Elnahal AS, Abdo AI, Raza S, et al. Can secondary metabolites extracted from Moringa seeds suppress ammonia oxidizers to increase nitrogen use efficiency and reduce nitrate contamination in potato tubers? Ecotox Environ Safe. 2019;185:109689.10.1016/j.ecoenv.2019.109689Suche in Google Scholar PubMed
[47] Hu HW, Zhang LM, Dai Y, Di HJ, He JZ. pH dependent distribution of soil ammonia oxidizers across a large geographical scale as revealed by high-throughput pyrosequencing. J Soil Sediment. 2013;13:1439–49.10.1007/s11368-013-0726-ySuche in Google Scholar
[48] Li YY, Chapman SJ, Nicol GW, Yao HY. Nitrification and nitrifiers in acidic soils. Soil Biol Biochem. 2018;116:290–301.10.1016/j.soilbio.2017.10.023Suche in Google Scholar
[49] Liu ZJ, Ma PY, Zhai BN, Zhou JB. Soil moisture decline and residual nitrate accumulation after converting cropland to apple orchard in a semiarid region: evidence from the Loess Plateau. Catena. 2019;181:104080.10.1016/j.catena.2019.104080Suche in Google Scholar
[50] Zhu XQ, Miao P, Wang PZ, Zhang SF, Chen ZJ, Zhou JB. Variations and influencing factors of nitrate accumulation in the deep soil profiles of apple orchards on the Loess Plateau. Agr Ecosyst Environ. 2022;335:108005.10.1016/j.agee.2022.108005Suche in Google Scholar
[51] Li XX, Sørensen P, Olesen JE, Petersen SO. Evidence for denitrification as main source of N2O emission from residue-amended soil. Soil Biol Biochem. 2016;92:153–60.10.1016/j.soilbio.2015.10.008Suche in Google Scholar
[52] Smith KA. Changing views of nitrous oxide emissions from agricultural soil: key controlling processes and assessment at different spatial scales. Eur J Soil Sci. 2017;68:137–55.10.1111/ejss.12409Suche in Google Scholar
[53] Johnson DW. Nitrogen retention in forest soils. J Environ Qual. 1992;21:1–12.10.2134/jeq1992.00472425002100010001xSuche in Google Scholar
[54] Puri G. Microbial immobilization of 15N-labelled ammonium and nitrate in a temperate woodland soil. Soil Biol Biochem. 1999;31:929–31.10.1016/S0038-0717(98)00172-2Suche in Google Scholar
[55] Huddell AM, Galford GL, Tully KL, Crowley C, Palm CA, Neill C, et al. Meta-analysis on the potential for increasing nitrogen losses from intensifying tropical agriculture. Glob Change Biol. 2019;26:1668–80.10.1111/gcb.14951Suche in Google Scholar PubMed
[56] Northup RR, Yu Z, Dahlgren RA, Vogt KA. Polyphenol control of nitrogen release from pine litter. Nature. 1995;377(6546):227–9.10.1038/377227a0Suche in Google Scholar
[57] Elrys AS, Chen ZX, Wang J, Uwiragiye Y, Helmy AM, Desoky ESM, et al. Global patterns of soil gross immobilization of ammonium and nitrate in terrestrial ecosystems. Glob Change Biol. 2022;28:4472–88.10.1111/gcb.16202Suche in Google Scholar PubMed
[58] Widdig M, Heintz-Buschart A, Schleuss PM, Guhr A, Borer ET, Seabloom EW, et al. Effects of nitrogen and phosphorus addition on microbial community composition and element cycling in a grassland soil. Soil Biol Biochem. 2020;151:108041.10.1016/j.soilbio.2020.108041Suche in Google Scholar
[59] Gay JD, Currey B, Brookshire ENJ. Global distribution and climate sensitivity of the tropical montane forest nitrogen cycle. Nat Commun. 2022;13:1–8.10.1038/s41467-022-35170-zSuche in Google Scholar PubMed PubMed Central
[60] Ballard TC, Sinha E, Michalak AM. Long-term changes in precipitation and temperature have already impacted nitrogen loading. Environ Sci Technol. 2019;53:5080–90.10.1021/acs.est.8b06898Suche in Google Scholar PubMed
[61] Schaefer SC, Alber M. Temperature controls a latitudinal gradient in the proportion of watershed nitrogen exported to coastal ecosystems. Biogeochemistry. 2007;85:333–46.10.1007/s10533-007-9144-9Suche in Google Scholar
[62] Zhang BW, Li WJ, Chen SP, Tan XR, Wang SS, Chen ML, et al. Changing precipitation exerts greater influence on soil heterotrophic than influence on soil heterotrophic than autotrophic respiration in a semiarid steppe. Agr For Meteorol. 2019;271:413–21.10.1016/j.agrformet.2019.03.019Suche in Google Scholar
[63] Tang WG, Chen DX, Phillips OL, Liu X, Zhou Z, Li YD, et al. Effects of long-term increased N deposition on tropical montane forest soil N2 and N2O emissions. Soil Biol Biochem. 2018;126:194–203.10.1016/j.soilbio.2018.08.027Suche in Google Scholar
[64] Tian DS, Niu SL. A global analysis of soil acidification caused by nitrogen addition. Environ Res Lett. 2015;10:024019.10.1088/1748-9326/10/2/024019Suche in Google Scholar
[65] Liu WJ, Yu LF, Zhang T, Kang RH, Zhu J, Mulder J, et al. In situ 15N labelling experiment reveals different long-term responses to ammonium and nitrate inputs in N-saturated subtropical forest. Biogeosciences. 2017;122:2251–64.10.1002/2017JG003963Suche in Google Scholar
© 2025 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- 10.1515/biol-2025-1168
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”
Artikel in diesem Heft
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- 10.1515/biol-2025-1168
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”